响应面法优化高产ACE 抑制肽发酵乳生产工艺及其质构和风味特性研究
2023-10-19王家栩贾丽丽李嘉欣张密霞王志敏马春丽
王家栩,贾丽丽,李嘉欣,张密霞,王志敏,马春丽
(东北农业大学食品学院,黑龙江哈尔滨 150030)
血管紧张素转换酶(Angiotensin-I converting enzyme,ACE)是一种具有双结构域的二肽基羧基酶,在机体血压调节的过程中起着至关重要的生理作用[1]。它可以水解血管紧张素I,使之变成具有收缩血管功能的血管紧张素II,同时水解具有舒张血管作用的缓激肽和N-乙酰基-SDKP,造成机体血压的升高[2]。因此,抑制ACE 的生物活性对防治高血压具有十分重要的意义[3]。
来源于食源蛋白质的ACE 抑制肽因原料丰富、安全性高的特点具有显著优势[4],不仅为开发新型ACE 抑制剂药物提供了新思路,且可以作为功能性成分广泛应用于保健食品中,起到预防心脑血管疾病的作用。目前,已有报道从乳蛋白如牛乳酪蛋白[5]、羊乳酪蛋白[6]、乳清蛋白[7];植物蛋白如玉米蛋白[8]、藜麦分离蛋白[9]、花生蛋白[10];动物源蛋白如肌原纤维蛋白[11]、血红蛋白[12]等中获取了ACE 抑制肽,且取得良好效果。乳酸菌发酵是生产ACE 抑制肽的一种常见方法,在发酵过程中,微生物自身的蛋白酶系统能够水解乳蛋白,产生具有ACE 抑制活性的肽,赋予发酵乳ACE 抑制活性[13]。日本Calpis 公司和芬兰Evolu 公司推出了含有ACE 抑制肽(VPP和IPP)的发酵乳制品,并广受消费者好评[14-15]。因此,开发具有降血压活性的发酵乳对于食品工业有着广泛的应用价值和前景。
蛋白质强化被证实可以提高发酵乳的ACE 抑制活性并改善其品质[16]。一些研究结果显示,经过蛋白质强化后的发酵乳组织更加紧密,乳清析出更少,同时弹性模量和黏性膜量显著提高,形成了更强的凝胶网络[17]。此外,发酵乳的风味主要由乳酸和挥发性物质在发酵过程中生成,因此蛋白质的改变还会对发酵乳滋味产生影响[18]。Bierzuńska 等[19]研究发现,添加聚合乳清蛋白的酸奶的保水性、内聚性和黏性指数均明显提高,香味更加浓郁。El-Fattah 等[20]使用乳清蛋白对牛奶进行强化,制备了具有心脑血管保健功能的发酵乳。Iwaniak 等[21]发现经过β-酪蛋白强化后,高达干酪的肽谱发生变化,ACE 抑制活性显著提高。酪蛋白酸钠作为一种蛋白质强化剂,具有良好的增稠性、乳化性和起泡性[22-23],常被用于发酵乳中以起到增加黏度、提高硬度、减少乳清析出的作用[24]。在本研究中,使用酪蛋白酸钠作为蛋白质强化剂,以提高发酵乳的ACE 抑制活性、改善发酵乳的品质。除此之外,本实验室在前期研究中发现,分离来自生牛乳的Lb.plantarumM11 具有良好的耐酸、耐盐、耐胆盐以及蛋白水解能力,作为附属发酵剂添加至发酵乳中也可以显著提高发酵乳的ACE抑制活性。为此,本研究以获得高产ACE 抑制肽发酵乳为目标,使用响应面法对高产ACE 抑制肽发酵乳的生产工艺进行优化,并研究了Lb.plantarumM11 和酪蛋白酸钠的添加对发酵乳质构和风味特性的影响,为生产富含ACE 抑制肽的功能性发酵乳提供理论依据。
1 材料与方法
1.1 材料与仪器
脱脂乳粉、酪蛋白酸钠(CS)新西兰恒天然有限公司;发酵剂YO-MIX 883(含有嗜热链球菌和保加利亚乳杆菌)丹麦Danisco 公司;Lb.plantarumM11 分离来自新鲜无抗牛乳并保藏于东北农业大学实验室;马尿酰-组氨酰-亮氨酸(N-Hippuryl-His-Leu hydrate,HHL)、ACE(来自兔肺,酶活0.1 UN) 源叶生物科技(上海)有限公司;其他试剂 均为国产分析纯。
UV-1500C 紫外-可见分光光度计 上海美析仪器有限公司;GL-21M高速冷冻离心机 湖南湘仪实验仪器开发有限公司;TA.XT plus 质构仪 英国SMS 公司;SA-402B 型电子舌 日本INSENT 公司。
1.2 实验方法
1.2.1 发酵乳的制作工艺 12%的脱脂乳中加入8%白砂糖,充分混匀后分装,于95 ℃条件下灭菌15 min 后冷却至43 ℃。在超净工作台内接入发酵剂发酵至凝乳,然后在4 ℃条件下后熟12 h 备用[25]。
1.2.2 发酵乳ACE 抑制活性的测定 取5 mL 发酵乳样品,用0.1 mol/L NaOH 和0.1 mol/L HCl 溶液将样品pH 调至3.5,在4 ℃条件下以8000 r/min 离心15 min,取上清液调节pH 至8.3,再离心20 min,收集上清液备用。参考Roy 等[26]所述的方法测定发酵乳的ACE 抑制活性。按表1 顺序依次加入HCl 溶液(1 mol/L)、HHL(5 mmol/L)、样品、ACE(0.1 U/mL)、HCl 溶液(1 mol/L)以及硼酸缓冲液。然后向每组中加入1.7 mL 乙酸乙酯,振荡摇匀,于3000 r/min 离心10 min。离心结束后吸取上层乙酸乙酯置于120 ℃恒温箱中烘干,待温度降低到室温之后加入3 倍体积的蒸馏水,混匀之后在228 nm下测定溶液的吸光值。

表1 ACE 抑制率的测定方法Table 1 Detection methods of ACE inhibition
ACE 抑制率计算公式如下:
式中:A、B、C 分别为A 组、B 组和C 组在228 nm处的OD 值。
1.2.3 单因素实验 参考1.2.1 中所述方法制备发酵乳。在接种量1×107CFU/mL、发酵温度42 ℃的条件下,考察Lb.plantarumM11 和发酵剂YO-MIX 883 加入比例为0:1、1:3、1:2、1:1、2:1、3:1 和1:0时发酵乳的ACE 抑制活性;在Lb.plantarumM11和发酵剂YO-MIX 883 接种比例为2:1、发酵温度42 ℃的条件下,考察菌株总接种量为0.5×107、1×107、1.5×107、2×107、2.5×107CFU/mL 时发酵乳的ACE 抑制活性;在Lb.plantarumM11 和发酵剂YO-MIX 883 接种比例为2:1、菌株总接种量为1×107CFU/mL 的条件下,考察发酵温度为35、37、39、42、45 ℃时发酵乳的ACE 抑制活性;在Lb.plantarumM11 和发酵剂YO-MIX 883 接种比例为2:1、菌株总接种量为1×107CFU/mL、发酵温度42 ℃的条件下,考察添加0%、0.5%、1%、1.5%、2%、2.5%酪蛋白酸钠对发酵乳ACE 抑制活性的影响。
1.2.4 响应面试验 根据Box-Behnken 试验设计原理,在上述单因素实验的基础上选择酪蛋白酸钠添加量、Lb.plantarumM11 和发酵剂YO-MIX883 的接种量以及发酵温度作为考察因素进行三因素三水平的响应面试验。以ACE 抑制率(%)作为响应值,使用Design Expert V8.0.6 建立模型,以获得生产高产ACE 抑制肽发酵乳的最佳工艺条件。具体试验因素及水平设计见表2。

表2 响应面试验因素及水平设计Table 2 Factors and levels for response surface test
1.2.5Lb.plantarumM11 和酪蛋白酸钠对发酵乳质构和风味特性的影响
1.2.5.1 实验分组和处理 制备四组不同的发酵乳,以进一步探究Lb.plantarumM11 和酪蛋白酸钠的强化对发酵乳质构和风味特性的影响。各组发酵乳发酵剂YO-MIX 883、Lb.plantarumM11 的接种量以及酪蛋白酸钠添加量见表3。

表3 各组发酵乳发酵剂YO-MIX 883、Lb.plantarum M11 的接种量以及酪蛋白酸钠添加量Table 3 inoculation quantity of YO-MIX 883,Lactobacillus plantarum M11 and additional quantity of sodium caseinate in each group
1.2.5.2 发酵乳质构特性的测定 使用TA-XT plus质构仪对发酵乳贮藏期的质构特性进行测定。将装有50 mL 发酵乳的发酵杯置于检测台中心,用A/BE-d35(盘径35 mm)探头测定发酵乳的硬度、稠度、粘聚性和粘性指数[27]。
1.2.5.3 电子舌对滋味的测定 使用电子舌测定发酵乳的滋味。电子舌配有5 根脂质膜传感器(AAE、CA0、CT0、C00、AE1)分别负责鲜味、咸味、酸味、苦味和涩味的测定。准确称量20 mL发酵乳样品,加入三倍体积的去离子水进行稀释,然后置于电子舌样品上的品尝杯中进行测定[28]。
1.3 数据处理
所有实验重复三次,数据以平均值±标准差表示。使用IBM SPSS 25 对实验数据进行统计分析,并用GraphPad Prism 9 和Origin 2018 进行绘图;使用Design Expert V8.0.6 分析响应面试验结果。
2 结果与分析
2.1 单因素实验
2.1.1Lb.plantarumM11 和发酵剂YO-MIX 883 接种比例对发酵乳ACE 抑制活性的影响 如图1 所示,随着Lb.plantarumM11 与发酵剂YO-MIX 883的接种比例由0:1 上升至2:1,发酵乳的ACE 抑制活性呈现上升趋势。但当Lb.plantarumM11 与发酵剂YO-MIX 883 的接种比例继续增大时,发酵乳的ACE 抑制活性降低,这可能是由于发酵剂含量过低,Lb.plantarumM11、嗜热链球菌和保加利亚乳杆菌之间无法形成良好的协同作用,蛋白水解度降低,ACE 抑制肽含量减少。相比单独使用发酵剂YOMIX 883 进行发酵(即Lb.plantarumM11 与发酵剂YO-MIX 883 接种比例为0:1 时),添加附属发酵剂Lb.plantarumM11 的组别ACE 抑制活性都提高了,这是因为附属发酵剂菌株的添加能够改变发酵乳的肽谱,促进ACE 抑制肽释放[29-30]。Moslehishad 等[31]研究证明,使用鼠李糖乳杆菌作为附属发酵剂可以促进发酵乳中生物活性肽的释放,提高发酵乳的ACE抑制活性,这与本研究所获结果相似。综上,后续实验中Lb.plantarumM11 和YO-MIX 883 接种比例确定为2:1。

图1 Lb.plantarum M11 与发酵剂YO-MIX 883 的接种比例对发酵乳ACE 抑制活性的影响Fig.1 Effect of inoculation ratio of Lb.plantarum M11 and YO-MIX 883 on ACE inhibitory activity of fermented milk
2.1.2 菌株接种量对发酵乳ACE 抑制活性的影响 如图2 所示,随着接种量由0.5×107CFU/mL 升高到1.5×107CFU/mL,发酵乳的ACE 抑制活性逐渐增强(P<0.05)。当接种量达到1.5×107CFU/mL 时,发酵乳具有最强的ACE 抑制活性,但当接种量进一步提高时,发酵乳的ACE 抑制活性反而下降。这可能是由于随着接种量的上升,菌株在相同时间内生长速度更快,产肽更多。但当接种量过大时,菌体产酸过快,达到发酵终点所需时间过短,从而抑制了ACE 抑制肽的产生[32]。综上,后续实验中菌株接种量取值范围为1×107~2×107CFU/mL。
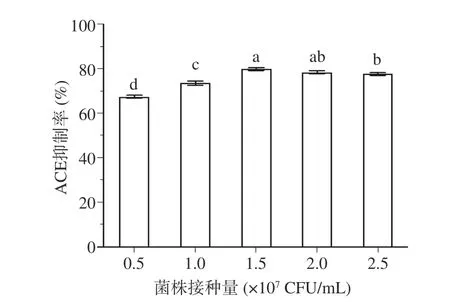
图2 菌株接种量对发酵乳ACE 抑制活性的影响Fig.2 Effect of inoculation quantity on ACE inhibitory activity of fermented milk
2.1.3 发酵温度对ACE 抑制活性的影响 如图3所示,发酵温度的改变对发酵乳的ACE 抑制率有着明显影响,即每组之间都呈现显著差异(P<0.05)。当发酵温度从35 ℃升高到39 ℃时,发酵乳的ACE抑制活性逐渐升高,在39 ℃达到最大值。但随着温度进一步升高,发酵乳的ACE 抑制活性呈现下降趋势。这是因为随着温度的升高,菌株生长代谢速度变快,使蛋白水解度上升,产肽量升高;而当温度超过菌株最适生长温度时,菌株的生长受到高温环境影响,产肽降低[32]。综上,较高或较低的温度都会降低发酵乳的ACE 抑制活性,后续实验中发酵温度的取值范围应为37~42 ℃。

图3 发酵温度对发酵乳ACE 抑制活性的影响Fig.3 Effect of fermentation temperature on ACE inhibitory activity of fermented milk
2.1.4 酪蛋白酸钠对发酵乳ACE 抑制活性的影响 如图4 所示,当酪蛋白酸钠的含量从0%上升至2%时,发酵乳的ACE 抑制活性呈现上升趋势。相比于不添加酪蛋白酸钠的组别,酪蛋白酸钠添加量为2%的发酵乳的ACE 抑制活性显著增强(P<0.05)。当酪蛋白酸钠添加量继续增加至2.5%时,其ACE抑制活性开始下降,但下降趋势不显著(P>0.05)。这一结果与Giacomettei 等[33]的研究结果一致,即发酵乳的ACE 抑制活性会随着蛋白浓度的上升而增加,但当蛋白质浓度达到一定阈值后,ACE 抑制活性不会继续上升,MALDI-TOF-MS 分析证实蛋白含量的增加在一定程度上可以提高肽的含量,从而起到提高ACE 抑制活性的作用。综上,为了获得ACE 抑制活性最强的发酵乳,酪蛋白酸钠添加量应该控制在1.5%~2.5%。
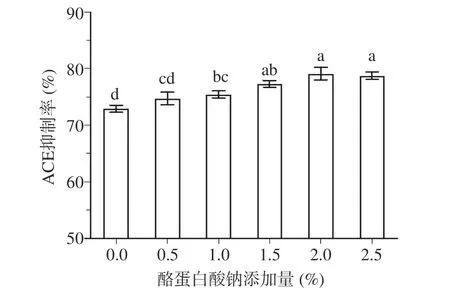
图4 酪蛋白酸钠对发酵乳ACE 抑制活性的影响Fig.4 Effect of sodium caseinate on ACE inhibitory activity of fermented milk
2.2 响应面试验优化结果
2.2.1 响应面优化试验设计及结果 在单因素实验的基础上,选择ACE 抑制率作为响应值,酪蛋白酸钠添加量(A)、接种量(B)以及发酵温度(C)作为变量,进行响应面试验,响应面试验结果如表4 所示。
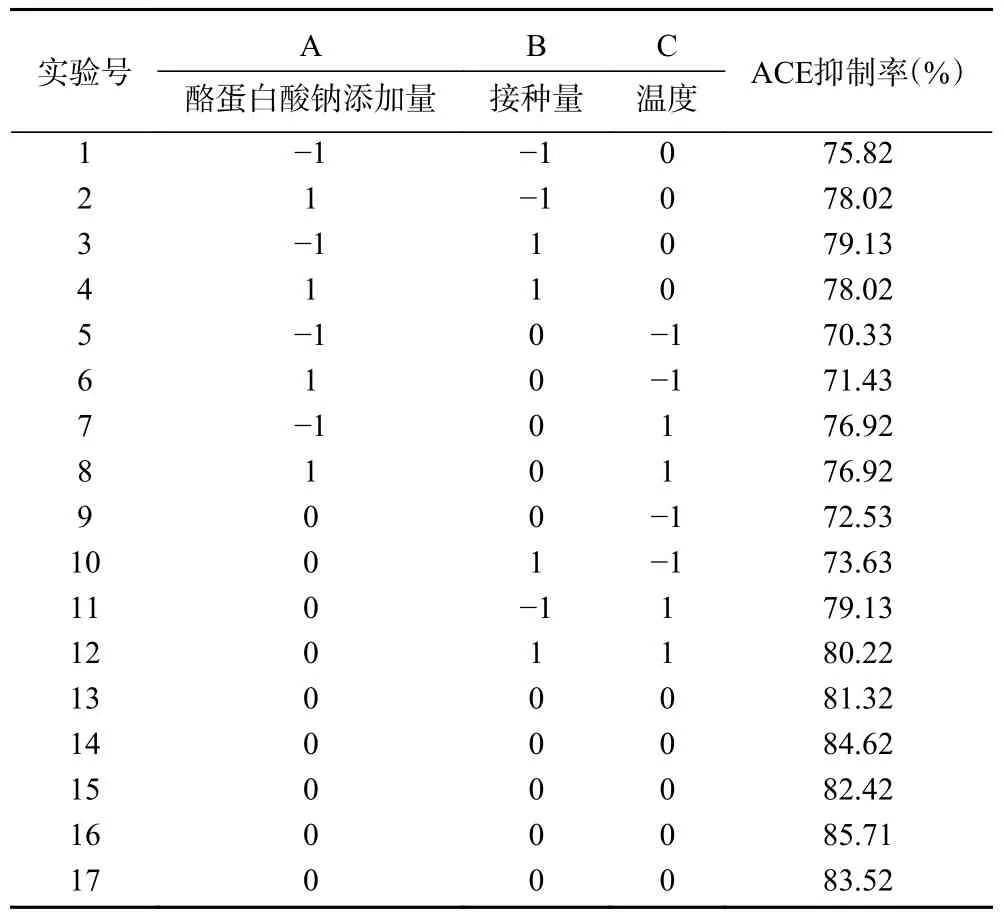
表4 响应面试验设计及结果Table 4 Design and results of response surface test
2.2.2 回归模型的建立及方差分析 对表4 结果进行多元回归拟合,建立高产ACE 抑制肽发酵乳发酵工艺的回归模型。最终获得的二次多项回归模型方程为:Y=84.40+0.25A+0.68B+3.16C-0.83AB-0.27AC-0.030BC-4.12A2-1.65B2-6.38C2。
二次多项回归模型的各因素方差分析结果见表5。回归模型P值为0.0003,表现为极显著,失拟项P值为0.9901,表现为不显著,说明其他因素对该试验结果影响很小,回归方程拟合效果较好,该试验设计方案合理可靠,模型具有统计学意义。决定系数R2为0.9624,校正决定系数R2Adj为0.9141,两者数值相近,说明该模型可以解释绝大部份的响应值变化,可以用于优化高产ACE 抑制肽发酵乳的生产工艺。变异系数CV 为1.70%,远小于10%,表明该试验结果的可信度和精确度较好。二次多项回归模型中的二次项C、A2、C2对模型影响极显著,B2对模型影响显著,各因素对发酵乳ACE 抑制率的影响从大到小依次为C(发酵温度)>B(接种量)>A(酪蛋白酸钠添加量)。

表5 二次多项回归模型的各因素方差分析结果Table 5 Analysis of various factors of the second multiple regression model
2.2.3 响应面交互分析 各因素交互作用的响应面3D 图见图5。三因素之间的交互作用响应面3D 图都表现为开口向下的曲面,曲面最高点即为试验最终所求的最优工艺条件。图5a 为接种量和酪蛋白酸钠添加量之间的交互作用结果,当发酵温度一定时,随着接种量和酪蛋白酸钠添加量的增加,发酵乳ACE 抑制率先升高后降低,响应面曲面连续,存在峰值。图5b 为发酵温度和酪蛋白酸钠添加量之间的交互作用结果,两个因素的交互作用存在明显峰值,发酵乳的ACE 抑制率在曲面最高点达到最大值。图5c 为发酵温度和接种量之间的交互作用结果,发酵温度对ACE 抑制率的影响明显强于接种量,与表3中显著性检验结果相吻合,发酵乳的ACE 抑制率随着发酵温度的升高先上升后下降,在所选范围内存在峰值。

图5 酪蛋白酸钠添加量、菌株接种量以及发酵温度对发酵乳ACE 抑制率影响的响应面3D 图Fig.5 Response surface 3D plot of the effects of sodium caseinate addition,inoculation quantity and fermentation temperature on ACE inhibition rate of fermented milk
2.2.4 发酵乳ACE 抑制率最优值的确定及验证试验响应面交互分析经过预测,当酪蛋白酸钠添加量为2%,接种量为1.60×107CFU/mL,发酵温度为40.12 ℃时,发酵乳具有最高的ACE 抑制率,在此条件下模型预测发酵乳ACE 抑制率可以达到84.87%。考虑到试验操作的可行性,修正发酵工艺为酪蛋白酸钠添加量2%、接种量1.5×107CFU/mL,发酵温度40 ℃。在此条件下进行3 次平行试验验证优化结果,其ACE 抑制率稳定在83.15%,与预测值较为接近,说明该模型可以准确地预测酪蛋白酸钠添加量、菌株接种量以及发酵温度对发酵乳ACE 抑制率的影响效果。
2.3 Lb.plantarum M11 和酪蛋白酸钠对发酵乳质构性质和风味特性的影响
2.3.1 发酵乳的质构特性 发酵乳的质构特性是评价发酵乳品质的重要因素,主要包括硬度、稠度、粘聚性以及粘性指数四个方面。四组发酵乳在贮藏期间的质构特性测试结果见表6。

表6 发酵乳在贮藏期的质构特性Table 6 Texture characteristics of fermented milk during storage
硬度代表探头在下压过程中达到最大深度所需要的力。四组发酵乳中,883+M11 组具有最低的硬度值。添加酪蛋白酸钠后发酵乳的硬度显著增大(P<0.05),且883+M11-CS 组的硬度比883-CS 组更大(P<0.05)。这可能是因为蛋白质浓度越高,发酵乳形成的凝胶强度越大,硬度提高。在贮藏期间,四组发酵乳的硬度都呈现上升趋势,在第28 d 达到最大值。
稠度代表力与时间之间形成的峰面积,反映了发酵乳的流动能力[34]。883+M11-CS 组的稠度显著高于其他三组(P<0.05),说明该组发酵乳内部的微观结构排列更为紧密,稳定性更高[35]。883+M11 组和883-CS 组的稠度都显著高于883 组(P<0.05),说明酪蛋白酸钠和Lb.plantarumM11 都可以提高发酵乳的稠度。王鑫磊等[18]也发现了类似的结果,这可能是附属发酵剂的添加生成了更多的胞外多糖,这些胞外多糖与蛋白质基质之间发生静电相互作用,使发酵乳胶着性增加,稠度上升。在贮藏期间,883 组和883+M11 组的稠度都呈现下降趋势,而添加酪蛋白酸钠的883-CS 组和883+M11-CS 组稠度变化趋势不明显。
粘聚性代表样品的内部粘合力,为负向最大力。883+M11-CS 组的粘聚性显著高于其他三组(P<0.05),883+M11 组与883 组之间的粘聚性差异不显著(P>0.05),但883-CS 组与883 组之间呈现显著性差异(P<0.05),这说明酪蛋白酸钠对发酵乳的粘聚性存在明显的影响,这可能是在酸化过程中,酪蛋白颗粒附着在簇和链结构上,从而形成由酪蛋白簇和链组成的三维网状结构[36],使发酵乳内部环境更完整紧密。发酵乳在贮藏期间的粘聚性指标总体呈现上升趋势,但这种趋势在贮藏期第1~7 d 内表现不显著(P>0.05)。
粘性指数代表力与时间之间形成的负峰面积,即发酵乳对返还探头的粘着力。883+M11-CS 组的粘性指数要显著高于其他组(P<0.05),883+M11 组和883-CS 组的粘性指数都显著高于883 组(P<0.05)。这说明酪蛋白酸钠和Lb.plantarumM11 的添加都提高了发酵乳的凝胶特性,且二者具有协同作用。在贮藏期间,883 组和883-CS 组的粘度指数呈现上升趋势,而883+M11 组和883+M11-CS 组的变化趋势不明显。
2.3.2 发酵乳滋味的测定 电子舌是一种通过模拟人体味觉系统对样品滋味进行测定的设备。与感官评价相比,电子舌获得的信息更加客观,且具有重复性[37]。由滋味雷达图6 可知,四组发酵乳之间的酸味差异最为明显,附属发酵剂Lb.plantarumM11 会增加发酵乳的酸味,而添加酪蛋白酸钠的883-CS 组和883+M11-CS 组酸味弱于883 组。此外Lb.plantarumM11 还可以增加发酵乳滋味的丰富性,这一结果与Lesme 等[38]研究结果一致,即附属发酵剂的使用可以提高发酵乳滋味的丰富性。四组发酵乳之间的咸味、苦味、涩味、涩味回味、苦味回味以及鲜味差异均不明显。滋味的主成分分析如图7 所示,PC1 贡献率为88.0%,PC2 贡献率为9.4%,总贡献率为97.4%,说明该分析可以解释绝大部分的发酵乳滋味信息,PC1 远远大于PC2,说明样品在X 轴方向距离越大,差异性越大。883 组和883+M11 组位于X 轴正半轴,与丰富性、鲜味、酸味和苦味呈正相关;而883-CS 组和883+M11-CS 组位于X 轴负半轴,与咸味、涩味、涩味回味和苦味回味呈正相关。此外,883+M11-CS 组与883 组在X 轴方向上距离较近,说明它们整体滋味相似。

图6 发酵乳的滋味雷达图Fig.6 Taste radar of fermented milk

图7 滋味的主成分分析Fig.7 Principal component analysis of taste
3 结论
发酵乳制品是生物活性肽的重要来源,生产研发具有心脑血管保健功能的发酵乳制品具有广泛的应用价值和前景[39]。本研究通过对原料乳进行酪蛋白强化,并使用Lb.plantarumM11 作为附属发酵剂,提高了发酵乳的ACE 抑制活性。结果表明,酪蛋白酸钠的使用和附属发酵剂Lb.plantarumM11的添加都可以显著提高发酵乳的ACE 抑制活性,当酪蛋白酸钠添加量为2%、接种量为1.5×107CFU/mL,发酵温度为40 ℃时,发酵乳具有最强的ACE 抑制活性,在此条件下其ACE 抑制率为83.15%。此外,酪蛋白酸钠和Lb.plantarumM11 的添加可以改善发酵乳的质构特性,增强发酵乳的硬度、稠度、粘聚性和粘度指数,能够在对风味无不良影响的基础上,满足消费者对饮食健康的追求。但生物活性肽只有经过肠道吸收进入血液循环后才能发挥其生物活性,胃肠道消化可能会对肽段产生巨大影响,导致生物活性发生改变。因此未来还需要进一步对发酵乳中的肽谱进行分析,验证其在体内的作用效果和降血压机制,发掘其健康功效和应用价值。
