紫花苜蓿胚性愈伤组织诱导及胚状体分化研究
2023-10-13王晓春高婷杨炜迪王川陈彩锦
王晓春 高婷 杨炜迪 王川 陈彩锦
(1 宁夏农林科学院动物科学研究所,750000,宁夏银川;2 宁夏农林科学院固原分院,756000,宁夏固原)
紫花苜蓿(MedicagosativaL.)被誉为“牧草之王”,具有适应性强、产草量高和营养丰富等优点,是世界各国人工草地和畜牧养殖中重要的草种[1]。近年来,随着我国“振兴奶业苜蓿发展行动”和“粮改饲”等政策的实施,苜蓿的种植面积逐渐扩大,已是我国种植面积最大的豆科牧草[2]。市场上流通的苜蓿种子主要是国外育成品种。紫花苜蓿传统育种方法周期长,制约了育种速度,国产自主苜蓿品种的研发任重道远。转基因分子育种技术通过外源目的基因的高效转化,可加速育种进程。
体细胞胚胎再生具有嵌合体少、多再生型和遗传背景一致的优点[3],是一种常用的高效遗传转化途径。有关紫花苜蓿体胚途径建立再生体系的研究相对较多,李玉珠等[4]以9 个苜蓿品种的下胚轴为外植体,通过不同激素配比,有2 个品种最终诱导出体细胞胚。温之雨等[5]通过愈伤诱导和体胚诱导等几个关键环节的研究,最终建立了紫花苜蓿品种三得利的高频再生体系。但紫花苜蓿体细胞胚胎再生途径依然存在培养周期长、分化率低和可重复性差等缺点。本研究以紫花苜蓿地方品种固原紫花为对象,开展不同外植体、激素种类和激素浓度配比的研究,旨在建立该品种高频再生体系,为该品种的遗传转化提供稳定的受体。
1 材料与方法
1.1 材料及无菌苗培养
供试材料为紫花苜蓿地方品种固原紫花。将固原紫花干种子在水龙头下冲洗,留下饱满的种子,在超净工作台消毒,先用75%的酒精振荡消毒1min,无菌水冲洗3 遍,再用0.1%升汞消毒15min,冲洗后接种在1/2 MS 培养基中培养。
1.2 外植体选取
取发芽7d 后无菌苗的下胚轴、茎段、子叶和健壮再生苗的幼嫩叶片和叶柄,并将下胚轴、茎段和叶柄切割成1cm 长小段,子叶和真叶划割四边,接种于诱导培养基内。
1.3 不同2,4-D 浓度对苜蓿愈伤/胚性愈伤组织的诱导
以MS 培养基为基本培养基,分别添加0、2、4、6、8mg/L 的2,4-二氯苯氧乙酸(2,4-D)处理,接种下胚轴、茎段和子叶3 种外植体,每个处理20 瓶,每瓶15 个外植体。待完全脱分化后,转入1/2 MS 培养基内分化,统计出愈率,记录完全脱分化时间、愈伤组织状态及体细胞胚发育情况。12d 继代一次,继代培养基为原培养基。
1.4 2,4-D 与不同细胞分裂素组合对苜蓿愈伤/胚性愈伤组织的诱导
以MS 培养基为基本培养基,通过低浓度(0.5mg/L)的细胞分裂素6-苄基氨基嘌呤(6-BA)、激动素(KT)与初筛出的2,4-D 浓度范围进行组合,配方见表1。待完全脱分化后观测出愈率,并记录愈伤组织状态及体细胞胚发育情况。
1.5 胚性愈伤组织/体胚苗的分化培养
将脱分化后的愈伤组织转入分化培养基。分化培养基以1/2 MS 培养基为基本培养基,添加低浓度6-BA、KT 和NAA,设置6 个分化培养基处理:D0(不添加对照,CK)[6]、D1(0.5mg/L KT+0.1mg/L NAA)、D2(0.5mg/L KT+0.3mg/L NAA)[7]、D3(1.0mg/L KT+0.5mg/L NAA)[8]、D4(0.4mg/L KT)[9]、D5(2.0mg/L 6-BA+0.5mg/L NAA)[10],D2 根据D3 降低了NAA 浓度。
1.6 再生苗的诱导及体细胞的分化
分化出的再生苗经培养基1/2 MS+0.1mg/L 吲哚丁酸(IBA)进行生根、壮苗培养后,以叶片和叶柄为外植体,再经诱导和分化培养,观察再生苗的诱导、分化情况。
1.7 培养条件
所有培养基pH 均调至5.8,蔗糖浓度30g/L,琼脂粉7.5g/L。在智能人工气候箱内培养,温度25℃±2℃,除愈伤阶段暗处理,其他培养均为16h/8h 光暗周期,光照强度2000lx,每天观察并及时挑拣出生菌的培养瓶。
1.8 数据处理
愈伤组织诱导率(%)=完全脱分化的愈伤组织数/接种的外植体数×100;分化率(%)=有体细胞胚发生的愈伤组织数/接种的愈伤组织数×100;成苗率(%)=发育出茎、叶的植株数/接种的有体胚发生的愈伤组织数×100。
利用Excel 对数据进行整理,用SPSS 17.0 软件对结果进行Duncan 法多重比较。
2 结果与分析
2.1 不同浓度2,4-D 对苜蓿愈伤/胚性组织诱导的影响
由表2 可知,不添加2,4-D 的对照处理外植体诱导率为0.0%,与添加2,4-D 处理间差异显著(P<0.05)。当2,4-D 浓度为2、4mg/L 时,诱导率都达到100.0%,约24d 后完全脱分化,愈伤块在颜色和质地上无明显区别,但整体上前者比后者体积小。当2,4-D 浓度为6mg/L 时,诱导率下降为84.7%,愈伤块体积较2,4-D 4mg/L 处理小;当增大到8mg/L 时,诱导率降到76.6%,愈伤块体积最小,且在继代培养过程中,脱分化愈伤组织开始褐化时间早于其他处理。由此可见,高浓度的2,4-D 对外植体愈伤化有抑制作用。

表2 不同浓度2,4-D 对苜蓿愈伤组织的诱导Table 2 Induction effects of different concentrations of 2,4-D on alfalfa callus
2.2 不同外植体的愈伤效果
将无菌苗的茎段、下胚轴和叶片3 种外植体分别放置在诱导培养基上诱导,诱导率均达到100.0%。下胚轴和幼茎放置3d 后愈伤化启动,增粗,逐渐蓬松。大概4d 后可见叶片增厚(图1)。下胚轴和幼茎愈伤速度前期稍快于叶片,但经继代培养,24d 后都能完全脱分化,不同来源外植体愈伤组织在形态上无明显区别。

图1 固原紫花的不同外植体及对应的出愈状态Fig.1 Different explants of Guyuan alfalfa and corresponding callus status
2.3 2,4-D 与不同细胞分裂素组合对苜蓿胚性愈伤组织诱导及分化的影响
常见的苜蓿胚性愈伤组织诱导是通过生长素2,4-D 与低浓度细胞分裂素6-BA、KT 的组合。通过2.2 结果可见,2,4-D 浓度在6mg/L 及以上时对苜蓿愈伤组织的诱导具有抑制作用。采用表1 配方,经过20~25d 的诱导,除了CK,其余10 个组合外植体都已完全脱分化,诱导率达到100%(表3)。当2,4-D 浓度为5mg/L 时,诱导出的愈伤块体积略大于其他浓度,但愈伤块在颜色和质地上无区别,说明1~5mg/L 2,4-D 组合0.5mg/L 的6-BA与KT 对愈伤组织的形成有促进作用。添加6-BA和KT 后形成的愈伤组织在质地上有区别,前者为黄绿色,后者整体偏白色(图2)。

图2 不同细胞分裂素(6-BA、KT)与2,4-D 组合形成的愈伤组织Fig.2 Callus formed by combinations of different cytokinins(6-BA,KT)with 2,4-D
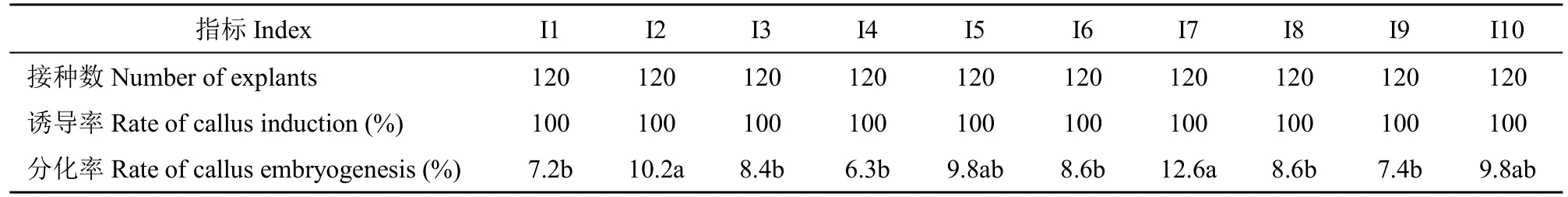
表3 不同诱导培养基的愈伤块的分化Table 3 Differentiation of callus in different induction media
分别将以上完全脱分化的愈伤块转入不添加任何激素且培养条件一致的分化培养基1/2 MS 中(参照李娟等[9]的文章),观察分化出的胚状体,统计分化率(表3)。
经过20~30d 的培养,主要形成2 种类型的愈伤组织,一种是有绿色球形胚或有绿色芽点的愈伤组织,多次继代,不失活,属于胚性愈伤组织;另一种是无组织状态、继代1 次后逐渐褐化、失活,为非胚性愈伤组织(图3)。I1~I10 培养基诱导出的愈伤组织在1/2 MS 培养基内都分化出了胚状体,但分化率都不高。添加6-BA 的组合(I1~I5)中,I2 组合的分化率最高(10.2%),I1 组合最低(7.2%)。添加KT 的组合(I6~I10)中,I7 组合的分化率最高(12.6%),I9 组合最低(7.4%)。由此可见,分化率最高的培养基2,4-D 添加浓度为2.0mg/L。不同浓度2,4-D 与6-BA 组合的胚状体平均分化率为8.4%,与KT 组合的胚状体平均分化率为9.4%,诱导培养基I7 分化率最高,2,4-D添加浓度为2.0mg/L、KT 为0.5mg/L。

图3 2 种不同类型愈伤组织Fig.3 Two different types of calluses
2.4 不同分化培养基对胚性愈伤组织/体胚苗的影响
将添加2.0mg/L 2,4-D 组合0.5mg/L KT 的培养基诱导出的完全脱分化愈伤块转入分化培养基,如图4 所示,5 个分化培养基胚状体分化率均高于CK,其中D4 的分化率(42.7%)显著高于其他处理(P<0.05),D5 的分化率最低,略高于CK(P>0.05)。
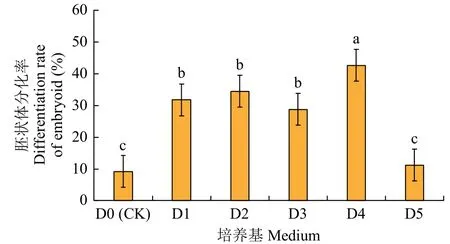
图4 不同分化培养基内胚状体/体胚苗的分化率比较Fig.4 Comparison of differentiation rates of embryo/seedlings in different differentiation media
2.5 再生苗的诱导分化培养
将以上分化出的再生苗经生根、壮苗培养(1/2 MS 培养基+0.1mg/L IBA)后扩繁,扩大再生苗数量。以再生苗的幼嫩叶片和叶柄为外植体,以不同浓度的2,4-D(1.0~8.0mg/L)组合0.5mg/L 6-BA、KT 培养基诱导,20d 后发现外植体均能100%诱导出胚性愈伤组织(黄绿色,有绿点),在不添加激素的分化培养基(1/2 MS)也分化出大量的胚状体,且能分化为无根小苗,剥离出后在壮苗培养基里可发育成完整植株。
3 讨论
3.1 不同来源外植体对苜蓿愈伤组织诱导及胚状体分化的影响
紫花苜蓿体胚诱导途径中常见的外植体有下胚轴、子叶、茎段、叶柄和根等,通常选择生理代谢旺盛、分化程度较低的离体组织。下胚轴分化能力相对较强,是最佳外植体[11-12]。以子叶为外植体,可成功建立苜蓿高频再生体系[13-14]。本研究发现,下胚轴和茎段出愈早于叶片,完全脱分化时间短于叶片2~4d,但经24d 诱导后,都能100%脱分化。不同外植体脱分化后的愈伤组织形态和质地没有明显区别。但有研究[10-15]显示,体胚分化能力与外植体来源有关。本研究中,3 种外植体的愈伤组织转入不添加任何激素的分化培养基后,分化率都较低,最终分化出胚状体的愈伤组织很少,多数都失活并褐化死亡,无法比较不同外植体的体胚分化能力。
3.2 2,4-D 与不同细胞分裂素组合对苜蓿愈伤组织诱导及体细胞胚发育的影响
2,4-D 浓度是影响苜蓿愈伤组织诱导率的主要因素[16]。不添加2,4-D(CK)的培养基苜蓿诱导率为0.0%,而添加2,4-D 的培养基都有愈伤组织出现。当2,4-D 浓度为2、4mg/L 时,诱导率达100.0%;当浓度为8mg/L 时,诱导率最低,愈伤块体积最小,开始褐化时间也早,说明高浓度的2,4-D 对苜蓿愈伤组织的诱导有抑制作用,这个结论与史毅等[10]研究结果一致。
2,4-D 在苜蓿愈伤组织的诱导和体细胞胚早期发育过程中起关键作用,且常与低浓度细胞分裂素配合使用[11]。王英哲等[13]研究显示,2,4-D 与KT 组合能分化出胚状体。张凌云等[7]认为,2,4-D与6-BA 组合是最佳诱导培养基。李玉珠等[4]研究显示,在相同浓度2,4-D 培养基上,有的品种添加KT 能诱导出胚状体,而有的品种添加6-BA 有胚状体出现。本研究中,当2,4-D 浓度为1.0~5.0mg/L时,与0.5mg/L 6-BA 或KT 组合,都有胚状体产生,但胚状体分化率都较低,故有必要开展分化培养条件研究。
3.3 不同分化培养基对胚状体分化的影响
高浓度的2,4-D 对苜蓿胚发生存在抑制作用[17],故通常在愈伤组织脱分化后降低2,4-D 浓度或不添加。本研究发现,5 个添加激素处理(3个添加KT 组合NAA,1 个只添加KT,1 个添加6-BA 组合NAA)分化率都高于不添加激素的1/2 MS 培养基对照,说明添加低浓度的KT、6-BA 与NAA 组合以及单独添加低浓度KT 对苜蓿体胚分化都有促进作用。细胞分裂素对促进体细胞胚成熟及进一步分化有显著作用[18]。KT 有促进组织细胞分裂和分化不定芽的作用[8],但KT 的添加不利于愈伤组织分化[19]。以上研究结果的不同可能与诱导愈伤组织的基本培养基、培养条件、苜蓿品种等因素有关。苜蓿体胚分化依然存在再生率不稳定和试验重复性差等问题,应根据不同苜蓿品种开展相应的分化培养条件、激素种类和浓度等研究。
3.4 再生苗对高频再生体系建立的影响
以再生苗为外植体,开展愈伤组织的诱导和分化,发现叶片和叶柄在不同浓度2,4-D(1.0~8.0mg/L)和低浓度6-BA、KT 组合下,在不添加激素的1/2 MS 培养基上均能100%产生体细胞胚,经剥离后都能发育成完整植株。从愈伤组织的诱导到完整植株的出现,与初代相比,再生周期缩短,植株更健壮,说明基因型是影响苜蓿体胚再生植株形成的最关键因素。这也验证了Sairam 等[20]关于苜蓿愈伤组织的再生能力受基因型控制且高度遗传的结论。由此可见,建立一个苜蓿品种稳定高频的体细胞胚胎再生途径,可先诱导、分化出少数再生植株,再进行大量扩繁,从而以再生植株的外植体建立再生体系。
4 结论
以苜蓿地方品种固原紫花为材料,通过2,4-D与不同细胞分裂素组合及胚状体分化培养基的比较筛选,得出胚性愈伤组织诱导率最高的培养基为MS 基本培养基+2.0mg/L 2,4-D+0.5mg/L KT,诱导率为12.6%;分化率最高的培养基为1/2 MS+0.4mg/L KT,分化率为42.7%。以分化出的再生苗开展诱导、分化,很容易建立稳定的体胚高频再生体系,可见苜蓿再生能力高度遗传,高度依赖基因型。
