水稻粉质胚乳突变体cse 的表型分析及基因定位
2023-10-13花芹林泉祥宋远辉孙家猛张祖普陈庆全李金才张海涛
花芹 林泉祥 宋远辉 孙家猛 张祖普陈庆全 李金才 张海涛
(1 安徽农业大学农学院,230036,安徽合肥;2 江苏红旗种业科技有限公司,225311,江苏泰州;3 江苏省(红旗)稻麦良种繁育工程技术研究中心,225311,江苏泰州)
水稻(OryzasativaL.)作为重要的粮食作物,其胚乳主要由近80%淀粉和10%贮藏蛋白组成,为人类日常膳食提供重要能量。淀粉的生物合成和贮藏蛋白积累直接影响稻米的外观、产量和食味品质。
胚乳发育中贮藏物质的合成受复杂基因调控。淀粉中ADP-葡萄糖焦磷酸化酶催化葡萄糖-1-磷酸和ATP 而产生活化的葡萄糖基供体ADP-葡萄糖和焦磷酸[1]。随后在歧化酶、淀粉磷酸化酶、可溶性淀粉合酶、颗粒结合淀粉合酶I、淀粉分支酶和淀粉脱支酶等催化协同酶的作用下合成[2]。这些关键酶的缺失有可能导致胚乳皱缩[3]、黏稠[4]、白核[5]、粉质[6]和暗淡[7]等异常表型。其中白核和粉质等突变胚乳常表现为淀粉粒的数量、大小与排列等变异。SSG6编码一种转氨酶同源蛋白,其突变体ssg6成熟核淀粉颗粒增大,但千粒重显著下降且呈轻微粉质状[8];fse1和flo6中出现较小的淀粉粒且排列松散,导致胚乳呈粉质状态[9-10];FLO19编码一个定位在质体的丙酮酸脱氢酶复合物E1 组分亚基α1,该基因突变导致复合型淀粉颗粒减少,单粒型淀粉颗粒增多,导致籽粒内胚乳呈不透明状[11]。除合成淀粉外,粉质胚乳表型也与贮藏蛋白积累有关。谷蛋白合成相关的基因GPA4、GPA5和GPA6发生突变均呈现粉质外观。GPA4编码一个保守膜蛋白GOT1B,通过与Sec23蛋白特异性互作调控COPII(coat protein complex II)组装,积累57kD 的谷蛋白前体,形成2 种异常的Ⅰ型蛋白体,gpa4-1突变体的种子中蛋白含量显著降低[12];GPA5编码一种植物特有的PX(phox-homology)结构域蛋白,与Rab5a 和VPS9a协同互作调控密集囊泡介导的后高尔基体向蛋白贮藏液泡的运输[13];GPA6编码一个Na+/H+反向转运体OsNHX5,gpa6只有部分谷蛋白被转运到II型蛋白体中,导致II 型蛋白体减小[14]。此外,一些基因突变导致淀粉与蛋白合成均受影响,形成粉质胚乳。转录因子OsNAC20和OsNAC26双突变体表现为淀粉和贮藏蛋白含量显著下降,呈现粉质胚乳[15]。FLO2编码一个具有3 个四肽重复基序(TPR)蛋白,TPR 包含2 个反向平行的α-螺旋结构,作为容纳靶蛋白的互补区域,与多个目标蛋白同时发生相互作用[6]。突变体中与FLO2相关的参与淀粉和贮藏蛋白生物合成的许多基因的表达水平显著降低,产生异常胚乳,表现为暗色和粉质特征,胚乳中贮藏物质含量减少,籽粒变小,F1种子育性降低[16-17]。
虽然水稻粉质胚乳成因研究取了一定进展,但相关基因调控网络仍不明晰,仍需进一步发掘相关品质突变体,阐明淀粉和贮藏蛋白的生物合成与调控机制,促进优质水稻育种。本文通过筛选中花11组织培养突变体库,获得一个呈粉质、垩白和皱缩外观胚乳的突变体cse(chalkinessandshrunken endosperm)。对突变体进行表型观察、理化性质测定、遗传分析、基因定位和候选基因克隆等研究,发现cse为flo2的一个新等位变异。
1 材料与方法
1.1 试验材料与定位群体构建
水稻粉质胚乳突变体cse来源于粳稻(Oryza sativaL.ssp.japonica)品种中花11 的组织培养。自然环境下,连续多代自交和田间考察发现,突变性状能够稳定遗传。将其与粳稻品种IRAT129 配制杂交组合获得F1。F1自交获得F2,F2群体用于遗传分析和基因定位。亲本、F1杂交种和F2群体材料均播种于安徽农业大学国家高新技术农业园(合肥),单株种植,株行距为18cm×25cm,田间管理同大田生产。其中亲本各种植8 行,每行10 株,设3 次重复,观察并记录分析水稻各生育期亲本间形态特征及农艺性状差异,并进行单株收获。利用JLGJ-45 型电动砻谷机(大吉光电,杭州)去除颖壳后,统计群体中正常表型单株和突变体表型单株的分离比,选取极端个体用于精细定位。利用SPSS 20.0 软件进行统计分析。
1.2 籽粒干物质积累测定
在中花11 和突变体穗部颖花开花后第3、7、14、21、28d 剪取穗顶部籽粒,置于105℃烘箱杀青30min 后,在80℃恒温干燥箱中烘至恒重,脱除颖壳后测量干重,每个样品设置10 次重复[18]。
1.3 碘染与扫描电镜
亲本种子去稃后用超纯水浸润12h,横切并滴加2%I2-KI 溶液染色,并置于尼康S261-60 型体视镜下观察并拍照记录。扫描电镜观察参照Zhang等[19]方法,使用E1010 型离子溅射仪(日立,日本)在断面镀上碳膜,采用S-4800 型高分辨率扫描电子显微镜(日立,日本)观察并拍照。
1.4 成熟种子的理化性质测定
去壳籽粒采用200T 型高速多功能粉碎机研磨成糙米粉,过100 目筛,50℃恒温箱中烘至恒重备用。采用A1481-1 植物淀粉含量试剂盒测定总淀粉含量;参照标准米质方法NY 147-88 测定直链淀粉含量[20];参照龙武华[21]的方法测定总蛋白含量;参照蒽酮比色法[22]测定可溶性糖含量;采用DA7200 型近红外快速分析仪(Perten,美国)测定糙米中脂肪酸、垩白度、碱消值和胶稠度[23];使用Pyris DSC8000 型差示扫描热量仪(Perten,美国)测定淀粉糊化特性[24]。参照于艳芳等[25]的方法测定尿素膨胀体积。每个样本进行3 次生物学重复。
1.5 目的基因定位与分子标记的开发
SSR 和Indel 分子标记的开发利用Gramene(http://www.gramene.org)和NCBI(https://www.ncbi.nlm.nih.gov)网站数据库下载日本晴和9311序列,通过BLAST 对比插入/缺失位点,利用Primer 3.0(http://bioinfo.ut.ee/primer3-0.4.0)开发分子标记,并由南京擎科生物科技有限公司合成。利用筛选出具有多态性的均匀分布于12 条染色体的197对SSR 和Indel 分子标记。选取F2群体中具有粉质胚乳表型的种子,用3%H2O2浸泡30min 破除休眠后,置于RXZ-50CD 型智能人工气候箱(江南仪器,浙江)培养至三叶期,采用CTAB 法[26]提取叶片总DNA,构建CSE定位的DNA 混池,寻找与目标基因的连锁位点,在此基础上利用F2和F3群体进行目标基因的初步定位。再开发新的标记,扩大定位群体,对目标基因进行精细定位(表1)。10µL PCR 反应体系如下,2×TaqPCR StarMix 5µL、正反向引物(10µmol/L)各1µL、DNA 模板1µL、10%聚乙烯吡咯烷酮(PVP,polyvinylpyrrolidone)1µL 和ddH2O 1µL。PCR 反应条件为94℃预变性2min;94℃变性30s,退火温度55℃~60℃退火30s,72℃延伸1min,共计35 个循环;最后72℃下终延伸5min,4℃保存。PCR 产物经8%非变性聚丙烯酰胺凝胶电泳、0.1%AgNO3染色后在3%的甲醛和NaOH 中显色,观察拍照并记录基因型[27]。

表1 引物信息Table 1 Information of primers
1.6 RNA 提取与Real-time PCR 表达分析
取野生型和突变体三叶期叶片、根、茎、旗叶、花后3、7、14、21 和28d 穗,于-80℃保存,使用多糖多酚植物总RNA 提取试剂盒(擎科)提取总RNA,超微量分光光度计(DENOVIX,美国)检测浓度。RNA 反转录试剂盒(艾瑞科)获得cDNA。利用CSE的CDS 设计荧光定量PCR 引物,水稻基因Ubiquitin(LOC_Os03g13170)作为内参(表1)。反应体系采用莫纳生物科技有限公司试剂盒MonAmpTMChemoHS qPCR Mix(MQ00401),20µL反应体系为0.4µmol/L 的正、反向引物、MonAmpTMChemoHS qPCR Mix 以及适宜浓度的cDNA;在BIO-RAD CFX96TM型荧光定量PCR 仪(BIORAD,美国)上进行扩增反应,反应条件为:95℃预变性10min;95℃变性10s,60℃延伸30s,40个循环。每个样品重复3 次,用2-△△CT法计算CSE的表达量[28]。
2 结果与分析
2.1 突变体表型分析
水稻粉质胚乳突变体cse在合肥经多年种植性状稳定。与野生型相比,cse在营养生长阶段分蘖数显著增加48.0%;生殖生长阶段,生长较缓慢,株高显著降低,主要表现在第3、4 节间分别缩短29.4%和40.0%、穗长缩短7.9%(表2,图1a~c)。考种结果(表2)表明,cse二次枝梗数和结实率分别降低29.7%和3.0%。cse成熟干燥颖果表现为轻微皱缩,胚胎外围呈透明状,内胚乳粉质垩白不透明;而野生型颖果饱满,胚胎轮廓清晰,胚乳硬质呈透明状(图1e~f)。与野生型相比,cse成熟籽粒的粒宽和粒厚显著降低,千粒重仅为野生型的62.0%(图1d,表2)。此外,我们分析了野生型和cse颖果发育过程中干重变化。从授粉后7d 开始,cse千粒重极显著低于野生型,并且随着籽粒的发育千粒重间差异逐渐扩大(图2)。以上结果表明,cse的突变不仅影响植株的发育,也影响籽粒的灌浆速率。

图1 野生型和突变体cse 的表型鉴定Fig.1 Phenotypic characterization of wild type and the cse

图2 野生型和cse 的不同时期干物质积累比较Fig.2 Dry matter accumulation in wild type and cse at different stages

表2 野生型和突变体农艺性状比较Table 2 Comparison of agronomic traits between the wild type and cse
2.2 胚乳淀粉颗粒染色与结构分析
野生型糙米横截面呈透明状且质地紧密,而突变体糙米横截面呈粉质状态且质地疏松。经2%的I2-KI 染色发现,突变体着色较野生型浅(图3a~b),表明cse淀粉含量较野生型更低。利用扫描电镜观察野生型和cse成熟颖果横断面显示,野生型胚乳淀粉颗粒紧密堆积、呈规则的多边形晶体结构(图3c);而cse淀粉粒呈不规则球形且淀粉颗粒间缝隙较大,排列疏松,多以单一、分散的淀粉粒存在(图3d)。上述结果表明,cse对胚乳中淀粉颗粒的发育至关重要。

图3 野生型和cse 的成熟胚乳碘染和扫描电镜观察Fig.3 Iodine staining and SEM observation of mature endosperm between wild type and cse
2.3 成熟籽粒的理化性质分析
由于cse胚乳中淀粉发育异常,我们还检测了成熟籽粒的理化性质。与野生型相比,cse的总蛋白质、脂肪酸和可溶性糖含量分别极显著增加21.5%、83.7%和34.8%,垩白度增加340%;但总淀粉含量降低22.2%,且直链淀粉含量(8.8%)仅为野生型(17.6%)的49.9%(表3)。直链淀粉含量通常与糊化温度、胶稠度相关。cse直链淀粉含量降低,其起始糊化温度、峰值、结束温度和焓变量也分别显著降低6.9%、5.1%、3.1%和28.3%(表4),糊化温度的关键指标碱消值也相应降低6.2%,但胶稠度增加11.2%。糊化特性影响米粉溶解性,尿素膨胀试验结果表明,两者在5mol/L 出现明显膨胀现象(图4),但突变体的米粉膨胀体积变化始终低于野生型。以上结果说明,cse突变影响了水稻颖果中淀粉和贮藏蛋白的累积,进而改变胚乳淀粉的理化性质。
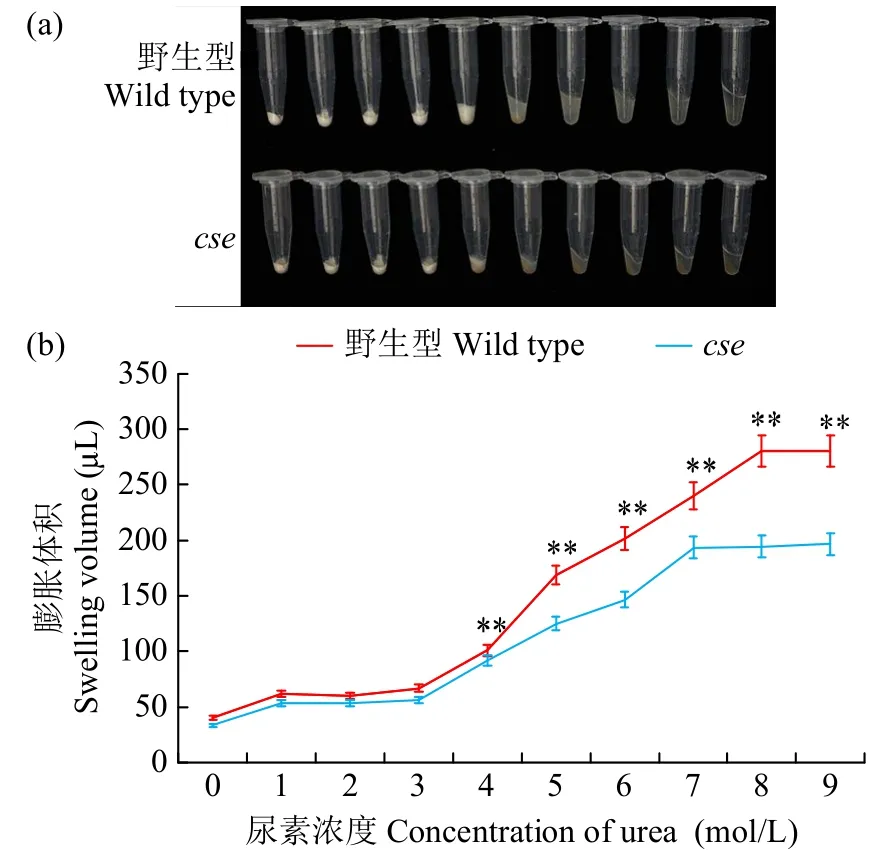
图4 野生型和cse 的尿素膨胀分析Fig.4 The swollen volume of wild type and cse starch in urea solutions of various concentrations
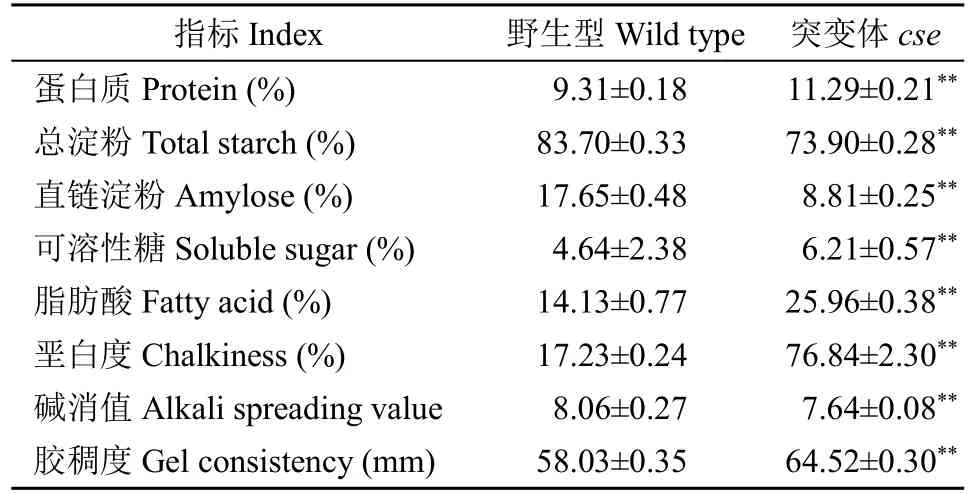
表3 野生型和突变体理化性质比较Table 3 Physicochemical properties comparison between the wild type and cse
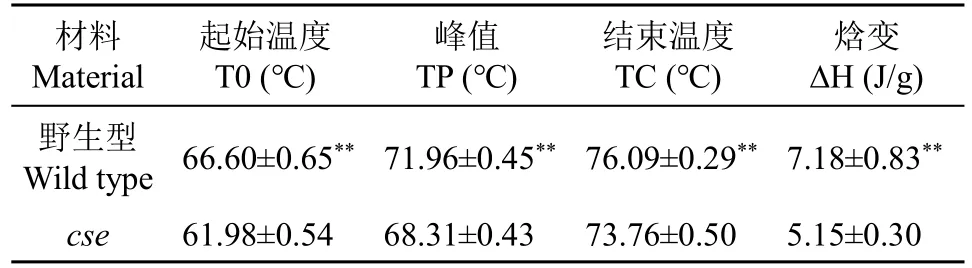
表4 野生型和突变体糙米粉的热特性参数Table 4 Thermal parameters of the wild type and cse
2.4 cse 遗传分析
以cse为母本,以粳稻品种IRAT129 为父本配制杂交组合,F1籽粒发育正常,F2籽粒出现正常表型和粉质胚乳表型分离。2020 年随机调查F2群体212 株,2021 年随机调查F2群体743 株(表5),cse符合3:1(χ22020、2021<χ20.05,1=3.84)的分离规律,推断cse的表型性状受1 对隐性核基因控制。
2.5 粉质胚乳基因的精细定位
利用2 个亲本cse和IRAT129 间具有多态性的197 对分子标记,对F2极端隐性表型单株构成的DNA 混池进行连锁分析,发现亲本cse的带型与位于第4 号染色体上标记Os4-10 一致(图5a)。进一步利用Os4-10 附近的标记进行连锁分析,并开发了另外5 对多态性引物Os4-11、Os4-12、Os4-12-11、Os4-13 和Os4-13-16,对550 株F2具有粉质胚乳外观单株进行初定位,发现目标基因cse位于分子标记Os4-13 和Os4-13-16 之间(图5b)。基于Gramene 中水稻基因序列信息,在Os4-13 和Os4-13-16 之间开发出4 对新的SSR 和Indel 标记(表2),借助3034 株F2隐性单株,将cse锁定在物理距离为40kb 的Os4-14-8 和Os4-15 标记间(图5c)。
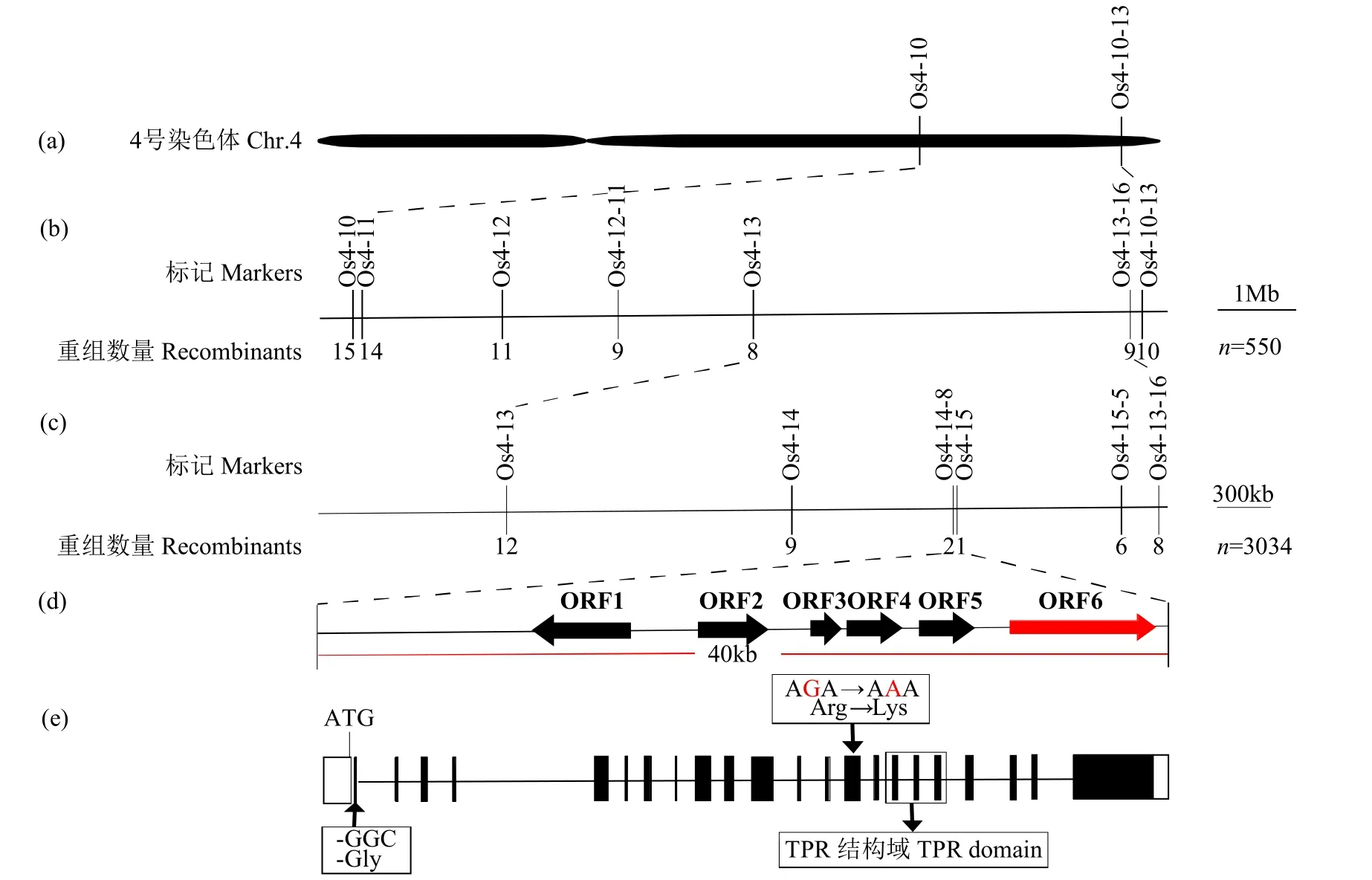
图5 cse 的精细定位Fig.5 Fine mapping of cse
Gramene(http://www.gramene.org/)网站和Rice Genome Annotation Project(http://rice.plantbiology.msu.edu/cgi-bin/gbrowse/rice/)上的基因预测信息表明上述40kb 候选区间内共有6 个候选基因(图5d,表6)。经反复测序发现,与野生型相比,cse的ORF1~5 不存在碱基序列差异,但ORF6(LOC_Os04g55230)的第1 个外显子处(-ATG 51bp)缺失3 个碱基(GGC),导致甘氨酸缺失;第14 个外显子处(-ATG 7887bp)碱基G 被替换成A,导致精氨酸突变为赖氨酸(图5e)。cse编码含有四肽重复结构域的蛋白质,是已经报道[6,21]基因OsFLO2/OsCNY8的新等位基因。

表6 精细定位区间内注释基因Table 6 Annotated genes in the fine mapping range
2.6 CSE 表达模式与同源分析
我们利用Phytozome13(https://phytozome.jgi.doe.gov)上LOC_Os04g55230序列信息,在水稻、拟南芥、小麦、玉米和大麦的基因组数据库中搜索同源蛋白,利用MEGA7.0 软件对CSE 及同源蛋白的氨基酸序列进行比对并构建系统进化树,发现水稻中存在2 个未有相关研究报道的同源基因LOC_Os07g23990和LOC_Os02g15660,同源率分别为73.5%和72.2%。拟南芥中同源基因ATFLO2(AT1G15290)参与种子中贮藏物质的积累,其T-DNA 插入突变体表现为种子易碎、数量减少以及千粒重降低[29];ATFLL1(AT1G01320)参与叶片中淀粉合成和糖代谢[30]。因此,表明CSE是粉质胚乳相关基因(图6)。

图6 CSE 和CSE 类蛋白的系统发育分析Fig.6 Phylogenetic analysis of CSE(LOC_Os04g55230)and CSE-like proteins
利用Real-time PCR 检测CSE在野生型中的表达模式,结果(图7)表明,CSE在检测的水稻组织包括叶片、根、茎、旗叶和不同时期发育的胚乳中均有表达,在旗叶中大量表达,授粉后14d 的发育胚乳中表达量达到峰值。
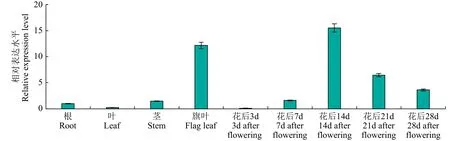
图7 CSE 在野生型不同组织及不同发育时期胚乳的表达模式分析Fig.7 Wild type expression analysis of CSE in different tissues and endospern of different growth periods
3 讨论
水稻胚乳的淀粉生物合成和贮藏蛋白积累是一个复杂而精密的生物过程,涉及一系列酶促反应。淀粉合成受到抑制,导致胚乳细胞淀粉颗粒排列疏松,颗粒间的缝隙引起光的散射,光在穿过胚乳损失而表现为白色不透明[31]。已有多个胚乳发育基因突变导致胚乳不透明的研究报道,如FLO5、WAXY和BEIIb[5,32]。胚乳缺陷突变体是阐明种子发育和淀粉生物合成分子机制的宝贵资源。本研究中,我们分离并鉴定了籽粒轻微皱缩、胚乳外围轮廓清晰透明及内胚乳粉质不透明表型的cse突变体。与野生型相比,cse籽粒灌浆速率下降、结实率、千粒重和淀粉含量降低,淀粉的糊化特性和结构发生改变。扫描电镜观察显示,cse横截面质地疏松,淀粉粒呈不规则球形,淀粉颗粒间缝隙较大、多以单一、分散的形式存在且碘染色力较弱;而野生型胚乳淀粉颗粒紧密堆积,呈规则的多边形晶体结构(图3)。cse种子的横截面外观与flo7和flo5/ss3a突变体相似,flo7仅在籽粒外围表现出粉状的白色胚乳,而flo5/ss3a突变体则表现出白核粉状胚乳[5,7]。淀粉合成由多个基因协同调控,因此,cse的粉质胚乳表型可能是水稻淀粉合成相关基因表达改变的共同作用。TPR 结构域广泛存在于多种蛋白质中,并在蛋白质折叠、蛋白质运输、细胞周期控制和翻译后修饰等方面发挥关键作用,如编码TPR 结构域的基因OsST2,对水稻分蘖起调控作用[33-34]。cse的第16、17、18 外显子编码3 个TPR基序,但第14 个外显子处存在一个碱基G 被替换成碱基A,导致精氨酸突变为赖氨酸,可能造成TPR 结构域功能缺陷,使总淀粉含量降低22.2%,直链淀粉含量仅为野生型的49.9%(表3)。由此推测,cse可能对淀粉的合成与积累产生影响。此外,flo2仅籽粒大小和淀粉品质降低[6],而cse还伴有植株变矮和分蘖数增加等性状,表明cse是flo2的新等位基因突变体。
脂质通过调节代谢产物转运活性间接影响淀粉粒的形成。编码单半乳糖基二酰基甘油(MGDG)合酶的o5点突变导致MGDG 和半乳糖基二脂酰基甘油(DGDG)减少,导致胚乳发育和淀粉合成异常[35]。此外,磷脂酸被证实与拟南芥中涉及半乳糖代谢和转运的几种蛋白结合,包括ABC 脂质转运蛋白和DGD1[36]。粉质胚乳突变体fse1中总DGDG含量显著降低,但总MGDG 含量有降低的趋势[9]。cse胚乳发育过程中,脂肪酸含量显著增加83.7%。cse脂质组成的改变可能导致种子发育异常,进而造成淀粉合成异常。因此,脂质介导的代谢物转运可能是揭示种子淀粉积累机制的重要途径,进一步对淀粉体中淀粉合成和膜脂合成之间的功能联系研究,有助于阐明这种联系背后的分子机制。
4 结论
cse是一个从组织培养后代植株中筛选出的胚乳发育异常突变体,该突变体籽粒受粉7d 后灌浆速率显著降低,成熟籽粒表现为轻微皱缩,胚胎外围呈透明状,内胚乳粉质不透明。扫描电镜结果显示,淀粉粒呈不规则球形且淀粉颗粒间缝隙较大,排列疏松,多以单一、分散的淀粉粒存在且碘染色较浅。对籽粒理化性质测定表明,突变体的蛋白质、脂肪酸和可溶性糖含量显著增加,总淀粉和直链淀粉含量降低。通过cse和IRAT129 配制杂交组合的F2遗传定位群体分析表明,该性状是由1 对隐性核基因控制。通过图位克隆将该基因定位于4 号染色体Os4-14-8 和Os4-15 标记之间,物理距离为40kb。测序表明,候选区间内ORF6(LOC_Os04g55230)第1 个外显子缺失3 个碱基(GGC)导致甘氨酸缺失,第14 个外显子内存在一个由G 到A 的突变,导致精氨酸突变为赖氨酸。cse编码的氨基酸改变,其蛋白功能活性受损,引起粉质胚乳表型。
