水稻加工和外观品质性状QTL鉴定
2023-09-14姚晓云陈春莲熊运华黄永萍彭志勤刘进尹建华
姚晓云 陈春莲 熊运华 黄永萍 彭志勤 刘进 尹建华
水稻加工和外观品质性状QTL鉴定
姚晓云 陈春莲 熊运华 黄永萍 彭志勤 刘进 尹建华*
[江西省农业科学院 水稻研究所/水稻国家工程研究中心(南昌)/国家水稻改良中心南昌分中心, 南昌 330200;*通信联系人, email: jxyjh2011@163.com]
【目的】稻米加工与外观品质是影响水稻生产效益和商品价值的关键因素,发掘相关性状主效QTL有助于进一步阐明水稻品质性状的分子遗传机制,也可为水稻优质育种提供材料。【方法】以优质粳稻龙稻5号为母本和高产籼稻中优早8号为父本杂交衍生的重组自交系群体为试验材料,在3个环境条件下对稻米加工和外观品质进行性状比较和QTL分析。【结果】检测到37个与加工和外观品质相关的QTL,分布在12条染色体上,三个环境分别发现15、15和21个QTL,其中仅10个QTL在2个环境中稳定表达,2个QTL在3个环境中重复检测到。加工和外观品质相关QTL具有明显的遗传重叠效应,第2、6、7、10和11染色体上存在主效QTL簇,对加工和外观品质存在明显的调控作用。【结论】第7染色体RM1306−RM420区间存在一个新的QTL(),该区间对垩白粒率和垩白度具有较强的调控效应;此外,上位性互作也是调控加工和外观品质的重要组成部分,主效QTL参与上位性互作效应。
水稻;加工品质;外观品质;QTL分析;
我国是水稻生产和消费大国,高产优质水稻生产关乎国家口粮安全和百姓福祉,不断提高稻谷产量和品质对于保障国家粮食安全和社会稳定具有重要意义[1-2]。稻米加工品质包括糙米率、精米率和整精米率,整精米率是重要加工品质指标;外观品质包括粒型、垩白和透明度等指标,垩白性状是外观品质主要因子。整精米率和垩白性状直接决定稻米外观形态和商品价值。垩白过高将导致整精米率降低和米饭适口性变差,少垩白、高整精米率的长粒型品种具有较高商品价值[2-3]。加工和外观品质主效QTL的发掘将有利于全面阐明稻米品质分子调控机制,也将为优质稻育种提供优异基因资源。
加工和外观品质性状均属于多基因控制的数量性状,受环境影响较大,基因型与环境互作效应是影响加工和外观品质表型的重要因素[4-7]。以往研究者对加工和外观品质开展了广泛的遗传定位研究,已有多个相关QTL被鉴定,仅有少数几个稳定表达的主效QTL被精细定位或克隆,主要包括、、、、、(http://www.gramene.org/, http://www.ricedata.cn/gene/)。Ren等[8]在第10染色体长臂末端定位了一个调控糙米率的主效QTL,其39.5 kb目标区间包含6个候选基因;Wu等[9]发现一个调控心白率的基因,该基因具有降低心白率、提高产量和食味值的功能;Li等[10]克隆了一个垩白主效基因,该基因编码液泡H+-焦磷酸转移酶,调节内膜转运系统pH平衡,增加蛋白质体数量,导致淀粉粒无法紧密排列而形成垩白;邱先进等[11]利用全基因组关联分析方法,检测到86个垩白显著关联位点,其中第5和12染色体SNP对垩白具有较高调控效应;Wang等[12]克隆了一个编码细胞壁转化酶基因,调控籽粒灌浆期碳源分流,影响储藏物质累积速度,导致籽粒形成垩白;Cai等[13]鉴定了一个垩白调控基因,突变体表现为白心胚乳且灌浆有缺陷,粒重、单株产量和淀粉含量显著降低,淀粉理化特性发生改变,其正常的复合淀粉颗粒显著减少,具有更多的单一淀粉粒填充了胚乳细胞。此外,还有4个垩白主效QTL[14]、[15]、[16]和[17]被精细定位。目前,尽管稻米加工和外观品质分子调控机理取得了一定进展,但相关分子遗传调控和环境响应机制仍不甚清楚,诸多问题仍亟待解决[18-22]。加工与外观品质的形成受多基因调控,且环境及基因-环境互作效应对整精米率和垩白的形成存在明显影响。因此,发掘更多环境特异性或钝感基因将为稻米品质改良奠定坚实基础。
本研究以龙稻5号(优质粳稻)和中优早8号(高产籼稻)杂交衍生的RIL群体为试验材料,在典型双季稻区江西省早季、晚季及海南三个生态环境下鉴定稻米加工和外观品质表型及QTL,以期发掘稳定表达的主效位点,为稻米品质遗传改良提供基因资源。
1 材料与方法
1.1 试验材料与田间试验
以优质粳稻龙稻5号(Longdao 5,LD5)为母本和高产籼稻中优早8号(Zhongyouzao 8,ZYZ8)为父本杂交衍生的重组自交系(recombinant inbred lines,RIL)群体为试验材料,共180个株系(F9)。亲本和RIL群体于2019年早季(E1)和晚季(E2)种植于江西省农业科学院高安综合试验基地,2020年种植于江西省农业科学院海南基地(E3),单苗栽插,每系4行,每行8株,田间管理同当地大田栽培管理。
1.2 性状测定
成熟后收获整个小区稻穗,晒干后放置15 d脱粒,常温存储3个月。根据农业部标准NY147−88《稻米品质的测定》(稍作修改)进行性状测定。称取100 g稻谷,碾磨成糙米和精米,测定糙米重和精米重,采用万深SC-E外观判别仪测定整米率、长宽比、垩白粒率和垩白度。糙米率、精米率和整精米率分别依据以下公式进行计算:糙米率(%)=糙米重(g)/稻谷重(g)×100;精米率(%)=精米重(g)/稻谷重(g)×100;整精米率(%)=整米率×精米率。设置2次重复,取其平均值进行统计分析。
1.3 遗传图谱构与QTL分析
遗传图谱包含223个遗传标记(SSR、InDel),图谱覆盖基因组1514.0 cM,标记间平均距离为6.79 cM,平均每条染色体标记数为18.58个,RIL群体和遗传图谱由刘进等[23]构建完成。采用QTL ICI Mapping 4.2的完备区间作图(ICIM)方法[24]进行加性效应、加性×加性上位互作效应QTL分析,LOD的阈值设为2.50。当实际求得的LOD值大于2.50时,就判定该区段存在1个QTL,并估算每个QTL的加性效应值和贡献率;应用ICIM-EPI进行上位性分析,LOD值为4.50。QTL的命名遵循McCouch等[25]的原则。
2 结果与分析
2.1 表型分析
不同生态环境下,双亲加工和外观品质性状(除糙米率外)存在显著差异,龙稻5号均表现为短圆粒,具有较高的糙米率、精米率和整精米率,垩白粒率和垩白度较小,而中优早8号均表现为长粒,糙米率、精米率和整精米率较低,垩白表型明显。RIL群体不同家系间加工和外观品质性状存在较大幅度变异,呈现双向超亲分离,近似于连续的正态分布。这表明,加工和外观品质性状均为多基因控制的数量性状,符合QTL作图的要求(表1)。
RIL群体加工和外观品质性状间存在显著相关性,不同生态环境下,相关系数存在一定差异。糙米率与精米率、整精米率、长宽比、垩白粒率(除晚稻)显著或极显著相关;精米率与整精米率、垩白粒率和垩白度极显著相关,其中精米率与整精米率极显著正相关,与垩白粒率和垩白度间极显著负相关;长宽比与精米率、整精米率相关性不显著,与垩白粒率、垩白度极显著负相关,垩白粒率与垩白度性状极显著正相关(表2)。这表明加工品质与外观品质各性状关系紧密、相互影响,各性状表型受环境与基因型互作效应共同调控。
2.2 QTL分析
检测到8个影响加工品质的QTL,3个环境下分别检测到2、2和6个,分布于第1、2、4、7、10和11染色体上,LOD值为2.52~4.01,解释5.00%~11.80%的表型变异,仅2个QTL在不同环境下稳定表达,其余QTL均仅在单一环境下被检测到(表3)。这些调控加工品质的QTL中,1个为调控糙米率的QTL,影响晚稻环境下的糙米率,加性效应值较小,为微效QTL;2个为精米率的QTL,位于第2和7染色体,其中贡献率较大,来自LD5等位基因具有增加精米率效应;5个为控制整精米率的QTL,分别位于第2、4、7、10和11染色体,其中和在两个环境下重复检测到,增效等位基因来自LD5,具有较强调控整精米率的效应。
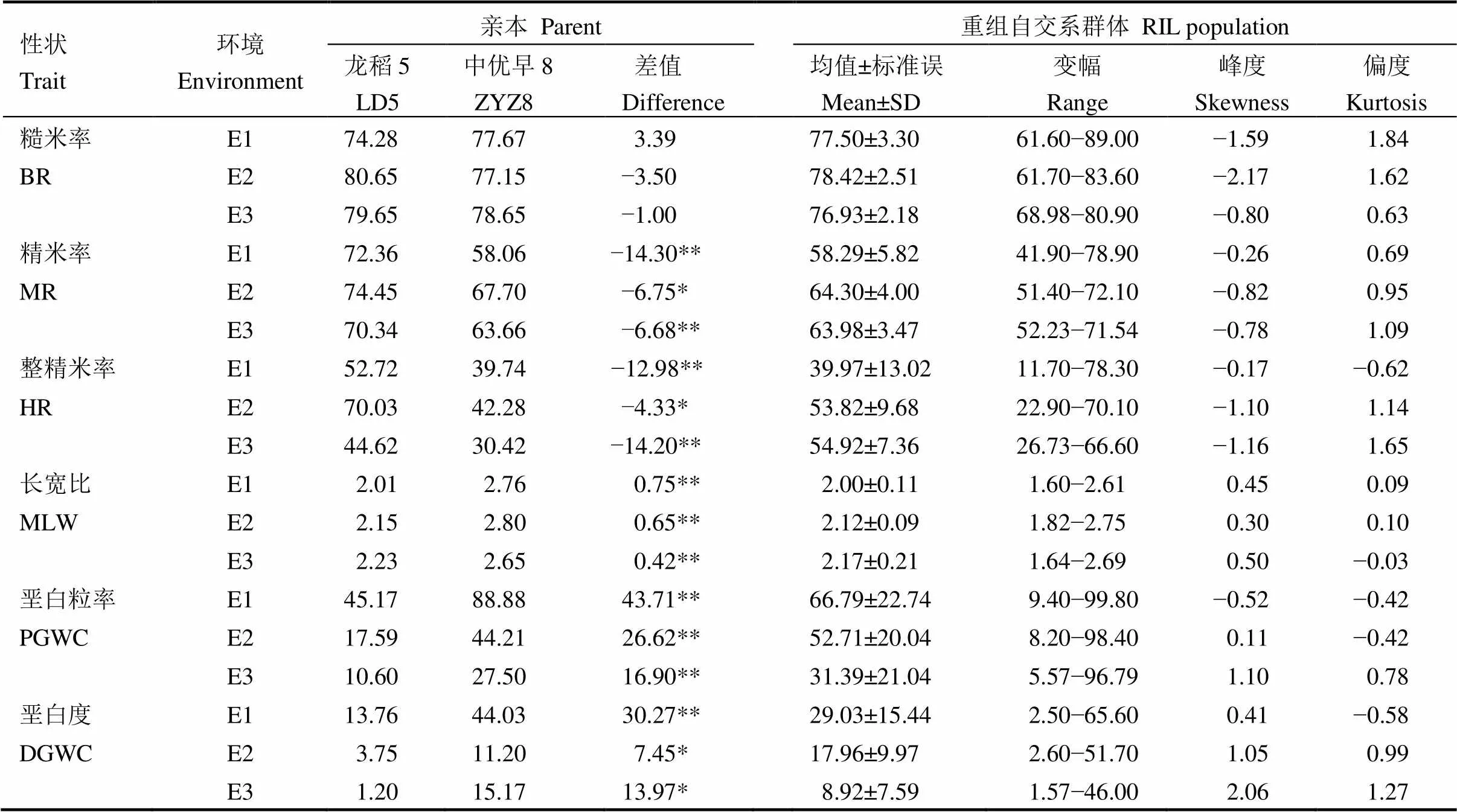
表1 亲本和RIL群体加工和外观品质表型分析
*和**分别表示0.05和0.01水平上差异显著。E1−2019年江西早季;E2−2019年江西晚季;E3−2020年海南。
E1, Early cropping season of 2019 in Jiangxi; E2, Late cropping season of 2019 in Jiangxi; E3, Hainan, 2020. LD5, Longdao 5; ZYZ8, Zhongyouzao 8; BR, Brown rice rate, MR, Milled rice rate; HR, Head milled rice rate; MLW, Milled length/width; PGWC, Percentage of grains with chalkiness; DGWC, Degree of grains with chalkiness. * and ** represent significant difference at the 5% and 1% level, respectively.
检测到28个外观品质相关的QTL,3个环境下分别检测到12、13和15个,分布于第1~8、第10~12染色体上,LOD值为2.50~12.89,解释3.33%~23.07%的表型变异,10个QTL不同环境下稳定表达(表3)。这些影响外观品质的QTL中,10个为调控长宽比的QTL,解释3.33%~23.07%的表型变异,加性效应值为−0.09~0.13;其中和在3个生态环境下稳定表达,和在两个环境下被检测到,其余QTL均在单一环境下表达。9个为控制垩白粒率的QTL,位于第1~4、6、7、10和11染色体上,解释4.61%~12.02%的表型变异,其中和在两个环境下重复检测到,来自LD5的等位基因具有明显降低垩白粒率的效应,其余位点均仅在单一环境下被发现。9个为影响垩白度的QTL,其中、和在不同环境下重复检测到,其余QTL均仅在单一环境中表达;表型贡献率和效应值较大,分别为21.25%和8.23,来自ZYZ8的等位基因具有明显增加垩白度的功能。研究还发现,加工和外观品质QTL存在明显成簇分布的现象,分别分布于第2、3、6、7、10和11染色体,形成6个主效QTL簇(图1),即存在多个加工与外观品质(milling and appearance quality,MAQ)的主效QTL染色体区间,分别为(、和)、(和)、(、)、(、)(、、、)和(、、)。

表2 不同环境下稻米加工和外观品质性状间相关性
*和**分别表示相关性达5%和1%的显著水平。Coefficients marked with *, and ** mean significant correlation at 5% and 1%, respectively.

表3 不同环境下水稻加工和外观品质性状QTL分析

表4 稻米加工和外观品质上位性互作效应分析

图1 稻米加工品质和外观品质相关QTL的染色体分布
Fig. 1. Location of QTLs for milling and appearance quality traits on the genetic map.
2.3 上位性分析
利用ICIM-EPI模型检测加工和外观品质上位性效应位点,共检测到25对上位性互作QTL,分布于第1~12染色体上(表4)。这些上位性位点中,4对为调控糙米率的上位性互作QTL,第3染色体R3M10−STS3.3()区间与第1、4和9染色体相应区段存在明显互作效应;3对为影响整精米率的上位性互作位点,第3染色体STS3.3−STS3.4()与第5、11染色体相应区间存在明显互作效应,第4染色体上的RM5688−RM471区间与第6染色体上的RM6395−RM5814()区间存在明显上位性互作效应,贡献率和效应值分别为12.16%和8.67;2对为调控长宽比互作位点,其中位于第5染色体的R5M13−RM3476区间与第8染色体RM8018−RM1376区间存在明显上位性互作效应,贡献率为12.06%;8对为影响垩白粒率上位性互作位点,分别位于第1、2、3、4、5、6、7、8和11染色体,第6染色体R6M14−RM3827()区间对第3、5染色体相应区间具有明显的上位性效应;8对为调控垩白度上位性位点,其中第3、4、7和12染色体相应区间对第1、2、4、6、8、11和12染色体区间具有明显上位性效应,且多个加性效应QTL具有上位性互作效应。这表明RIL群体中存在多个调控加工和外观品质的上位性互作QTL,上位性互作是调控加工和外观品质表型的重要组成部分,加性效应QTL对其他染色体区间具有重要互作调控效应。
2.4 垩白主效QTL初步验证
结合http://qtaro.abr.affrc.go.jp/网站信息和已发表文献对不同生态环境下多效性QTL簇进行整合分析,发现(、)和(、)为新鉴定的主效QTL簇,其中不同环境下稳定表达()被初步锁定在第7染色体长臂末端RM3555−RM420标记之间,来自LD5的等位基因具有明显降低垩白粒率和垩白度的功能。基于目标QTL定位区间分子标记对RIL群体180个株系进行目标基因分型,不同基因型垩白性状存在明显差异,基因型为-ZYZ8的株系垩白粒率和垩白度显著高于基因型为-LD5的株系,杂合型株系垩白表型值介于两亲本基因型株系之间,目标QTL具有较强的显性效应(图2)。这表明目标株系局部染色体区间基因型与表型存在明显相关性,主效QTL对垩白粒率和垩白度具有较强的调控效应。根据垩白性状在RIL群体中筛选极端表型株系,获取高垩白和低垩白株系各12个,利用目标区间局部基因代换分析将主效QTL定位在第7染色体RM1306−RM420标记之间,物理距离约为484.29 kb;利用RGAP和RAPD数据库对目标区间进行候选基因分析,共查询到71个注释基因,包含11个与胚乳淀粉合成、糖代谢、转录因子及代谢激酶相关的基因,其中和是最为可能的候选基因(表5)。

A和B为不同基因型株系垩白粒率和垩白度表型比较分析;C为主效QTL qChlak7极端株系鉴定与验证,灰色和白色表示龙稻5号(LD5)和中优早8号(ZYZ8)的基因型;G1~G7表示RIL群体株系的7种基因型,n表示每种基因型株系数;C中右图白色和灰色柱子分别表示7种基因型的垩白粒率和垩白度表型值,CK为LD5数值。**表示与对照存在极显著差异(P<0.01),ns表示无显著差异(P>0.05)。
Fig. 2. Genotyping of the major QTL,, located on the chromosome target regions using the extreme lines.

表5 垩白主效QTL qChlak7目标区域关键候选基因分析
加粗与*表示调控垩白性状主效QTL显著关联的关键候选基因。
Bold and * indicate the key candidate genes for significant association of the major QTLfor chalkiness traits.
3 讨论
稻米加工品质和外观品质是品质性状的重要组成,包括糙米率、精米率、整精米率、长宽比、垩白粒率和垩白度等指标,是稻米商品性最为直观的视觉体现,直接决定稻米商品性与市场价值[26-28]。稻米加工和外观品质性状属于多基因控制的数量性状,受基因型和环境效应共同决定,不同基因型品种对生态环境和栽培条件具有明显响应[29-31]。灌浆期温度对稻米品质影响很大,垩白性状对温度最为敏感,高温会缩短有效灌浆时间,导致淀粉及其他有机物质积累减少,透明度变差,垩白增加[32]。徐富贤等[33]研究表明,气象因子对稻米品质具有明显影响,随着播期推迟,糙米率、整精米率、长宽比、蛋白质指标呈增加趋势,垩白度、垩白粒率、胶稠度则呈下降趋势;王东明等[34]研究表明,CO2浓度升高对加工和外观品质存在明显影响,不同基因型品种响应规律存在明显差异;气温、昼夜温差、光照长短对整精米率、垩白、蛋白质和胶稠度性状存在明显影响[35-36]。本研究以RIL群体为试验材料,在高安早季(灌浆期温度较高,灌浆时间缩短)、晚季(对照)和三亚(灌浆成熟期日照短)三个生态环境鉴定加工和外观品质性状,不同生态环境下双亲和RIL群体加工和外观品质性状指标均存在明显差异,不同基因型株系对环境的响应存在明显差异,部分株系对环境钝感,表型变化较小,部分株系对环境敏感,不同环境下表型变化较大,这是导致加工和外观品质QTL鉴定发掘的难点。研究还表明,加工和外观品质各性状间关系紧密、相互影响,垩白与其他品质性状存在显著或极显著相关性;以江西晚稻为对照,江西早季高温环境导致整精米率降低和垩白显著增加,海南短日照环境下加工和外观品质性状发生变化,差异未达到显著水平。
近年来,研究者对稻米加工和外观品质进行了广泛的研究,已发现较多QTL位点,不同环境或群体鉴定的QTL存在明显差异,稳定表达的共性QTL较少[4, 37-38]。翁建峰等[37]利用CSSL群体对加工品质进行QTL鉴定,检测到30个稻米加工品质相关的QTL,仅、、和在不同环境中稳定表达;胡霞等[39]研究表明,水稻碾磨品质与产量性状相关不显著,检测到15个调控碾磨品质相关位点,其中、、和能在不同环境下共表达,还发现整精米率受环境影响较大;周勇等[40]采用CSSL群体对糙米率进行QTL定位,在第6和9染色体上检测到2个主效QTL;Ren等[5]鉴定了4个糙米相关的QTL,分布于第1、8、9和10染色体,能在不同环境下稳定表达,具有明显调控效应,最终将其定位在39.5 kb内;王小雷等[41]在4个环境下共检测到44个QTL,成簇分布于第2、3、5、6和10染色体上,6个QTL在多个环境下被重复检测到,其中第1染色体和第12染色体是两个新的稳定表达QTL。本研究在高安早季、晚季和海南三种生态环境下检测到36个与加工和外观品质相关的QTL,分布在第1~8, 10~12染色体上,在三个环境下分别发现14、15和21个QTL,仅10个QTL在2个环境下稳定表达,2个QTL在3个环境中重复检测到,占总QTL数的27.78%和5.56%,其余QTL均仅在单一环境表达。这表明加工和外观品质由加性效应QTL及互作效应和环境效应形成网络共同调控,不同遗传群体和试验环境下检测到的相关QTL数量和染色体分布存在较大差异,加工品质QTL稳定性较差,不易检测到稳定表达的主效QTL,这与前人研究结果相似[30-32]。同时,本研究还检测到25对上位性互作位点,分布于第1~12染色体上,主效QTL参与上位性互作效应,这也表明上位性互作是调控加工和外观品质的重要组成部分,加工和外观品质表型由环境因素、基因型及各类网络互作共同决定。
研究还表明,加工和外观品质相关QTL成簇分布于第2、3、6、7、10和11染色体,形成6个QTL簇,分别为、、()、()、和。与前人研究比较发现,位于第2染色体的多效性QTL簇与/[42-43]基因的功能和位置相近,可能是粒型基因的多效性效应;位于第10和11染色体的多效性QTL簇和与Ren等[5]、Qiu等[44]定位区段相近,可能与基因多效性有关;和是2个新发现的多效性QTL簇,两者均主要影响垩白性状,其中(、)在不同生态环境下稳定表达,RIL群体基因分型和表型比较分析表明,是调控垩白的主效QTL位点,并利用RIL群体极端株系将定位在RM1306−RM420标记区间,物理距离约为484.29kb,筛选出12个可能的候选基因。目前,研究团队正在筛选和构建相对剩余杂合体等次级定位群体,以期精细定位目标基因并阐明基因生物学功能,研究将有望为稻米外观品质遗传改良提供参考。
[1] 张昌泉, 赵冬生, 李钱峰, 顾铭洪, 刘巧泉. 稻米品质性状基因的克隆与功能研究进展[J].中国农业科学, 2016, 49: 4267-4283.
Zhang C Q, Zhao D S, Li Q F, Gu M H, Liu Q Q. Progresses in research on cloning and functional analysis of key genes involving in rice grain quality[J]., 2016, 49: 4267-4283. (in Chinese with English abstract)
[2] 程式华. 中国水稻育种百年发展与展望[J].中国稻米, 2021, 27(4):1-6.
Cheng S H. One-hundred years’ development and prospect of rice breeding in China[J]., 2021, 27(4): 1-6. (in Chinese with English abstract)
[3] 周立军, 江玲, 翟虎渠, 万建民. 水稻垩白的研究现状与改良策略[J]. 遗传, 2009, 31(6): 563-572.
Zhou LJ, Jiang L, Zhai H Q, Wang J M. Current status and strategies for improvement of rice grain chalkiness[J]., 2009, 31(6): 563-572. (in Chinese with English abstract)
[4] 邱先进, 袁志华, 何文静, 刘环, 徐建龙, 邢丹英. 水稻垩白性状遗传育种研究进展[J]. 植物遗传资源学报, 2014, 15(5): 992-998.
Qiu X J, Yuan Z H, He W J, Liu H, Xu J L, Xing D Y. Progress in genetic and breeding research on rice chalkiness[J]., 2014, 15(5): 992-998. (in Chinese with English abstract)
[5] Peng B, Wang L Q, Fan C C, Jiang G H, Luo L J, Li Y B, He Y Q. Comparative mapping of chalkiness components in rice using five populations across two environments[J]., 2014, 15: 1.
[6] Gao F Y, Zeng L H, Qiu L, Lu X J, Ren J S, Wu X T, Su X W, Gao Y M, Ren G J. QTL mapping of grain appearance quality traits and grain weight using a recombinant inbred population in rice (L.)[J]., 2016, 15(8): 1693-1702.
[7] Yun P, Zhu Y, Wu B, Gao G J, Sun P, Zhang Q L, He Y Q. Genetic mapping and confirmation of quantitative trait loci for grain chalkiness in rice[J]., 2016, 36: 162.
[8] Ren D Y, Rao Y C, Huang L C, Leng Y J, Hu J, Lu M, Zhang G H, Zhu L, Gao Z, Dong G J, Guo L B, Qian Q,Zeng D L. Fine mapping identifies a new QTL for brown rice rate in rice (L.)[J]., 2016, 9: 4.
[9] Wu B, Yun P, Zhou H, Xia D, Gu Y, Li P B, Yao J L, Zhou Z Q, Chen J X, Liu R J, Cheng S Y, Zhang H, Zheng Y Y, Lou G M, Chen P L, Wan S S, Zhou M S, Li Y H, Gao G J, Zhang Q L, Li X H, Lian X M, He Y Q. Natural variation inregulates redox homeostasis in rice endosperm to affect grain quality[J]., 2022, 36(5): 1912-1932.
[10] Li Y B, Fan C C, Xing Y Z, Yun P, Luo L J, Yan B, Peng B, Xie W B, Wang G W, Li X H, Xiao J H, Xu C G, He Y Q.encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice[J]., 2014, 46(4): 398-404.
[11] 邱先进, 袁志华, 陈凯, 杜斌, 何文静, 杨隆维, 徐建龙, 邢丹英, 吕文恺. 用全基因组关联分析解析籼稻垩白的遗传基础[J] .作物学报, 2015, 41(7): 1007-1016.
Qiu X J, Yuan Z H, Chen K, Du B, He W J, Yang L W, Xu J L, Xing D Y, Lu W K. Genetic dissection of grain chalkiness in indica mini-core germplasm using genome-wide association method[J]., 2015, 41(7): 1007-1016. (in Chinese with English abstract)
[12] Wang E, Wang J J, Zhu X D, Hao W, Wang L Y, Li Q, Zhang L X, He W, Lu B R, Lin H X, Ma H, Zhang G Q, He ZH. Control of rice grain-filling and yield by a gene with a potential signature of domestication[J]., 2008, 40(11): 1370-1374.
[13] Cai Y C, Li S F, Jiao G A, Sheng Z H, Wu Y W, Shao G N, Xie L H, Peng C, Xu J F, Tang S Q, Wei X J, Hu P S.encodes a plastidic pyruvate kinase involved in rice endosperm starch synthesis, compound granule formation and grain filling[J]., 2018, 16(11): 1878-1891.
[14] Zhu A K, Zhang Y X, Zhang Z H, Wang B F, Xue P, Cao Y R, Chen Y Y, Li Z H, Liu Q E, Cheng S H, Cao L Y. Genetic dissection of for a quantitative trait locus for percentage of chalky grain in rice (L.) [J]., 2018, 9: 1173.
[15] Zhou L J, Chen L M, Jiang L, Zhang W W, Liu L L, Liu X, Zhao Z G, Liu S J, Zhang L J, Wang J K, Wan J M. Fine mapping of the grain chalkiness QTLin rice (L.)[J]., 2009, 118(3): 581-590.
[16] Guo T, Liu X L, Wan X Y, Weng J F, Liu S J, Liu X, Chen M J, Li J J, Su N, Wu F Q, Cheng Z J, Guo X P, Lei C L, Wang J L, Jiang L, Wan J M. Identification of a stable quantitative trait locus for percentage grains with white chalkiness in rice ()[J]., 2011, 53(8): 598-607.
[17] Gao Y, Liu C L, Li Y Y, Zhang A P, Dong G J, Xie L H, Zhang B, Ruan B P, Hong K, Xue D W, Zeng D L, Guo L B, Qian Q, Gao Z Y. QTL analysis for chalkiness of rice and fine mapping of a candidate gene for[J]., 2016, 9: 41.
[18] 王小雷, 刘杨, 孙晓棠, 欧阳林娟, 潘锦龙, 彭小松, 陈小荣, 贺晓鹏, 傅军如, 边建民, 胡丽芳, 徐杰, 贺浩华, 朱昌兰. 不同环境下稻米品质性状QTL的检测及稳定性分析[J]. 中国水稻科学, 2020, 34(1): 17-27.
Wang X L, Liu Y, Sun Y, Sun X T, Oyang L J, Pan J L, Peng X S, Chen X R, He X P, Fu J R, Bian J M, Hu L F, Xu J, He H H, Zhu C L. Identification and stability analysis of QTL for grain quality traits under multiple environments in rice[J]., 2020, 34(1): 17-27. (in Chinese with English abstract)
[19] 杨亚春, 倪大虎, 宋丰顺, 李泽福, 易成新, 杨剑波. 不同生态地点下稻米外观品质性状的QTL定位分析[J].中国水稻科学, 2011, 25(1): 43-51.
Yang Y C, Ni D H, Song F S, Li Z F, Yi C X, Yang J B.Identification of QTLs for rice appearance quality traits across different ecological sites[J]., 2011, 25(1): 43-51. (in Chinese with English abstract)
[20] Zhao X Q, Daygon V D, McNally K L, Hamilton R S, Xie F M, Reinke R F, Fitzgerald M A. Identification of stable QTLs causing chalk in rice grains in nine environments[J]., 2016, 129: 141-153.
[21] 彭强, 李佳丽, 张大双, 姜雪, 邓茹月, 吴健强, 朱速松. 不同环境基于高密度遗传图谱的稻米外观品质QTL定位[J]. 作物学报, 2018, 44(8): 1248-1255.
Peng Q, Li J L, Zhang D S, Jiang X, Deng R Y, Wu J Q, Zhu S S. QTL mapping for rice appearance quality traits based on a high-density genetic map in different environments[J]., 2018, 44(8): 1248-1255. (in Chinese with English abstract)
[22] 陈喜娜, 袁泽科, 胡珍珍, 赵全志, 孙红正. 利用QTL-Seq定位粳稻整精米率QTL[J]. 中国水稻科学, 2021, 35(5): 449-454.
Chen X N, Yuan Z K, Hu Z Z, Zhao Q Z, Sun H Z. QTL-seq mapping of head rice rate QTLs in japonica rice[J]., 2021, 35(5): 449-454. (in Chinese with English abstract)
[23] 刘进, 姚晓云, 刘丹, 余丽琴, 李慧, 王棋, 王嘉宇, 黎毛毛. 不同生态环境下水稻穗部性状QTL鉴定[J]. 中国水稻科学, 2019, 33 (2): 124-134.
Liu J, Yao X Y, Liu D, Yu L Q, Li H, Wang Q, Wang J Y, Li M M. Identification of QTL for panicle traits under multiple environments in rice (L.)[J]., 2019, 33(2): 124-134. (in Chinese with English abstract)
[24] Wang J K, Li H H, Zhang L Y. Meng L. QTL IciMapping 4.2[OE/OL]. http://www.isbreeding.net.
[25] McCouch S R. Gene nomenclature system for rice[J]., 2008, 1: 72-84.
[26] Zheng T Q, Xu J L, Li Z K, Zhai H Q, Wan J M. Genomic regions associated with milling quality and grain shape identified in a set of random introgression lines of rice (L.)[J]., 2007, 126: 158-163.
[27] 方雅洁, 朱亚军, 吴志超, 陈凯, 申聪聪, 石英尧, 徐建龙. 全基因组关联定位籼稻种质资源外观和加工品质QTL[J]. 作物学报, 2018, 44(1): 32-42.
Fang Y J, Zhu Y J, Wu Z C, Chen K, Shen C C, Shi Y Y, Xu J L. Genome-wide association study of grain appearance and milling quality in a worldwide collection of indica rice germplasm[J]., 2018, 44(1): 32-42. (in Chinese with English abstract)
[28] 李一博, 赵雷. 水稻品品质性状的遗传改良及其关键科学问题[J].生命科学, 2016, 28(10): 1168-1179.
Li Y B, Zhao L.Genetic improvement and key scientific questions of grain quality traits in rice[J]., 2016, 28(10): 1168-1179. (in Chinese with English abstract)
[29] 吉志军, 尤娟, 王龙俊, 王绍华, 杜永林, 张国发, 王强盛, 丁艳锋. 不同基因型水稻稻米加工品质和外观品质的生态型差异[J]. 南京农业大学学报, 2005, 28(4): 16-20.
Ji Z J, You J, Wang L J, Wang SH, Du Y L, Zhang Q S, Ding Y F. Ecotype differences in milling qualities and appearance qualities of different rice genotypes[J]., 2005, 28(4): 16-20. (in Chinese with English abstract)
[30] 王云霞, 杨连新. 水稻品质对主要气候变化因子的响应[J]. 农业环境科学学报, 2020, 39(4):822-833.
Wang Y X, Yang L X. Response of rice quality to major climate change factors[J]., 2020, 39(4): 822-833. (in Chinese with English abstract)
[31] Wan X Y, Wan J M, Weng J F, Jiang L, Bi J C, Wang C M, Zhai H Q. Stability of QTLs for rice grain dimension and endosperm chalkiness characteristics across eight environments[J]., 2005, 110: 1334-134.
[32] 李进波, 戚华雄. 水稻灌浆期间高温对水稻外观品质的影响[J]. 湖北农业科学, 2019, 58(22): 28-30, 53.
Li J B, Qi H X. Effects on appearance quality of rice under high temperature during grain filling stage[J].,019, 58(22): 28-30, 53. (in Chinese with English abstract)
[33] 徐富贤, 刘茂, 周兴兵, 郭晓艺, 张林, 蒋鹏, 朱永川, 熊洪.长江上游高温伏旱区气象因子对杂交中稻产量与稻米品质的影响[J]. 应用与环境生物学报, 2020, 26(1): 106-116.
Xu F X, Liu M, Zhou X B, Guo X Y, Zhang L, Jiang P, Zhu Y C, Xiong H. Effects of meteorological factors on yield and quality of mid-season hybrid rice in a high temperature and drought area in the upper reaches of the Yangtze River[J]., 2020, 26(1): 106-116. (in Chinese with English abstract)
[34] 王东明, 陶冶, 朱建国, 刘钢, 朱春梧. 稻米外观与加工品质对大气CO2浓度升高的响应[J]. 中国水稻科学, 2019, 33(4): 338-346.
Wang D M, Tao Z, Zhu J G, Liu G, Zhu C W. Responses of rice appearance and processing quality to elevated atmospheric CO2concentration[J]., 2019, 33(4): 338-346. (in Chinese with English abstract)
[35] 吴海兵, 刘道红, 钟鸣, 汪友元.气候因子对稻米品质形成及其影响机制的研究进展[J]. 湖北农业科学, 2019, 58(2): 13-18.
Wu H B, Liu D H, Zhong M, Wang Y Y. Research progress of climate factor on quality formation and influence mechanism in rice[J]., 2019, 58(2): 13-18. (in Chinese with English abstract)
[36] 李承欣, 王敬国, 刘化龙, 孙健, 王江旭, 赵宏伟, 邹德堂. 水、旱条件下稻米品质相关性状的QTL定位及其与环境互作分析[J]. 农业生物技术学报, 2016, 24(10): 1491-1499.
Li C H, Wang J G, Liu H L, Sun J, Wang J X, Zhao H W, Zou D T. QTL mapping and QTL × environment interaction analysis of grain quality-related traits in rice under water-and dry-cultivation conditions[J]., 2016, 24(10): 1491-1499. (in Chinese with English abstract)
[37] 翁建峰, 万向元, 郭涛, 江玲, 翟虎渠, 万建民. 利用CSSL群体研究稻米加工品质相关QTL表达的稳定性[J]. 中国农业科学, 2007, 40(10): 2128-2135.
Weng J F, Wan X Y, Guo T, Jiang L, Zhai H Q, Wan J M. Stability analysis of QTL for milling quality of rice (L.) using CSSL population[J]., 2007, 40(10): 2128-2135. (in Chinese with English abstract)
[38] Zhao X Q, Daygon V D, McNally K L, Hamilton R S, Xie F M, Reinke R F, Fitzgerald M A. Identification of stable QTLs causing chalk in rice grains in nine environments[J]., 2016, 129: 141-153.
[39] 胡霞, 石瑜敏, 贾倩, 徐琴, 王韵, 陈凯, 孙勇, 朱苓华, 徐建龙, 黎志康. 影响水稻穗部性状及籽粒碾磨品质的 QTL 及其环境互作分析[J]. 作物学报, 2011, 37(7): 1175-1185.
Hu X, Shi Y M, Jia Q, Xu Q, Wang Y, Chen K, Sun Y, Zhu L H, Xu J L, Li Z K. Analyses of QTLs for rice panicle and milling quality traits and their interaction with environment[J]., 2011, 37(7): 1175-1185. (in Chinese with English abstract)
[40] 周勇, 高云, 朱松松, 朱金燕, 王军, 裔传灯, 仲维功, 梁国华. 基于染色体单片段代换系的水稻糙米率QTL的定位[J]. 华北农学报, 2013, 28(6): 1-5.
Zhou Y, Gao Y, Zhu S S, Zhu J J, Wang J, Yi C D, Zhong W G, Liang G H. QTL Mapping of brown rice rate using chromosome single segment substituted lines in rice[J]., 2013, 28(6): 1-5. (in Chinese with English abstract)
[41] 王小雷, 刘杨, 孙晓棠, 欧阳林娟, 潘锦龙, 彭小松, 陈小荣, 贺晓鹏, 傅军如, 边建民, 胡丽芳, 徐杰, 贺浩华, 朱昌兰. 不同环境下稻米品质性状QTL的检测及稳定性分析[J]. 中国水稻科学, 2020, 34(1): 17-27.
Wang X L, Liu Y, Sun Y, Sun X T, Oyang L J, Pan J L, Peng X S, Chen X R, He X P, Fu J R, Bian J M, Hu L F, Xu J, He H H, Zhu C L.Identification and stability analysis of QTL for grain quality traits under multiple environments in rice[J]., 2020, 34(1): 17-27. (in Chinese with English abstract)
[42] Song X J, Huang W, Shi M, Zhu M Z, Lin H X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J]., 2007, 39:623-630.
[43] Hu J, Wang Y X, Fang Y X, Zeng L J, Xu J, Yu H P, Shi Z Y, Pan J J, Zhang D, Kang S J, Zhu L, Dong G J, Guo L B, Zeng D L, Zhang G H, Xie L H, Xiong G S, Li J Y, Qian Q. A rare allele ofenhances grain size and grain yield in rice[J]., 2015, 8(10): 1455-1465.
[44] Qiu X J, Chen K, Lü W K, Ou X X, Zhu Y J, Xing D Y, Yang L W, Fan F J, Yang J, Xu J L, Zheng T Q, Li Z K. Examining two sets of introgression lines reveals background independent and stably expressed QTL that improve grain appearance quality in rice [J]., 2017, 130: 951-967.
Identification of QTL for Milling and Appearance Quality Traits in Rice (L.)
YAO Xiaoyun, CHEN Chunlian, XIONG Yunhua, HUANG Yongping, PENG Zhiqing, LIU Jin, YIN Jianhua*
[Rice Research Institute, Jiangxi Academy of Agricultural Sciences/National Engineering Laboratory for Rice (Nanchang)/Nanchang Sub-center, National Rice Improvement Center, Nanchang 330200, China;*Corresponding author, email: jxyjh2011@163.com]
【Objective】Milling and appearance quality(MAQ) are the key factors affecting the production efficiency and commodity value of rice. Exploringthe major QTLs for MAQ related traitshelp further clarify the molecular genetic mechanism of quality traits, which also provides genetic materials for high-quality breeding in rice.【Method】A recombinant inbred line(RIL) population derived from the cross between Longdao 5 (high-quality) and Zhongyouzao 8 (high-yield) was used to identify QTLs for milling and appearance quality traits in multiple environments.【Result】Thirty-sevenQTLs related to processing and appearance quality were detectedon 12 chromosomes. 15, 15, and 21 QTLs were found in the three environments, respectively. Only 10 QTLs were stably expressed in two environments, and two QTLs were repeatedly detected in all the three environments. Processing and appearance quality related QTLs showed significant genetic overlap effects. In addition, QTLs of the milling and appearance quality had obvious genetic overlap effect. There were five major QTL clusters on the chromosomes 2, 6, 7, 10 and 11, which had obvious regulatory effects on milling and appearance quality.【Conclusion】() is a novel pleiotropic QTL clusteronchromosome 7 between RM1306 and RM420, which exerts obvious regulatory effects on percentage of chalky grainsand chalkiness degree. In addition, the epistatic effect plays an important role in the regulation of milling and appearance quality traits, and then the major effect QTL was involved in epistatic interaction effect.
rice; milling quality; appearance quality; QTL mapping;
10.16819/j.1001-7216.2023.220908
2022-09-30;
2022-12-08。
江西现代农业科研协同创新专项(JXXTCXQN202208, JXXTCXQN202205);江西省水稻产业技术体系项目(JXARX-02);江西省青年基金资助项目(20202BABL215002, 20212BAB215030)。
