干旱胁迫对桔梗光合作用和生理特性的影响
2023-09-04信龙飞冀保毅刘红云张艳玲
信龙飞,娄 闯,冀保毅,刘红云,张艳玲
(信阳农林学院,河南 信阳 464000)
近年来,全球气候变暖,季节性降水分布不均,世界各地出现气候异常现象,高温干旱频繁发生[1]。据统计,我国干旱、半干旱地区约占国土面积的1∕2,主要分布在西北和华北等地区[2]。水分已经成为制约植物正常生长、发育成熟和群体繁殖的主要因子之一[3-4]。干旱胁迫会破坏植株的水分平衡,使植物代谢发生紊乱[5]。较多研究表明,干旱胁迫下,植物体内活性氧过量积累,导致膜脂过氧化反应增强,光系统受到损伤,进而影响植物光合进程,造成能量供应不足,代谢产物积累减少,生长发育受阻,产量和品质下降[6-8]。植物为抵御干旱胁迫,演化出一系列防御机制,如抗氧化防御系统和渗透调节等。相关研究结果表明,植物通过提高自身渗透调节物质(脯氨酸、可溶性糖、可溶性蛋白等)含量维持细胞内环境的稳定,保持生理功能正常进行;通过增强体内抗氧化酶活性,清除积累的活性氧,避免氧化损伤[9-10]。总之,干旱胁迫下,植物体内生理代谢过程的改变是植物响应干旱胁迫的重要生物学过程[11-12]。
桔梗(Platycodon grandiflorum)为桔梗科多年生草本植物,是传统道地药材,兼具观赏和药用价值,是一种药、食、赏兼用植物[13]。桔梗在我国大部分地区均有分布,主要种植在安徽、河北、河南、江苏和内蒙古等地,生长发育过程中常受到干旱胁迫,影响其产量和品质[14]。张坚等[15]的研究结果表明,桔梗具有一定程度的干旱适应能力,且适当的干旱可以提高桔梗幼苗的总皂苷含量。然而,目前关于桔梗响应干旱胁迫的研究多是在利用聚乙二醇模拟干旱胁迫的条件下进行的,且主要集中在桔梗苗期阶段[15-16]。探究不同土壤水分条件下植株的光合作用和生理特性变化规律,更有利于解析桔梗响应干旱胁迫的生理调控机制。鉴于此,拟采用盆栽的试验方法,通过设置不同的土壤相对含水量,研究干旱条件下2 年生桔梗的光合和生理特性,以期为深入探讨桔梗耐旱机制及其在干旱地区的生产实践提供理论依据。
1 材料和方法
1.1 试验材料
试验材料为位于河南省信阳市的信阳农林学院农学院试验田中生长状况一致、形态特征相近的2 年生桔梗。将其移栽至花盆内,花盆上口径为21.00 cm、下口径为12.50 cm、高为29.00 cm,盆内装土4.00 kg。每盆移栽1株,共60株。
1.2 试验设计
采用完全随机的试验设计,将全部植株随机分成4 组,每组15 株,设置不同干旱处理,分为对照(CK)、轻度干旱(T1)、中度干旱(T2)、重度干旱(T3),分别对应土壤相对含水量为80%、60%、40%、20%。每天18:00 对盆栽进行称质量并补充水分,以维持各处理的土壤水分含量。干旱处理20 d 后,分别选择不同处理植株中部同一高度的叶片,测定其光合和生理指标,每个处理重复3次。
1.3 测定项目及方法
1.3.1 光合参数和光响应曲线的测定 使用便携式光合仪TARGAS-1 测定桔梗叶片光合参数和光响应曲线。时间为10:00—11:00,从各处理中随机选择3 株生长状况一致的植株,选取其上部相同部位叶片作为测试叶片,叶室面积调节为2 cm2,将外置光源的光照强度逐渐减少,读取并整合数据,得到桔梗叶片的净光合速率(Pn)、胞间CO2浓度(Ci)和气孔导度(Gs)。参照叶子飘[17]的方法利用双曲线修正模型拟合光响应曲线,得到光响应曲线及其参数。
1.3.2 光合色素含量的测定 参照杨慧杰等[18]的方法,采用乙醇浸泡法提取桔梗叶片中的光合色素,用紫外分光光度计测定叶片中的光合色素含量。
1.3.3 生理指标的测定 参照李合生[19]的方法,采用硫代巴比妥酸法测定丙二醛(MDA)含量;采用紫外吸收法测定过氧化氢酶(CAT)活性;采用磺基水杨酸提取-茚三酮显色法测定脯氨酸(Pro)含量;采用蒽酮法测定可溶性糖(SS)、可溶性蛋白(SP)含量。
1.4 数据处理
用Microsoft Excel 2010和GraphPad Prism 8.0进行数据整理并绘制图表,用SPSS 22.0对数据进行方差分析。
2 结果与分析
2.1 干旱胁迫对桔梗气体交换参数的影响
从图1可以看出,随着干旱程度的加重,桔梗植株叶片气体交换参数均呈现下降趋势。与CK 相比,T1—T3 处理Pn 分别显著下降了14.27%、52.89%、77.12%,Gs 分别显著下降了30.76%、58.10%、82.62%,且各处理间差异显著(P<0.05);T1—T3 处理Ci 较CK 分别下降了8.39%、12.35%、31.47%(P<0.05),其中T1与T2处理差异不显著(P>0.05),但均与T3 处理差异显著(P<0.05)。表明干旱胁迫会限制桔梗的光合作用,导致碳同化量减少,进而影响植株生长。

图1 干旱胁迫对桔梗叶片气体交换参数的影响Fig.1 Effect of drought stress on gas exchange parameters of P.grandiflorum leaves
2.2 干旱胁迫对桔梗光响应曲线的影响
如图2 所示,当外界光源给予植株叶片的光照强度小于200µmol∕(m2·s)时,随着光合有效辐射的逐渐增加,桔梗叶片的Pn呈现出线性增加趋势。但是,当外界光源的光照强度继续增加时,各处理桔梗植株叶片Pn的增加幅度逐渐减缓,光照强度高于1 000µmol∕(m2·s)时,叶片Pn 逐渐达到最高值并保持不变,此光照强度即为其光饱和点。当处于光饱和点时,不同干旱胁迫处理的桔梗叶片Pn 不同,CK的最高,其后依次是T1、T2、T3处理。
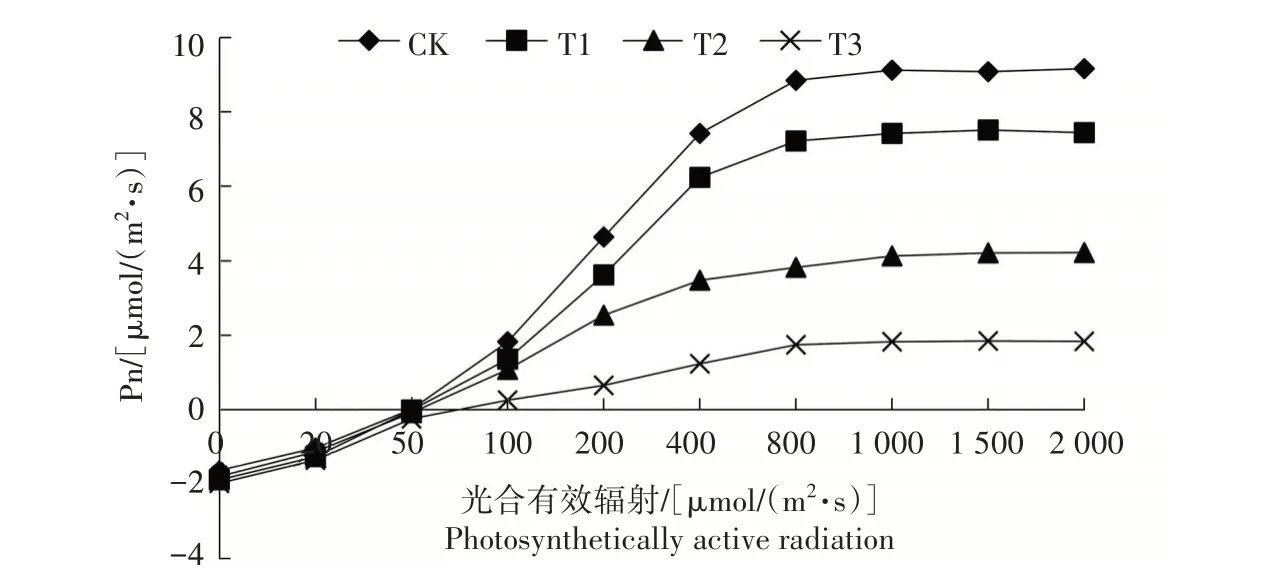
图2 干旱胁迫下桔梗叶片光响应曲线变化特征Fig.2 Change of light response curves of P.grandiflorum leaves under drought stress
2.3 干旱胁迫对桔梗光响应曲线参数的影响
对光响应曲线进行双曲线修正模型拟合分析,其决定系数(R2)均在0.95 以上,说明模型拟合度较高。从表1可以发现,随着干旱程度的加深,桔梗的光补偿点(LCP)呈现出逐渐上升的趋势,T3 处理的LCP 最高,为74.45 µmol∕(m2·s),T1—T3 处理LCP分别比CK 上升8.43%、31.07%、67.76%,其中T2、T3处理与CK 差异显著(P<0.05)。最大净光合速率(Amax)、光饱和点(LSP)、表观量子效率(AQY)和暗呼吸速率(Rd)的变化则相反,均呈现出逐渐降低的趋势。与CK 相比,T1—T3 处理Amax 分别下降18.66%、56.18%、81.24%,AQY 分别下降22.20%、46.81%、72.04%,Rd 分别下降15.61%、32.49%、59.92%,各处理间差异显著(P<0.05)。T1 处理LSP与CK 差异不显著,T2、T3 处理LSP 分别较CK 显著下降9.53%、15.52%(P<0.05)。表明干旱胁迫下,桔梗植株对光的利用能力逐渐变弱,最大光合速率降低,自身制造有机物的能力下降。

表1 干旱胁迫下桔梗光响应曲线参数Tab.1 Light response curve parameters of P.grandiflorum under drought stress
2.4 干旱胁迫对桔梗光合色素含量的影响
如图3所示,随着土壤相对含水量的减少,桔梗叶片光合色素含量先升高后降低。T1 处理桔梗叶片中光合色素含量最高,其中叶绿素a、叶绿素b、类胡萝卜素含量分别为17.13、7.76、5.68 mg∕g,较CK分别增加了18.71%、28.48%、12.25%。随着土壤干旱程度的进一步加重,植株叶片中光合色素含量逐渐下降,T2 处理的叶绿素b 含量较CK 显著增加15.89%(P<0.05),而叶绿素a、类胡萝卜素、总叶绿素含量与CK 差异不显著(P>0.05)。T3 处理光合色素含量下降幅度最大,达到最低点,其叶绿素a、叶绿素b、总叶绿素、类胡萝卜素含量分别比CK 下降了32.43%、17.72%、28.09%、15.81%,且差异显著(P<0.05)。表明轻度和中度干旱可引起桔梗叶片光合色素含量升高,而重度干旱胁迫则会导致植株叶片光合色素尤其是叶绿素分解速率加快,进而影响植株的光合作用。

图3 干旱胁迫对桔梗叶片光合色素含量的影响Fig.3 Effect of drought stress on photosynthetic pigment content of P.grandiflorum leaves
2.5 干旱胁迫对桔梗MDA含量的影响
在逆境条件下,植物细胞膜会受到一定程度的损伤,MDA 的含量能反映其损伤的严重程度。图4显示,CK 桔梗叶片MDA 含量较低,干旱胁迫后,MDA 含量逐渐增加。T1、T2 处理的MDA 含量分别比CK 显著增加25.66%、39.14%(P<0.05),但T1 与T2 处理间差异不显著(P>0.05)。T3 处理MDA 含量最高,为12.03µmol∕g,比CK 增加97.86%,且与其他处理差异显著(P<0.05)。表明随着干旱程度增加,桔梗细胞膜脂过氧化反应明显加强,细胞膜受损伤程度增大,尤其是重度干旱胁迫下,植株细胞膜脂过氧化损伤最为严重。
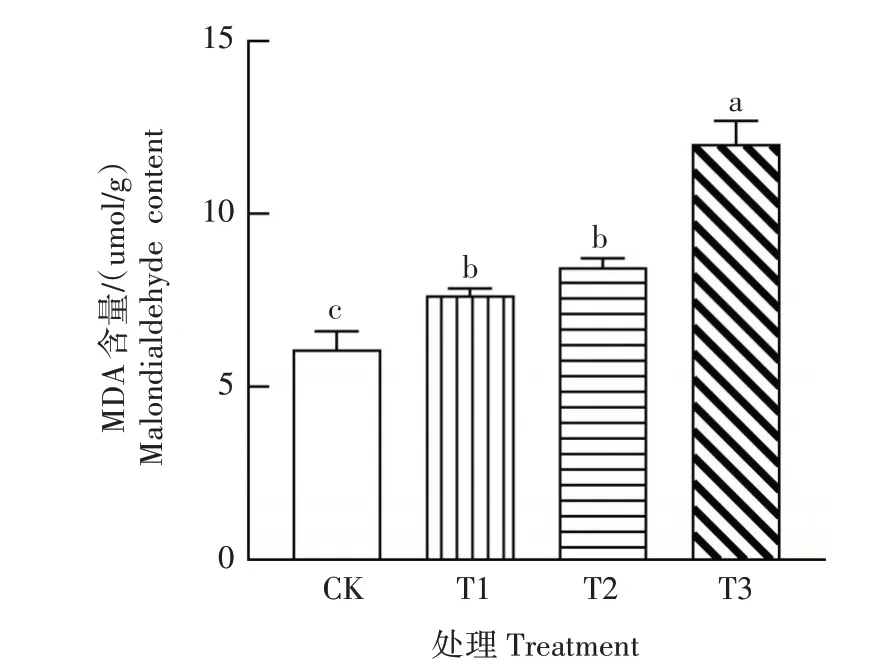
图4 干旱胁迫对桔梗叶片MDA含量的影响Fig.4 Effect of drought stress on MDA content in P.grandiflorum leaves
2.6 干旱胁迫对桔梗CAT活性的影响
CAT 作为一类重要的抗氧化酶,其活性可以反映植物的抗性。从图5 可以看出,随着土壤干旱程度的增加,桔梗叶片CAT 活性先升后降,T2 处理的CAT活性达到最大值。T1、T2处理CAT活性分别比CK 增加11.82%、21.01%,差异显著(P<0.05),而T3处理CAT活性较T1、T2处理显著降低(P<0.05)。这表明,轻度和中度干旱条件下,桔梗植株通过提高CAT 活性减轻氧化损伤,但在重度干旱下,CAT 活性的增加幅度较小,对氧化损伤的缓解作用下降,导致桔梗植株膜脂过氧化程度加重。
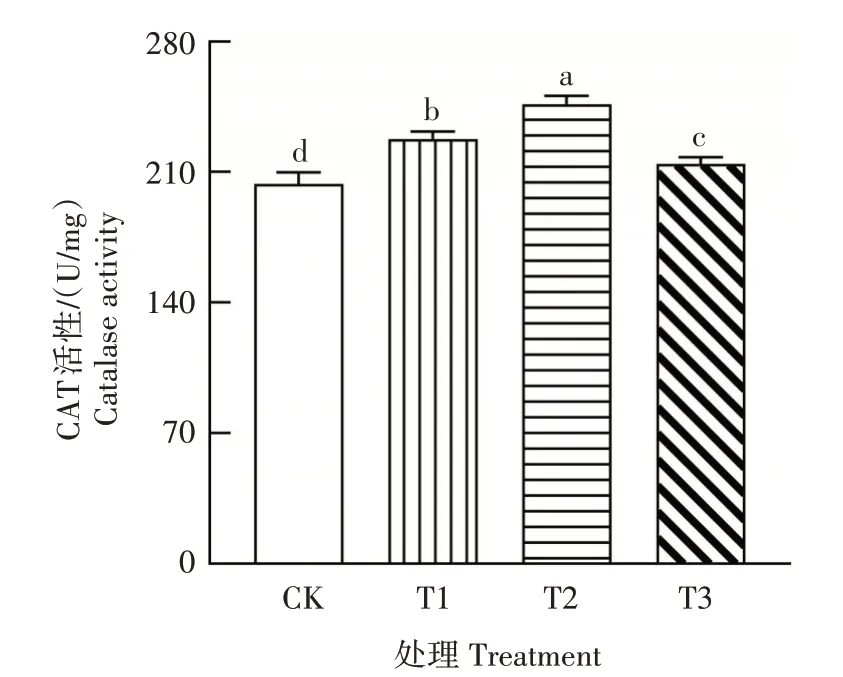
图5 干旱胁迫对桔梗叶片CAT活性的影响Fig.5 Effect of drought stress on CAT activity in P.grandiflorum leaves
2.7 干旱胁迫对桔梗渗透调节物质的影响
如图6所示,当土壤相对含水量逐渐减少时,植株体内的Pro、SS、SP 含量均逐渐增加。与CK 相比,T1、T2、T3 处理桔梗叶片Pro 含量分别增加了19.50%、66.31%、83.84%,且各处理间差异显著(P<0.05)。T1处理的SS含量与CK 相比有所增加,且差异显著(P<0.05);T2、T3 处理SS 含量较CK 分别增加了46.51%、75.23%,且差异显著(P<0.05)。T1、T2处理SP 含量分别比CK 显著增加了59.51%、63.97%(P<0.05),但T1、T2 处理间无显著差异(P>0.05)。T3 处理SP 含量最高,较CK 显著增加了93.73%,且与其他处理差异均显著(P<0.05)。表明干旱胁迫下,桔梗可通过积累渗透调节物质,维持细胞的相对稳定,提高植株的耐旱性。
3 结论与讨论
植物有机体在遭受干旱等逆境胁迫时,通过调控内部生理生化等一系列变化活动来适应环境[20-21]。植物叶片作为光合作用最重要的组织器官,一方面能够调控叶片气孔保卫细胞的水势变化,诱导保卫细胞吸水或失水,调节叶片气孔开闭;另一方面也可通过内外环境气体交换来进行光合作用,保证能量供应[22]。研究表明,在干旱胁迫下,植物通过气孔的张开程度,调控CO2、O2和水蒸气等气体的进出,会对植株的光合作用产生影响[23]。本研究结果表明,随着干旱程度的加深,桔梗叶片Gs、Ci 和Pn 均呈现下降趋势。其中,Gs 下降可降低干旱环境下植株的蒸腾速率,保持叶片含水量的相对稳定;Ci 下降可使植株叶片无法进行高强度的光合作用,从而缓解干旱胁迫对植株光合系统的损伤。高冠龙等[24]的研究结果表明,当植物遭受水分胁迫时,植株叶片的Pn、Gs 和Ci 同时下降,说明Pn 降低主要是受气孔限制;Pn 和Gs下降,而Ci不变或升高时,则说明Pn 降低主要是受非气孔限制。本研究中,干旱胁迫处理后,桔梗叶片的Gs、Pn 和Ci 较CK均显著下降,表明在不同干旱条件下,导致桔梗植株叶片净光合速率降低的主要因素为气孔限制。
通过双曲线修正模型拟合光响应曲线,获得植株叶片的AQY、Amax、Rd、LSP、LCP 等一系列光响应曲线参数,可解析不同处理叶片光合作用的响应机制[17]。其中,AQY 表示植株叶片对弱光的利用能力,其值越大,表明植物体内吸收与转换光能的色素蛋白复合体利用弱光的能力越强[25]。本研究中,正常条件下,桔梗叶片AQY 处于较高水平,对光能利用率较高,干旱处理后,植株叶片AQY逐渐降低,即植株的光能利用率受到明显的抑制,这与李思忠等[25]的研究结果一致。LCP 和LSP 的高低反映植物对弱光和强光适应能力的强弱,当植株叶片LCP 较低、LSP 较高时,意味着植株对光环境的适应性较强;当植株叶片LCP 较高、LSP 较低时,植株则难以利用外界的光环境。本研究结果表明,轻度干旱胁迫下,桔梗的LCP 和LSP 没有显著变化。随着干旱程度的加深,桔梗叶片的LCP逐渐升高,而LSP逐渐降低,在重度干旱胁迫下,桔梗叶片LCP 最高、LSP最低,此时植株光能利用率和对环境适应能力最弱。同时,桔梗叶片Amax 和Rd 也随着干旱程度的增加而降低,表明干旱胁迫会导致桔梗光合速率下降,进而抑制植株有机物的积累,影响其正常生长发育,植株通过降低呼吸作用抵御干旱胁迫,这与黄明等[26]的研究结果一致。
叶绿素为植物进行光合作用的主要色素,色素含量与植株光合能力呈正相关[27]。程宇飞等[28]在关于新西兰麻的研究中发现,植株叶片中叶绿素a、叶绿素b和总叶绿素含量随土壤干旱化程度的加深均呈现出先上升后下降的趋势。本研究结果与前人相似,在轻度和中度干旱条件下,桔梗叶片光合色素含量升高,尤其是轻度干旱胁迫处理的叶绿素a、叶绿素b 和总叶绿素以及类胡萝卜素含量与CK 差异显著。当干旱程度继续增加时,光合色素含量明显下降。叶绿素含量的增加可能是桔梗对干旱胁迫的应激反应,轻度和中度干旱胁迫能刺激桔梗光合色素的合成,以维持较高的光合生产力,抵抗胁迫带来的影响,这说明桔梗具有一定抵御干旱胁迫的能力。但随着干旱危害程度的进一步加深,光合色素的合成受阻,分解速率加快,导致光合色素含量下降,光合能力减弱,说明桔梗对重度干旱胁迫的耐受性较差。逆境条件下,植物体内活性氧产生速率加快,膜脂过氧化程度加重,氧化损伤标志物MDA 含量显著增加,对植物细胞膜造成伤害[29-30]。本研究中,随着土壤干旱程度的增加,桔梗叶片MDA 含量逐渐升高,膜脂过氧化明显加强,植株细胞膜损伤程度增大,尤其在重度干旱胁迫下,植株体内MDA 含量达到最高值,细胞受损程度严重,这与裴斌等[31]在沙棘叶片上的研究结果相似。本研究中,干旱胁迫下桔梗光合速率和光合色素含量下降,原因是膜脂过氧化程度加重,造成叶绿素合成和代谢失衡,蛋白质变性,导致叶绿素减少和光系统结构受到氧化损伤。
植物为适应外界环境的变化,进化形成了复杂的防御机制,能够有效清除活性氧积累,减少膜脂过氧化对植物机体造成的损伤[32]。较多研究表明,CAT 作为植物体内重要的保护酶,在植物抗逆性方面发挥着重要作用,可有效清除植物体内产生的过量活性氧,减轻细胞膜脂过氧化损伤,保护细胞膜的完整性[33]。本研究中,随着干旱程度的增加,桔梗叶片中CAT 活性先升后降,中度干旱胁迫时达到峰值,且CAT 在不同干旱胁迫下均保持较高的活性。但重度干旱胁迫下,CAT 活性增加幅度较小,不能有效缓解干旱胁迫对桔梗植株造成的氧化损伤,导致膜脂过氧化程度加剧。植物还可通过细胞内渗透调节物质的积累降低渗透势,维持细胞膨压的相对稳定,提高植株细胞对干旱胁迫的渗透调节能力[34]。Pro、SS 和SP 等是植物主要的渗透调节物质,它们作为一种亲水性有机溶剂,能够稳定蛋白质结构,保护植株体内的大分子物质,与植物耐旱性呈正相关[35]。研究表明,在干旱胁迫下,植物体内的渗透调节物质会有显著积累[36]。本研究结果也证实,随着土壤干旱程度的加深,桔梗叶片Pro、SS、SP 等含量迅速增加,说明渗透调节物质在桔梗耐旱性方面扮演着重要角色,这与郑钢等[37]在黄薇上的研究结果一致。
综上所述,干旱胁迫下,桔梗通过调节气孔的开闭,降低蒸腾速率,减少水分蒸发,以适应干旱胁迫,但同时也会引起叶片Ci 降低,导致Pn 下降。轻度干旱胁迫下,桔梗光合色素含量增加,LCP 和LSP没有显著变化。随着干旱胁迫的加重,桔梗的LSP、AQY 和Rd 逐渐降低,LCP 升高,光合色素含量下降,对光能的利用能力减弱,同时植株通过提高抗氧化酶活性和增加渗透调节物质的积累来维持细胞渗透势的相对稳定,以提高桔梗的抗旱性。但长时间的重度干旱会对桔梗植株的生长和发育造成不可逆损伤。因此,桔梗种植应注意补充土壤水分,避免土壤相对含水量长时间处于40%以下。
