菜用大豆有机酸的全基因组关联分析
2023-09-02许文静张红梅刘晓庆朱月林陈华涛
刘 慧,许文静,杨 硕,张 威,张红梅,刘晓庆,朱月林,陈华涛
(1.南京农业大学 园艺学院,江苏 南京 210095;2.江苏省农业科学院 经济作物研究所,江苏 南京 210014)
菜用大豆(Glycinemax(L.)Merr.)又称毛豆、鲜食大豆,是指在R6(鼓粒盛期)—R7(初熟期)生育期间采青食用的大豆专用型品种[1]。我国作为世界上最大的菜用大豆出口国,虽然菜用大豆产业已得到快速的发展,但有关菜用大豆食味品质的研究还有待进一步加强,这制约了优质菜用大豆产业健康发展,因此,食味品质的相关研究已成为菜用大豆改良育种方向之一。鲜食风味是菜用大豆食味品质的关键因素,其形成与有机酸有着密切的关联。有机酸作为多种氨基酸生物合成的底物,有利于籽粒中氨基酸的积累从而提高菜用大豆的鲜味[2]。Sowalsky等[3]的研究结果表明,酒石酸、苹果酸和柠檬酸与涩味之间存在一定的联系,并且产生的酸味强度具有差异性。因此,不同种类的有机酸有其独特的风味特征。柠檬酸产生酸感快且酸度高于苹果酸,但后味时间短;酒石酸略有涩感,酸味爽口;苹果酸酸味爽口,微有涩苦,呈味时间长,食用适量可给人清爽之感[4],其他有机酸如奎宁酸则主要产生苦味。有机酸不仅能影响菜用大豆的苦味、涩味、酸味,还能通过掩盖糖的味道来影响甜味,Green等[5]的研究发现,柠檬酸可以掩盖味觉细胞对于果糖和蔗糖的感知,而毛岳忠[6]研究发现,苹果酸能增强味觉细胞对于蔗糖的感知。菜用大豆籽粒中有机酸的种类、含量及其构成比例是菜用大豆鲜食风味形成的重要基础,因此,检测菜用大豆的有机酸成分分布,研究有机酸合成机制对于菜用大豆的品质改良具有重要的实际意义。
数量性状的研究对于农作物的遗传改良具有重要的实际意义,在不同的植物中已报道了许多与有机酸含量相关的QTLs。Liebhard等[7]研究表明,苹果果实中可滴定酸含量的变化几乎完全可以由8,16号连锁群上的2个主效QTL位点决定,分别能够解释后代中酸度表型变异的46%,42%,其中位于第16连锁群上的主效基因位置与Maliepaard等[8]报道的结果相一致。菜用大豆中的酸含量属于多个基因协同控制的数量性状,但截至目前,有关菜用大豆酸含量调控机理的研究相对滞后,与菜用大豆有机酸含量相关的SNPs及候选基因有待发掘。与传统的连锁定位方法相比,全基因组关联分析法(Genome wide association study,GWAS)节省了构建群体的时间,能够对不同种质进行遗传结构分析,提高QTL定位的精准性,已经成为快速有效的发掘植物数量性状基因的重要手段[9-10]。本研究以含有264份菜用大豆种质资源的自然群体为试验材料,分别在2020,2021年通过高效液相色谱法(High performance liquid chromatography,HPLC)测定酒石酸、苹果酸和柠檬酸含量,结合GWAS鉴定与有机酸显著关联的SNP位点并进一步挖掘相应的候选基因,旨在初步解析菜用大豆有机酸含量的遗传学基础,为调控菜用大豆有机酸含量的分子机制研究提供理论基础。
1 材料和方法
1.1 试验材料
264份菜用大豆种质资源,包括212份栽培种及52份地方种,均由江苏省农业科学院经济作物研究所大豆课题组提供。分别于2020年6月和2021年6月种植于江苏省农业科学院南京市六合基地试验田(32°28′12″N,118°37′48″E)。田间试验采用随机区组试验设计,3次重复。每份材料重复种植3垄,每垄播种10穴,每穴2株苗,垄长为1.5 m,垄间距40 cm,穴距15 cm。田间管理按照当地常规方法进行。
1.2 仪器与试剂
植物组织研磨仪;高效液相色谱仪Agilent1100;Aglient ZORBAX SB-Aq色谱柱;超声波振荡仪;可见分光光度(UV-2600,日本岛津);超纯水机;ML204型万分之一天平;酸度计、离心机、水浴锅、烘箱及其他实验室常用设备。
甲醇(色谱纯);磷酸(质量分数≥85.0%);苹果酸、酒石酸、柠檬酸(色谱级纯分析标准品),以上试剂均购自上海阿拉丁生物科技有限公司。
1.3 菜用大豆有机酸检测
1.3.1 色谱条件 色谱柱:Aglient ZORBAX SB-Aq C18柱(粒径5 μm,柱长250 mm×4.5 mm);流动相:甲醇-0.1%磷酸水溶液(pH=2.30),梯度洗脱;流速0.8 mL/min;检测波长212 nm;色谱柱温度30 ℃;进样量体积10 μL。流动相洗脱条件见表1。
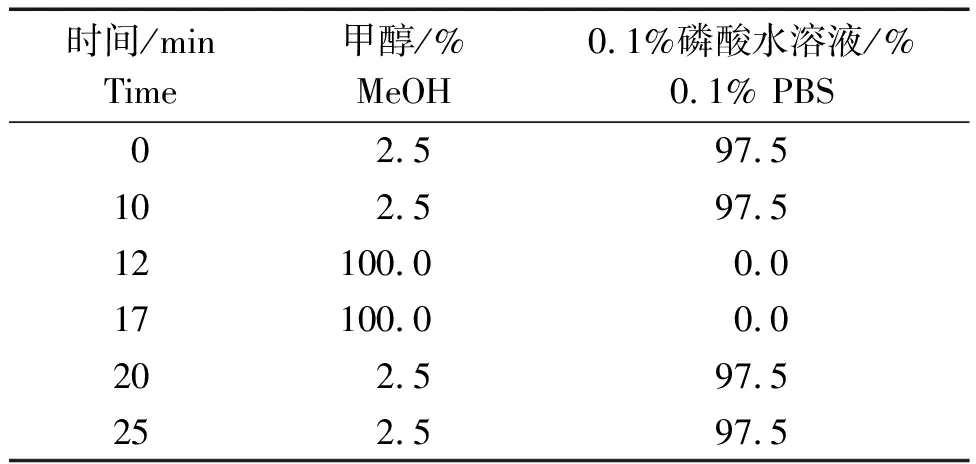
表1 流动相洗脱条件
1.3.2 标准液的配制 精密称取酒石酸、苹果酸各50 mg,柠檬酸10 mg(精确至0.1 mg),用流动相溶解并定容至10 mL棕色容量瓶中。配制成酒石酸、苹果酸、柠檬酸标准品储备液,质量浓度分别为5 000,5 000,1 000 mg/L。
1.3.3 绘制标准曲线 以1.3.2中的标准品储备液为母液,共配制5种不同浓度梯度混标溶液。其中苹果酸和酒石酸浓度分别为5,50,200,1 000,5 000 mg/L,柠檬酸浓度分别为1,10,40,200,1 000 mg/L。以上溶液均经0.45 μm微孔滤膜过滤后,按照1.3.1的色谱检测条件,利用高效液相色谱仪分析后,以浓度x(mg/L)对色谱峰峰面积y绘制标准曲线,并计算回归方程。
1.3.4 有机酸提取方法 根据盖钧镒等[11]的报道,菜用大豆最适宜的采收期为R6期,因此,本试验选择R6期(鼓粒盛期)即鲜荚翠绿,籽粒饱满时进行取样,设置3次生物学重复。将收取的鲜荚放入烘箱先105 ℃杀青30 min,然后以80 ℃烘干至恒质量,取出干籽粒放入植物组织研磨仪中(40 Hz,2 min)研磨至粉末备用。菜用大豆干籽粒粉末过0.45 mm筛,称取粉末1 g(精确至0.000 1 g),置于50 mL量瓶中,加入40 ℃蒸馏水约40 mL,40 ℃下超声提30 min,冷却至室温,定容,混匀。在4 000 r/min下离心20 min,取上清液1 mL,经0.45 μm滤膜过滤,超声脱气后,供分析用。
1.3.5 HPLC分析菜用大豆有机酸 按照1.3.1的色谱检测条件,从同一菜用大豆样品中平行取3份进行重复性试验,根据标品绘制的标准曲线分析计算菜用大豆样品中各有机酸组分的质量浓度,取3次进样测得的平均值作为菜用大豆有机酸组分最终的检测结果。
1.4 数据处理
各有机酸含量数据采用Excel 2019计算平均值、标准差及变异系数,应用SPSS(IBM SPSS Statistics 26.0,SPPS,USA)软件分别对酒石酸、苹果酸和柠檬酸含量进行相关性分析。
1.5 全基因组关联分析
本研究中,用于全基因组关联分析的自然群体SNP标记来自前期的重测序工作,共2 597 425个,该群体的连锁不平衡衰减区间为120 kb,具体群体结构分析见江苏省农业科学院豆类作物研究室的研究报道[12],采用基于群体结构(Q)+亲缘关系(K)的混合线性模型(Mixed linear model,MLM)进行全基因组关联分析以控制假阳性关联位点。全基因组关联分析采用基于R软件的GAPIT算法包进行计算[13],设定-log10(P)=5.0作为显著阈值,当关联到的SNP阈值≥5.0时,则被认为是显著关联位点。
1.6 候选基因分析
全基因组关联分析得到2个不同的环境下(2020,2021年)检测到的显著SNP位点后,挑选出其上下游120 kb区间范围内所有的基因。参照在线数据库Phytozome 13网站(https://phytozome.jgi.doe.gov/pz/portal.html)中的大豆基因组注释信息确定目标性状的候选基因。
2 结果与分析
2.1 HPLC色谱分析
在1.3.1的色谱检测条件下,采用HPLC法分离并检测有机酸标准品和菜用大豆籽粒样品中的酒石酸、苹果酸和柠檬酸,可以清晰地将3种有机酸分离出来,且峰型良好,洗脱顺序分别为酒石酸、苹果酸、柠檬酸,平均保留时间分别为4.316,5.410,9.868 min(图1)。
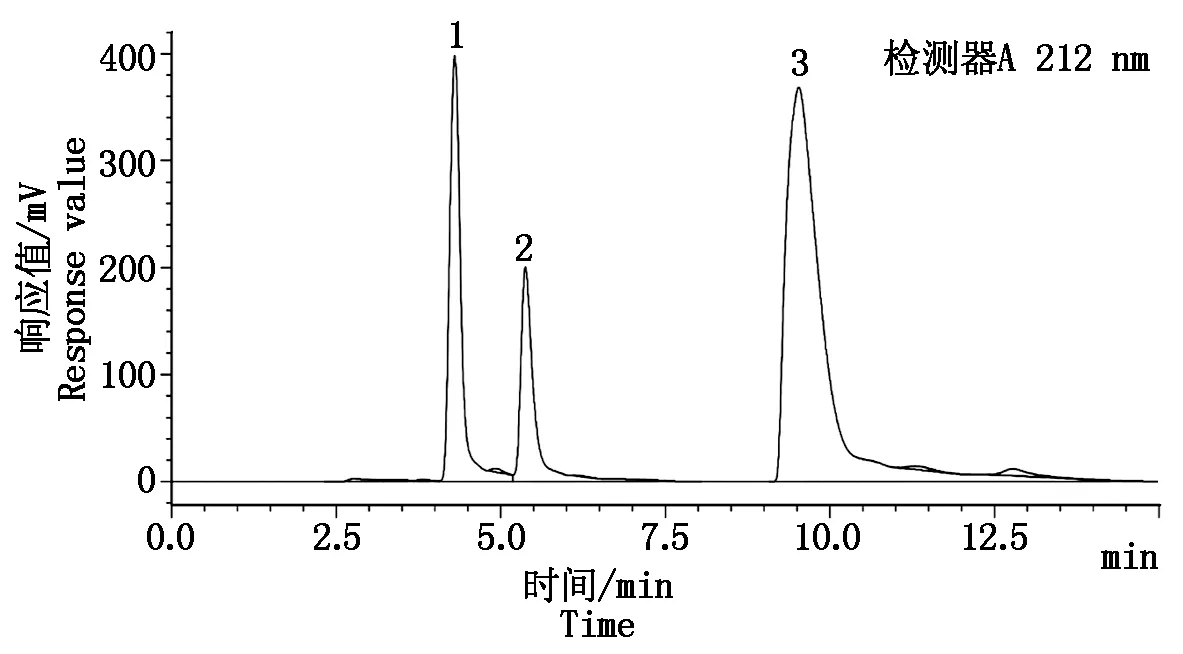
1.酒石酸;2.苹果酸;3.柠檬酸。
2.2 菜用大豆3种有机酸组分线性回归方程
分别将不同浓度梯度的3种有机酸的混合标准溶液经高效液相色谱仪分析后,以质量浓度为横坐标(x),峰面积为纵坐标(y)进行线性回归分析。计算得到酒石酸、苹果酸和柠檬酸标准品的线性回归方程(表2)。3种有机酸标准品的峰面积和质量浓度的相关系数为0.997 9~0.999 1,在设定的浓度范围内呈现良好的线性关系,根据信噪比确定的定量限(RSN=10)为2.5~10 μg/mL。

表2 菜用大豆3种有机酸组分线性回归方程
2.3 表型分析
菜用大豆3种有机酸含量的描述性统计分析(表3)显示,2020,2021年供试群体的酒石酸含量分别为1.66~9.19 mg/g,0.48~13.52 mg/g,平均含量分别为4.13,4.16 mg/g;苹果酸含量分别为1.47~36.46 mg/g,1.89~25.19 mg/g,平均含量分别为7.26,8.99 mg/g;柠檬酸含量分别为2.56~12.05 mg/g,1.54~18.31 mg/g,平均含量分别为7.12,10.88 mg/g。上述结果均表明,供试群体有机酸含量差异较大,种质资源遗传变异丰富,为特异种质的筛选提供了有利条件。

表3 菜用大豆3种有机酸含量的描述性统计分析
2020年菜用大豆酒石酸、苹果酸和柠檬酸变异系数分别为31.25%,67.29%,19.22%,2021年3种有机酸变异系数分别为52.07%,47.07%,26.35%,且3种有机酸组分中柠檬酸变异系数最低。遗传变异系数能够体现各性状的变异丰富度,一般来说,当变异系数大于10%时,可以认为种质间差异较大,且变异系数越大,表明该性状的变异潜力越丰富。2 a中3种有机酸的变异系数均大于10%,反映出供试群体不同品种间有机酸各组分存在丰富的变异,这些遗传变异的存在可为菜用大豆食味品质的改良提供广泛的遗传学基础。
2.4 相关性分析
本研究对264份材料酒石酸、苹果酸、柠檬酸含量在2020,2021年2 a间进行相关性分析,结果表明,264份材料柠檬酸含量在2020,2021年,2 a间呈显著正相关,相关系数为0.156*(表4)。同时,基于供试群体3种有机酸2 a表型数据的平均值,对不同有机酸组分含量进行相关性分析。结果发现,酒石酸含量与苹果酸、柠檬酸含量间均存在显著正相关,相关系数分别为0.695*,0.739*,苹果酸含量与柠檬酸含量也有着显著的正相关,相关系数为0.790*(表5)。

表4 264份材料酒石酸、苹果酸、柠檬酸含量在2020,2021年2 a的相关系数
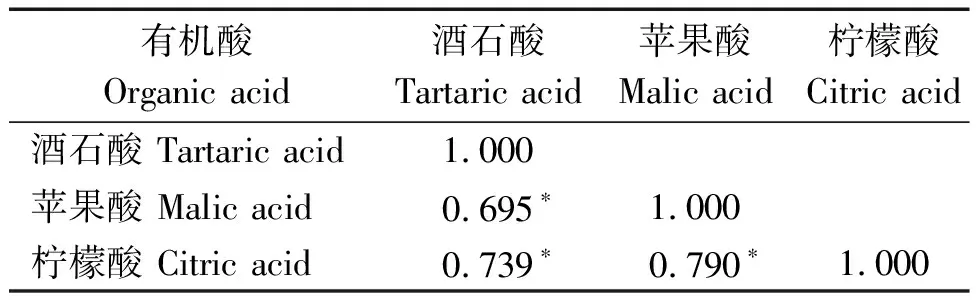
表5 菜用大豆3种有机酸组分相关性分析
2.5 菜用大豆3种有机酸组分含量特异种质筛选
基于供试群体3种有机酸含量,分别对每种有机酸筛选出2份高含量的特异种质。将264份菜用大豆种质资源按照酒石酸含量由高到低排序,并将位于前15%的品种划分为高酒石酸含量组(Tartaric acid high content group,T1),在2020,2021年的检测中均属于T1组的种质为阜04-35和桂春豆112(表6)。同理,将供试群体按照苹果酸含量由高到低排序,并将前15%的品种划分为高苹果酸含量组(Malic acid high content group,M1),2 a中均属于M1组的种质包括华春9号和邯12-204(表7);将供试群体按照柠檬酸含量由高到低排序,前15%的品种被划分为高柠檬酸含量组(Citric acid high content group,C1),2 a中均属于C1组的种质有灌豆3号和新沂大紫花(表8)。以上筛选到的种质,进一步丰富了我国菜用大豆有机酸含量特异种质资源。
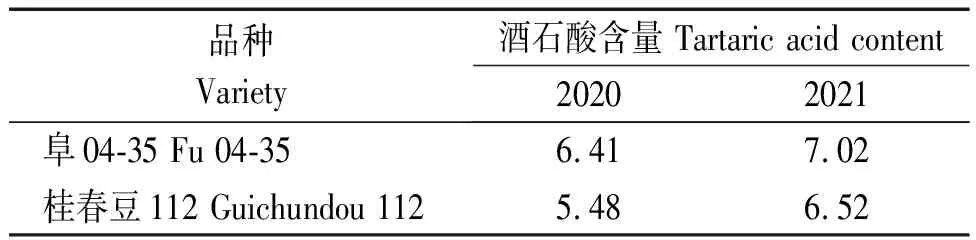
表6 2 a的检测中均筛选到的高酒石酸含量的特异种质

表7 2 a的检测中均筛选到的高苹果酸含量的特异种质
2.6 菜用大豆酒石酸、苹果酸和柠檬酸含量GWAS分析
为了深入研究菜用大豆有机酸含量的遗传机制,利用MLM模型对2020,2021年的酒石酸、苹果酸和柠檬酸含量表型值进行GWAS分析(图2,3),设定显著阈值为-log10(P)= 5.0,如表9所示,2020年检测到与酒石酸显著关联的SNP位点31个,每个SNP解释了7.99%~11.23%的表型变异;2021年检测到与酒石酸显著关联的SNP位点23个,每个SNP解释了7.00%~10.62%的表型变异。2020年检测到与苹果酸显著关联的SNP位点169个,每个SNP解释了7.39%~24.66%的表型变异;2021年检测到与苹果酸显著关联的SNP位点20个,每个SNP解释了7.30%~9.05%的表型变异。在2020年并没有检测到与柠檬酸显著关联的位点,2021年检测到与柠檬酸显著关联的SNP位点43个,分别位于2,6,13号染色体上,单个SNP的表型变异解释率为8.06%~10.96%。上述结果表明,菜用大豆籽粒中有机酸含量是由多基因控制的。
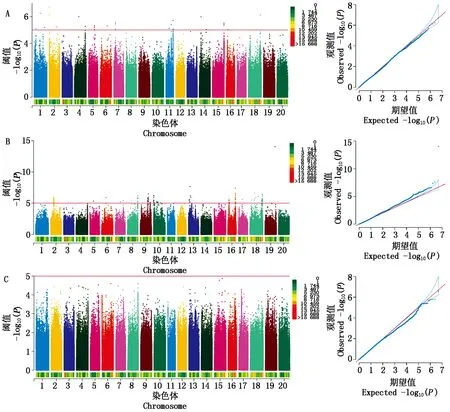
A、B和C分别是酒石酸、苹果酸和柠檬酸的GWAS结果。图3同。
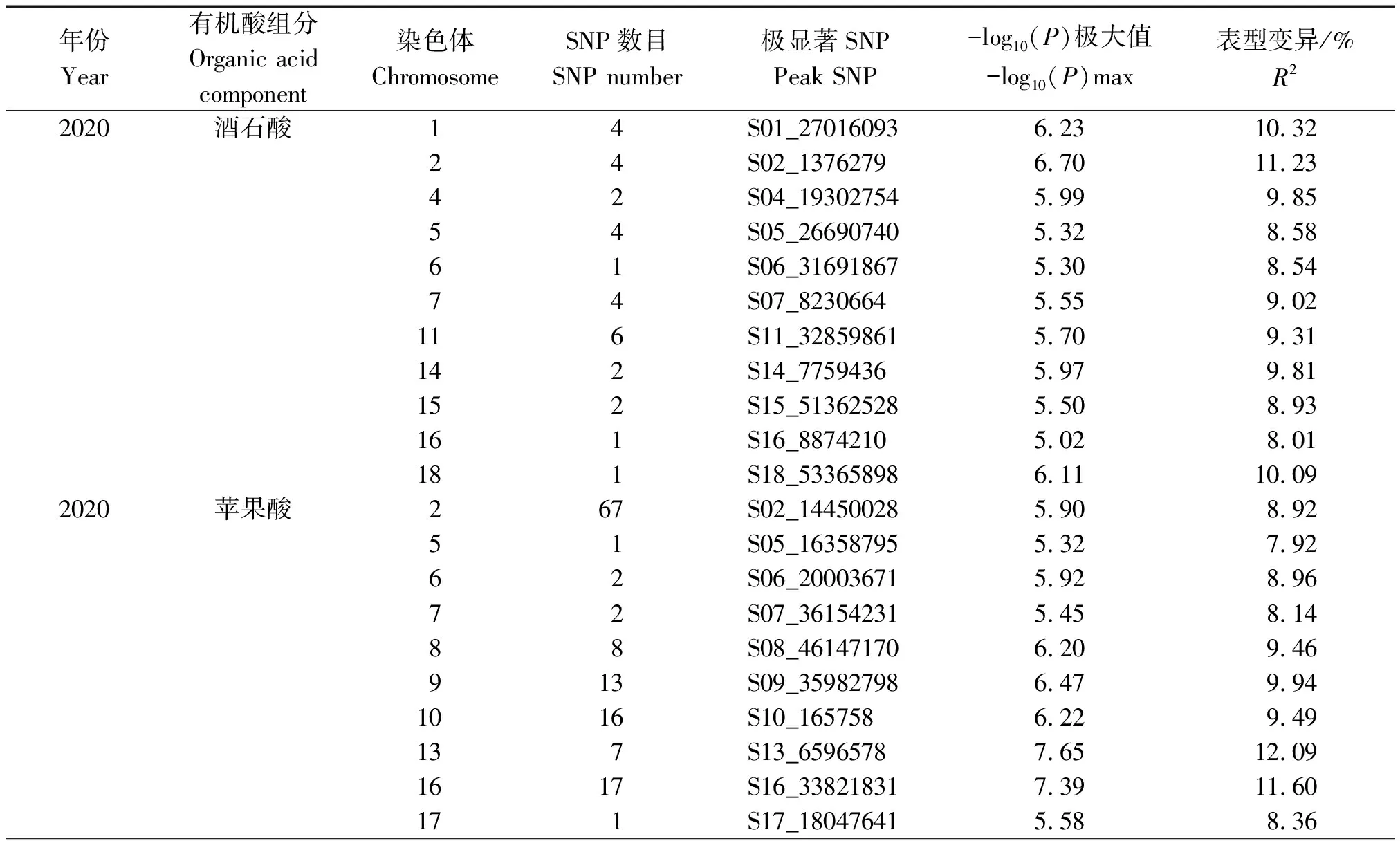
表9 3种有机酸组分显著相关SNPs分布
2.7 菜用大豆3种有机酸组分相关候选基因挖掘
为了研究显著关联的SNP位点等位变异的表型效应,分别对2020,2021年中酒石酸、苹果酸和柠檬酸检测到的阈值最高的显著SNP位点进行单倍型分析。发现SNP S02_1376279碱基变化为A至T,携有SNP S02_1376279-A的菜用大豆种质的酒石酸平均含量为4.01 mg/g,显著低于基因型S02_1376279-T的菜用大豆种质的6.40 mg/g(图4-A)。S19_44610972的等位变异为C/T,携有SNP S19_44610972-C的菜用大豆种质的苹果酸平均含量为6.52 mg/g,显著低于基因型S19_44610972-T的菜用大豆种质的19.52 mg/g(图4-B)。S17_41027045的等位变异为T/A,携有S17_41027045-T的菜用大豆种质的酒石酸平均含量要显著低于携有S17_41027045-A的菜用大豆种质的酒石酸平均含量(图4-C)。S13_25565436和S06_6589288的等位变异均为G/A,携有S13_25565436-G的菜用大豆种质的苹果酸平均含量要显著低于携有S13_25565436-A的菜用大豆种质,而携有S06_6589288-G的菜用大豆种质的柠檬酸平均含量显著高于携有S06_6589288-A的菜用大豆种质(图4-D、E)。

同一图中不同小写字母表示在0.05水平下差异显著。
在检测到的与3种有机酸显著关联(-log10(P)≥ 5.0)的SNP位点上下游120 kb范围内进行候选基因的筛选及功能预测。参照大豆参考基因组的基因功能注释信息,鉴定到8个与有机酸含量显著相关的候选基因(表10),这些基因可能参与菜用大豆籽粒中有机酸的调控和积累,作为重要候选基因进一步分析。共挖掘到3个与酒石酸含量显著相关的候选基因,其中,定位于Chr.02上的基因Glyma.02G015700和Glyma.02G015800分别编码延胡索酸酶1(Fumarase 1,FUM1)和延胡索酸酶2(Fumarase 2,FUM2);定位于Chr.13上的基因Glyma.13G077800编码结节蛋白MTN21/EAMA样转运蛋白家族蛋白。此外,挖掘到5个与苹果酸含量显著相关的候选基因,其中,定位于Chr.05上的基因Glyma.05G112700编码铝激活苹果酸转运蛋白12(Aluminum-activated malate transporters 12,ALMT12);定位于Chr.16上的基因Glyma.16G176000和Glyma.16G176100分别编码UDP-糖基转移酶88A1(UDP-glucosyl transferase 88A1,UGT88A1)和SPX(SYG1/PHO81/XPR1)结构域蛋白;定位于Chr.19上的基因Glyma.19G188600和Glyma.19G189100分别编码真核天冬氨酸蛋白酶家族蛋白和泛素蛋白连接酶7(Ubiquitin-protein ligase 7,UPL7)。
3 结论与讨论
有机酸作为菜用大豆中重要的风味物质,对菜用大豆风味的形成具有显著的影响。目前,有机酸含量的检测通常采用酸碱滴定法、离子色谱法、气相色谱法和高效液相色谱法等。其中,高效液相色谱法可同时定量测定多种有机酸,且操作简便、准确度高、稳定性好,因此,被越来越多的利用于有机酸快速检测领域。本研究对264份菜用大豆种质资源进行有机酸含量测定,2020,2021年供试群体酒石酸含量分别为1.66~9.19 mg/g,0.48~13.52 mg/g,平均含量分别为4.13,4.16 mg/g;2020,2021年菜用大豆自然群体苹果酸含量分别为1.47~36.46 mg/g,1.89~25.19 mg/g,平均含量分别为7.26,8.99 mg/g;2020,2021年菜用大豆自然群体柠檬酸含量分别为2.56~12.05 mg/g,1.54~18.31 mg/g,平均含量分别为7.12,10.88 mg/g。2021年菜用大豆籽粒中3种有机酸平均含量最高的是柠檬酸,其次是苹果酸,这与Song等[14],Hu等[2]及张玉梅等[15]的研究结果一致。2 a中3种有机酸变异系数均大于10%,变异分析结果表明,供试菜用大豆种质资源遗传多样性丰富,有机酸含量存在非常广泛的表型变异,具有很大的遗传改良潜力。在此基础上筛选出6份高有机酸含量的特异种质资源,其中在2020,2021的检测中均属于高酒石酸含量组的种质为阜04-35和桂春豆112;2 a中均属于高苹果酸含量组的种质为华春9号和邯12-204;2 a中均属于高柠檬酸含量组的种质为灌豆3号和新沂大紫花,以上鉴定到的种质为菜用大豆有机酸品种改良育种提供优秀的材料。
基于MLM模型的全基因组关联分析2 a中共检测到了54个与酒石酸显著相关的SNP,189个与苹果酸显著相关的SNP以及43个与柠檬酸显著相关的SNP。但在2 a中3种有机酸均未鉴定到共同显著关联位点,这可能是因为有机酸是典型的数量性状,由多基因控制,含量极易受环境条件等因素的影响,因此,即便使用相同的自然群体,在不同环境条件下检测到一系列不同的关联位点的情况也是存在的[16]。此外,Hu等[2]在菜用大豆的籽粒中共鉴定出36种有机酸,其中含量最高的5种有机酸分别为柠檬酸、丁二酸、富马酸、苹果酸和草酰乙酸,本研究仅测定了酒石酸、苹果酸和柠檬酸这3种有机酸,而含量较高的丁二酸、富马酸还未进行研究,如果丁二酸和富马酸在年份之间有很大的变动,势必会引起其他有机酸的变化,从而会导致共定位的难度增大。因此,后续研究的重点一方面可以增加研究有机酸的种类,另一方面通过多年多点的不同环境试验,对菜用大豆自然群体有机酸含量进行检测,以期获得更加可靠的显著关联SNP位点,使有机酸遗传标记的开发和候选基因的发掘更加精确。以显著关联的SNP位点上下游120 kb范围内的基因为候选基因,根据基因功能注释信息,共鉴定到3个与酒石酸含量显著相关的候选基因及5个与苹果酸含量显著相关的候选基因。其中,Glyma.05G112700在拟南芥中的同源基因为AtALMT12,编码铝激活苹果酸转运蛋白12。在不同物种中均证实了ALMT家族基因能介导植物中苹果酸的转运,且参与果实中的苹果酸含量的调控,在果实酸化过程中发挥着重要作用[17-19],然而,对于该家族基因在菜用大豆中的功能及其调控机制尚不清楚。前人研究表明拟南芥中的AtALMT12调控苹果酸的运输,并且参与调节气孔运动[20],由此推测菜用大豆中的Glyma.05G112700基因可能与其具有相似的功能,与菜用大豆中苹果酸的含量具有相关性,但其调控机理还需进一步研究。综上所述,本试验结果可以为今后研究菜用大豆有机酸的基因位点提供参考依据,同时为候选基因的功能研究奠定基础,进而为菜用大豆育种提供参考借鉴。
