延胡索不同组织器官中内生真菌菌群结构与多样性分析
2023-08-18王梦迪黄情儿秦路平
吴 婷,王梦迪,黄情儿,朱 波*,秦路平*
延胡索不同组织器官中内生真菌菌群结构与多样性分析
吴 婷1, 2,王梦迪1,黄情儿1,朱 波1*,秦路平1*
1. 浙江中医药大学药学院,浙江 杭州 311402 2. 浙江省丽水市人民医院药学部,浙江 丽水 323000
明确延胡索不同组织器官中内生真菌菌群结构与多样性。通过形态和分子手段相结合的方法对延胡索块茎、叶、茎、根中的内生真菌进行分离、鉴定及菌群结构分析。共分离得到内生真菌2161株,根据菌株的形态学特征划分成106个形态型,分子鉴定为57个分类单元,归属到41个属,其中木霉属、毛链孢属为块茎的专有属,腐霉属、白僵菌属为叶的专有属,亚球壳属、核瑚菌属为茎的专有属,小囊菌属为根的专有属;相似性分析表明,延胡索不同组织器官内生真菌组成结构上存在差异;Shannon-wiener 多样性指数及Simpson指数分析显示延胡索根和茎中内生真菌多样性指数、丰度均显著高于块茎和叶。延胡索组织器官中内生真菌资源丰富,为延胡索内生真菌资源的开发利用提供了基础资料和参考依据。
延胡索;内生真菌;组织器官;菌群结构;多样性
延胡索W. T. Wang是罂粟科紫堇属植物,药用部位为干燥块茎,具有活血、行气、止痛的功效,可用于胸胁、脘腹疼痛、胸痹心痛、经闭痛经、产后瘀阻、跌扑肿痛[1]。延胡索分布于浙江、安徽、陕西、重庆、河南等地,临床应用广泛,市场需求巨大。现代研究还发现,延胡索具有催眠、抗心肌缺血、扩张冠状动脉、降压以及抗胃肠溃疡、抗肿瘤、抗炎、抗菌、抗病毒、提高抗应激能力等药理作用[2-3]。
植物内生真菌(plant endophytic fungi)是指能与植物体共生存,但不会引起植物产生明显病症的一类微生物[4]。已有研究表明,植物内生真菌遍及植物体当中,普遍分布在植物的根、茎、叶、块茎、花、果实和种子等组织器官或细胞间隙中,形成了植物体内的微生态体系[5]。
目前关于延胡索内生真菌的研究较少,马韦韦等[6]对陕西城固县的延胡索块茎进行分离鉴定,筛选出代谢产功能酶菌株;杨阳等[7]对8个产地延胡索块茎的内生真菌资源进行分离鉴定,发现块茎中存在丰富的内生真菌资源。已有研究发现,广东化橘红、麻花秦艽等菌群结构特征在不同组织部位存在差异性与特异性[8-9],且与药用植物品质密切相关。因此,本实验对内生真菌在延胡索块茎、叶、茎、根的菌群结构和多样性进行分析,明确延胡索不同组织器官内生真菌的分布,旨在为延胡索内生真菌的应用提供理论依据,为进一步开发延胡索内生真菌及提高延胡索药材品质奠定基础。
1 材料与试剂
1.1 材料
2021年4~5月,采收浙江、陕西、重庆、安徽、河南延胡索新鲜健康植株,分别为安徽宣城(S1)、重庆开县(S2)、陕西董家营(S3)、河南信阳(S4)、陕西三合镇(S5)、浙江缙云(S6)、浙江东阳(S7)、浙江磐安(S8),经浙江中医药大学药学院张巧艳教授鉴定为罂粟科多年生草本延胡索W. T. Wang。
1.2 试剂与仪器
PDA培养基(马铃薯200 g,葡萄糖20 g,琼20 g,1000 mL,pH自然);Takara9770A试剂盒购自美谷生物科技(浙江)有限公司;SW-CJ-1D型超净工作台(苏州净化有限公司);YXQ-LS-75S Ⅱ型立式压力蒸汽灭菌器(上海博讯有限公司);恒温培养箱、手术剪、手术刀、滤纸等。DYCP-31DN型琼脂糖水平电泳槽(北京六一有限公司);ABI Proflex型PCR(赛默飞世尔科技中国有限公司);GelDocXR型凝胶成像仪(北京科誉兴业科技发展有限公司);Eppendorf 5427R型离心机(Eppendorf中国有限公司)。无水乙醇(分析纯,国药集团),次氯酸钠(分析纯,国药集团),青霉素钠(上海索宝来有限公司)。
2 方法
2.1 不同形态型内生真菌菌株的分离与纯化
将收集的延胡索植株自来水下冲洗30 min除去表面的泥土和杂质,用无菌滤纸吸干其表面水分,进行3步表面消毒处理(75%乙醇1 min,2.5%次氯酸钠5 min,75%乙醇1 min),用无菌水洗涤延胡索植株表面残留酒精后再用无菌滤纸吸干其表面水分。接着用无菌手术刀将延胡索根、茎、叶、块茎切成小组织块(0.5 cm×0.5 cm×0.5 cm),置于含50 mg/L青霉素的PDA平板中,密封,25 ℃培养,定期观察。待内生真菌充分长出后,采用尖端挑取法进行纯化,直至出现形态单一的菌落。
2.2 不同形态型菌株的DNA提取与PCR扩增
采用Takara9770A试剂盒对内生真菌进行基因组DNA提取。PCR扩增采用引物ITS(5’-GGAA- GTAAAAGTCGTAAGG-3’)和ITS5(5’-TCCTCCG- CTTATTGATATGC-3’)进行PCR扩增。PCR反应体系(50 μL)组成:模板DNA,2 μL;引物ITS4,1 μL;引物ITS5,1 μL;2×TapPCR MasterMix(含染液),25 μL;ddH2O,21 μL。PCR反应循环参数与步骤:95 ℃初始变性3 min;95 ℃变性40 s;52 ℃退火50 s;72 ℃延伸 1 min;变性-退火-延伸循环35次;最后72 ℃延伸 10 min。扩增产物经琼脂糖凝胶电泳检测后送上海生工生物有限公司进行序列测定。
2.3 序列数据分析
登陆NCBI(http://www.ncbi.nlm.nih.gov/)利用BLAST将测序获得的各菌株IT S和5.8 S基因序列与GenBank中的已知序列进行比对,查找相似性最高的菌种,结合形态学特征,确定分离菌株的分类地位。
2.4 内生真菌多样性及相似性分析
定殖率指植物受到内生真菌侵染的组织块数占所有组织块数的百分比;分离率指分离得到的某个指定类型的内生真菌菌株数占分离样本组织块总数的百分比;相对频率(relative frequency,RF)表示所有样本中分离得到的某种内生真菌的菌株数占分离的总菌株数的百分比;运用Shannon-Wiener多样性指数(’)与Simpson多样性指数()来分析内生真菌菌群的生物多样性;运用Soenson相似性指数来计算比较不同组织器官之间内生真菌种类组成的相似性程度。优势属或优势目的判定方法为P>1/。P为内生真菌的菌株数占全部内生真菌菌株数的比值,为每个组织器官内生真菌种类数,当P>1/时即可判定为优势属或优势目[10]。
3 结果与分析
3.1 延胡索不同组织器官的内生真菌分布
从延胡索健康植株共分离得到内生真菌2161株,其中根109株、茎114株、叶196株,块茎1742株,见表1。4个组织器官内生真菌的定殖率和分离率,见表2。结果表明,不同组织器官的内生真菌的定殖率和分离率均具有统计学意义。块茎、叶、茎、根的定殖率高低为块茎>叶>茎>根,分离率高低为块茎>叶>茎>根,表明延胡索植株不同组织器官中内生真菌的侵染程度与丰富程度存在显著差异。
表1 延胡索中内生真菌的分离
Table 1 Endophytic fungi isolated from C. yanhusuo
代号分类参考序列相似度/%分离数量/株总计 S1S2S3S4S5S6S7S8 T5腐皮镰刀菌Fusarium solaniMK027264100.00108(Rh)61(Rh)79(Rh)98(Rh)100(Rh)70(Rh)79(Rh)50(Rh)645(Rh) T7木贼镰刀菌Fusarium equisetiMK764999100.0069(Rh),2(Ra)56(Rh)50(Rh),2(Fo)41(Rh),2(Ra)50(Rh),1(Ra)49(Rh), 2(Ca),6(Ra)63(Rh)30(Rh),2(Fo),3(Ca)408(Rh),4(Fo),5(Ca),11(Ra) T79锐顶镰刀菌Fusarium acuminatumMH782585100.00016(Rh)03(Ra)5(Rh),2(Ra)9(Rh)6(Rh),2(Ca)8(Rh)44(Rh),2(Ca),5(Ra) T89层出镰刀菌Fusarium proliferatumMH933697100.0000007(Rh)11(Rh)0018(Rh) T97粉红螺旋聚饱霉Clonostachys roseaMH911376100.002(Ra)71(Rh)41(Rh)12(Rh)30(Rh)20(Rh),3(Ra)22(Rh)21(Rh)217(Rh),5(Ra) T83木霉属Trichoderma sp.KU93347589.1601(Ca)8(Rh)004(Rh),2(Ca)9(Rh)10(Rh)31(Rh),3(Ca) T86Volutella consorsMH89258197.96014(Rh)0004(Rh)6(Rh)024(Rh) Y81紧密帚枝霉Sarocladium strictumMF497404100.00000003(Fo)003(Fo) Y92球孢白僵菌Beauveria bassianaMG34508199.380000002(fo)02(fo) T25Monilochaetes melastomaeNR16112497.6649(Rh)26(Rh)27(Rh)24(Rh)20(Rh)31(Rh)27(Rh)20(Rh)224(Rh) G94黄瓜织球壳菌Plectosphaerella cucumerinaMH855387100.00003(Ca)0001(Ra)03(Ca),1(Ra) J86变黑轮枝菌Gibellulopsis nigrescensMH856082100.00003(Ca)000003(Ca) T17球毛壳Chaetomium globosumMK120280100.000000016(Rh)6(Rh)20(Rh),10(Ra)42(Rh),10(Ra) Y77Chaetomium lentumMH861858100.000007(fo)00007(fo) J88Arcopilus aureusMG889930100.003(Ca)00000019(Ra)3(Ca),19(Ra) G78Dichotomopilus variostiolatusMG889985100.004(Ra)00000004(Ra) T11菜豆间座壳Diaporthe phaseolorumMH78258999.04000000011(Rh)11(Rh) T99Microdochium bolleyiMH860598100.000009(Rh)00009(Rh) Y89Peroneutypa scopariaMK99342299.6100000002(Fo)2(Fo) Y82帚霉属Scopulariopsis sp.LM65247972.6600002(Fo)0002(Fo) G81Microascus alveolarisKX923834100.001(Ra)00000001(Ra) T15Ophiosphaerella agrostidisKM43427298.5500000007(Rh)7(Rh) T22土栖棘壳孢Setophoma terrestrisKY56133799.1700002(Ra)3(Ca)6(Rh)06(Rh),3(Ca),2(Ra) Y75菊异茎点霉Paraphoma chrysanthemicolaMH24508299.7901(Fo)2(Fo)000003(Fo) Y10Stemphylium solainMH14245699.59000001(Fo)001(Fo) Y85新西兰匍柄霉Stemphylium eturmiunumMH843731100.0000002(Fo)0002(Fo) Y95链格孢菌Alternaria alternataMK881030100.0096(Fo),16(Ca)03(Fo),3(Ca)9(fo),1(Ca)2(Fo),2(Ra)4(Ra)02(Fo),1(Ca)112(Fo),21(Ca),6(Ra) Y96极细链格孢霉Alternaria tenuissimaKF99674899.3920(Ca)2(Ca)6(Fo),4(Ca),1(Ra)15(fo),2(Ca)2(Fo),1(Ca)4(Ra)5(fo),2(Ca)2(Ca)28(fo),33(Ca),5(Ra)
续表1
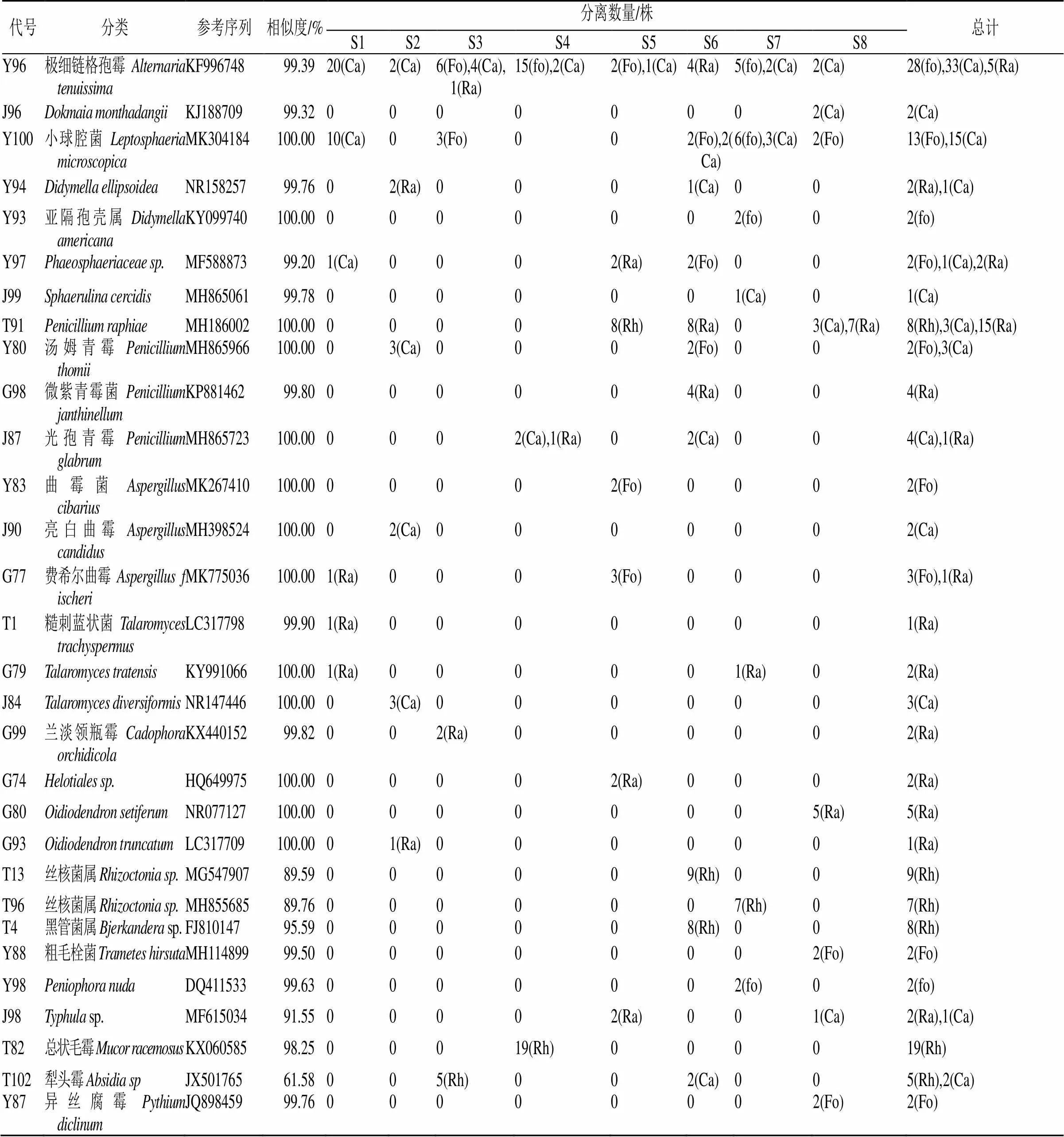
代号分类参考序列相似度/%分离数量/株总计 S1S2S3S4S5S6S7S8 Y96极细链格孢霉Alternaria tenuissimaKF99674899.3920(Ca)2(Ca)6(Fo),4(Ca),1(Ra)15(fo),2(Ca)2(Fo),1(Ca)4(Ra)5(fo),2(Ca)2(Ca)28(fo),33(Ca),5(Ra) J96Dokmaia monthadangiiKJ18870999.3200000002(Ca)2(Ca) Y100小球腔菌Leptosphaeria microscopicaMK304184100.0010(Ca)03(Fo)002(Fo),2(Ca)6(fo),3(Ca)2(Fo)13(Fo),15(Ca) Y94Didymella ellipsoideaNR15825799.7602(Ra)0001(Ca)002(Ra),1(Ca) Y93亚隔孢壳属Didymella americanaKY099740100.000000002(fo)02(fo) Y97Phaeosphaeriaceae sp.MF58887399.201(Ca)0002(Ra)2(Fo)002(Fo),1(Ca),2(Ra) J99Sphaerulina cercidisMH86506199.780000001(Ca)01(Ca) T91Penicillium raphiaeMH186002100.0000008(Rh)8(Ra)03(Ca),7(Ra)8(Rh),3(Ca),15(Ra) Y80汤姆青霉Penicillium thomiiMH865966100.0003(Ca)0002(Fo)002(Fo),3(Ca) G98微紫青霉菌Penicillium janthinellumKP88146299.80000004(Ra)004(Ra) J87光孢青霉Penicillium glabrumMH865723100.000002(Ca),1(Ra)02(Ca)004(Ca),1(Ra) Y83曲霉菌Aspergillus cibariusMK267410100.0000002(Fo)0002(Fo) J90亮白曲霉Aspergillus candidusMH398524100.0002(Ca)0000002(Ca) G77费希尔曲霉Aspergillus f ischeriMK775036100.001(Ra)0003(Fo)0003(Fo),1(Ra) T1糙刺蓝状菌Talaromyces trachyspermusLC31779899.901(Ra)00000001(Ra) G79Talaromyces tratensisKY991066100.001(Ra)000001(Ra)02(Ra) J84Talaromyces diversiformisNR147446100.0003(Ca)0000003(Ca) G99兰淡领瓶霉Cadophora orchidicolaKX44015299.82002(Ra)000002(Ra) G74Helotiales sp.HQ649975100.0000002(Ra)0002(Ra) G80Oidiodendron setiferumNR077127100.0000000005(Ra)5(Ra) G93Oidiodendron truncatumLC317709100.0001(Ra)0000001(Ra) T13丝核菌属Rhizoctonia sp.MG54790789.59000009(Rh)009(Rh) T96丝核菌属Rhizoctonia sp.MH85568589.760000007(Rh)07(Rh) T4黑管菌属Bjerkandera sp.FJ81014795.59000008(Rh)008(Rh) Y88粗毛栓菌Trametes hirsutaMH11489999.5000000002(Fo)2(Fo) Y98Peniophora nudaDQ41153399.630000002(fo)02(fo) J98Typhula sp.MF61503491.5500002(Ra)001(Ca)2(Ra),1(Ca) T82总状毛霉Mucor racemosusKX06058598.2500019(Rh)000019(Rh) T102犁头霉Absidia spJX50176561.58005(Rh)002(Ca)005(Rh),2(Ca) Y87异丝腐霉Pythium diclinumJQ89845999.7600000002(Fo)2(Fo)
Ra-根 Ca-茎 Fo-叶 Rh-块茎
Ra-root Ca-stem Fo-leaf Rh-tuber
表2 不同组织器官内生真菌定殖率与分离率
Table 2 Colonization rate and isolation rate of endophytic fungi in different tissues of C. yanhusuo
组织定殖率/%分离率/% 根10.6112.04 茎10.6211.88 叶17.0820.41 块茎83.65 181.45
根据菌株的培养特征将其划分为106个形态型,通过5.8S和ITS区片段序列的相似性对比和系统学分析,其中有57个鉴定到种,41个鉴定到属,30个鉴定到科,18个鉴定到目。66.67%的延胡索内生真菌表现出组织特异性,但未发现4个部位共有菌种。
延胡索内生真菌的分布具有明显的组织特异性。在目的分类水平上,块茎的优势目为肉座菌目(Hypocreales,RF=79.62%)和球团目(Glomerellales,RF=12.86%);叶的优势目为腔菌目(Pleosporales,RF=82.14%);茎的优势目为腔菌目(Pleosporales,RF=65.79%)、散囊菌目(Eurotiales,RF=13.16%)、肉座菌目(Hypocreales,RF=8.77%);根的优势目为腔菌目(Pleosporales,RF=13.76%)、肉座菌目(Hypocreales,19.27%)、散囊菌目(Eurotiales,RF=22.02%)、粪壳菌目(Sordariales,RF=30.28%)。在属的分类水平上,块茎的优势属为粘帚霉属(RF=12.46%)、镰刀菌属Link ex Fr.(RF=64.01%)、毛链孢属Halst. ex Harter(RF=12.86%);叶的优势属为链格孢属Nees(RF=71.43%)、小球腔菌属Ces.(RF=6.63%);茎的优势属Nees为链格孢属(RF=47.37%)、小球腔菌属Ces.(RF=13.16%)、镰刀菌属Link ex Fr.(RF=6.14%);根的优势属为链格孢属Nees(RF=10.09%)、镰刀菌属Link ex Fr.(RF=14.68%)、青霉菌属Link(RF=18.35%)、(RF=17.43%)、毛壳属(RF=9.17%)(图1和图2)。其中木霉属Pers.、毛链孢属Halst.ex Harter为块茎的专有属,腐霉属Pringsh.、白僵菌属Vuillemin为叶的专有属,亚球壳属Sacc.、核瑚菌属(Pers.) Fr.为茎的专有属,小囊菌属为根的专有属。
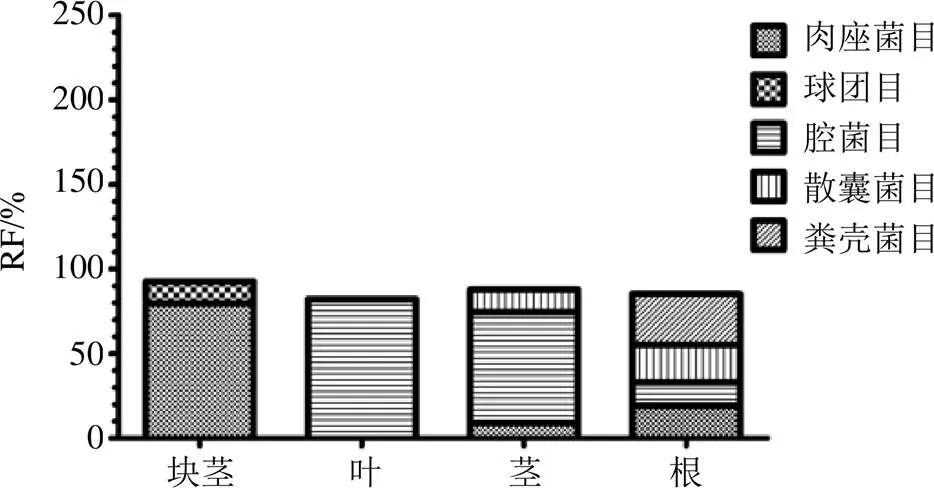
图1 4个组织器官中优势目的RF值
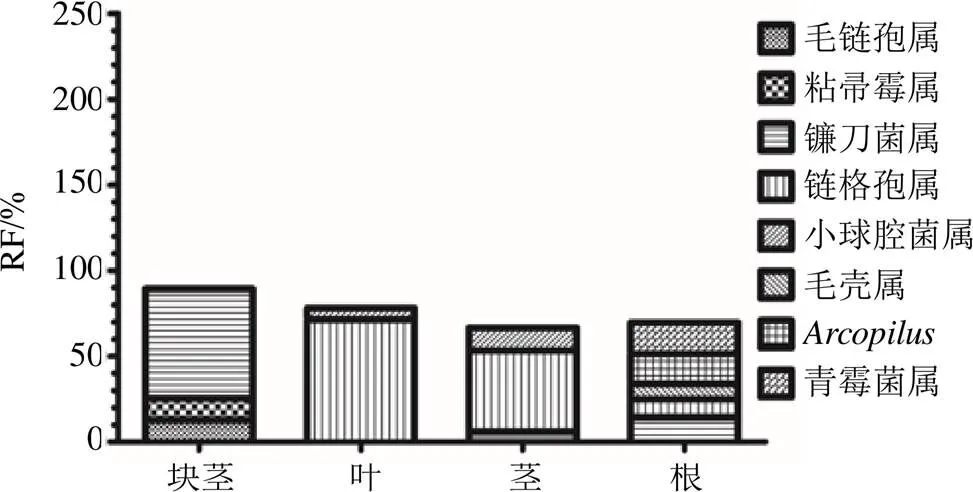
图2 4个组织器官中优势属的RF值
3.2 内生真菌多样性及丰度分析
方差分析和结果表明,4个组织器官延胡索内生真菌丰度和值具有极显著差异,多样性指数(’)具有显著性差异,见表3。延胡索根和茎中内生真菌多样性指数、丰度均明显高于块茎和叶,根中的内生真菌’是叶的1.62倍。这表明植物不同组织器官内生真菌菌群的分布及组成不同。
表3 4个组织器官多样性指数和丰度
Table 3 Diversity index and relative frequency of endophytic fungi in different tissues of C. yanhusuo
组织SH’D 块茎191.857 50.773 8 叶201.709 60.645 1 茎212.407 70.854 7 根242.767 50.913 3
3.3 不同组织器官内生真菌相似性比较
相似性指数(s)用于比较2个不同组织器官的内生真菌种类组成的相似程度。在种水平上,不同组织器官的s在0.051 3~0.577 8,茎部与根部内生真菌菌群相似度最高,块茎部与叶部相似性最低;在属的水平上,不同组织器官的s在0.062 5~0.666 7,茎部与根部相似度最高,块茎部和叶部相似性最低。总的来看,各组织器官内生真菌类群之间存在较大的差异,组织差异性明显,见表4和表5。
4 讨论
本实验从延胡索块茎、叶、茎、根4个组织器官中分离得到2161株内生真菌,通过5.8 S和ITS区片段序列的相似性对比和系统学分析,鉴定归属为41个属,显示出丰富的内生真菌多样性。在延胡索不同的组织器官中,茎部与根部内生真菌菌群相似度最高,块茎部与叶部相似性最低。内生真菌的分布规律可能与宿主本身的特性及内生真菌的种类相关,因此在宿主的不同组织器官中具有不同的分布和结构。延胡索各组织器官既有特殊的菌群,也有广泛存在的菌群,体现了延胡索不同组织器官内生真菌的特异性和差异性,原因可能是各组织器官的结构差异和营养物质的差异影响了内生真菌的种类、侵染能力、方式与生存能力。
表4 不同组织器官在种水平上的内生真菌Cs
Table 4 Similarity index of endophytic fungi in different tissues of C. yanhusuo at species level
组织器官Cs 块茎叶茎根 块茎1.000 0 叶0.051 31.000 0 茎0.300 00.292 71.000 0 根0.279 10.227 30.577 81.000 0
表5 不同组织器官在属水平上的内生真菌Cs
Table 5 Similarity index of endophytic fungi in different tissues of C. yanhusuo at generic level
组织器官Cs 块茎叶茎根 块茎1.000 0 叶0.062 51.000 0 茎0.312 50.294 11.000 0 根0.294 10.222 20.666 71.000 0
链格孢属、镰刀菌属、粘帚霉属、毛链孢属、毛壳属为延胡索的优势菌株。优势属中,镰刀菌属和毛链孢属内生真菌可引起植物病害,而粘帚霉属和毛壳属内生真菌已经广泛用于生物防治。链格孢属广泛存在于土壤及植物组织中,在枸杞[11]、铁皮石斛[12]等多种植物中都有发现,链格孢属是生态系统中最为常见的病原菌,但同时也是一种新的微生物资源,在抑菌、杀虫和抗原生生物方面发挥着重要作用,蕴藏巨大潜在应用价值[13]。镰刀菌属会侵染多种植物,引起严重的根腐病,造成巨大的经济损失[14]。目前对毛链孢属的研究较少,赵永强等[15]发现毛链孢属能导致甘薯黑皮病。近年来,人们利用粘帚霉属真菌对立枯丝核菌、腐霉菌等土传植物病原菌的抗菌、溶解、竞争、寄生等作用来防治它们对植物造成的脱落、根腐等病害。在国外,链孢粘帚霉和绿粘帚霉已被开发成生防制剂产品[16]。毛壳属真菌是常见的内生真菌类群,也是具有重要应用价值的资源真菌,其能够通过诱导宿主植物产生系统抗性、重寄生作用以及产生次级代谢产物拮抗病原菌等方式防治植物病害[17]。
不同产地间延胡索内生真菌多样性存在差异,浙江缙云产延胡索内生真菌多样性指数最高,’达2.734 8;而安徽宣城多样性最低,’值仅为1.729 7。菌群结构方面,各产地延胡索最优势属均为镰刀菌属,其他优势属略有区别,但以毛链孢属、链格孢属、粘帚霉属为主。供试8个产地延胡索均存在产地特有菌群,如安徽宣城延胡索特有菌为(RF=1.04%)、(RF=0.26%)等;河南信阳特有菌为(RF=2.86%)、(RF=3.67%)等;浙江缙云特有菌为草莓花枯病菌(RF=3.17%)、烟管菌(RF=2.82%)等。这些产地特异性菌群可能与该产地土壤、降水量、湿度、温度等生境因子相关,为延胡索道地性形成机制提供了参考依据。
根据’、指数表明延胡索根、块茎中的内生真菌种类较丰富。延胡索的药用部位是块茎,木霉属为块茎的特有种群,杨阳等[7]研究发现木霉属内生真菌T83与延胡索镇痛活性成分延胡索乙素积累存在密切关系。木霉菌被认为是防治真菌性植物病害的潜在生防菌,它们可以直接与根相互作用,增加植物生长,抵抗疾病和对非生物胁迫的耐受性[18]。因此,下一步的工作中将重点研究延胡索内生真菌对宿主植物生长发育的作用,对药用部位活性物质形成、含量变化的影响。目前,关于内生真菌对药用植物活性成分的影响仍处于探索阶段,本研究通过对延胡索不同组织器官进行内生真菌群落结构分析,为进一步研究内生真菌与延胡索药材品质关系奠定了良好的基础。
利益冲突 所有作者均声明不存在利益冲突
[1] 中国药典[S]. 一部. 2020: 145-146.
[2] 李雅萌, 王亚茹, 周柏松, 等. 延胡索中重金属及有害元素的测定 [J]. 特产研究, 2019, 41(2): 66-68.
[3] 杨梦萍, 陈晓瑶, 张悦健, 等. 基于文献计量学的延胡索研究现状及热点分析 [J]. 药物评价研究, 2023, 46(7): 1406-1416.
[4] 张志东, 朱静, 唐琦勇, 等. 不同扩增引物对高通量测序分析盐角草内生真菌多样性的影响 [J]. 微生物学通报, 2020, 47(2): 411-419.
[5] 李梦歌, 惠建超, 阿依佳玛丽·依玛尔, 等. 陕西宜君核桃内生真菌多样性研究 [J]. 云南农业大学学报: 自然科学, 2018, 33(5): 818-825.
[6] 马韦韦, 邓百万, 陈文强. 延胡索内生真菌的分离鉴定及产功能酶菌株的初步筛选 [J]. 陕西理工学院学报: 自然科学版, 2016, 32(4): 65-72.
[7] 杨阳, 李茜茜, 唐藕凤, 等. 延胡索块茎中内生真菌菌群结构及其与延胡索乙素含量的相关性研究 [J]. 中草药, 2020, 51(7): 1901-1908.
[8] 舒永馨, 罗梅, 陈欣瑜, 等. 广东化橘红内生真菌多样性分析 [J]. 仲恺农业工程学院学报, 2021, 34(1): 8-11.
[9] 陈昕, 李琪, 曹倩倩, 等. 麻花秦艽不同组织部位可培养内生菌群结构及其与龙胆苦苷含量的相关性 [J]. 食品与生物技术学报, 2019, 38(4): 21-29.
[10] 吴令上. 南方红豆杉内生真菌多样性、次生代谢产物及其与宿主的相关性研究 [D]. 上海: 第二军医大学, 2012.
[11] 刘伟, 张勤德, 王多文, 等. 枸杞新病害叶枯病症状表现及病原菌鉴定 [J]. 北方园艺, 2018(12): 141-145.
[12] 刘国华, 赵楠, 赵桂华. 铁皮石斛枯梢病的病原菌及发病规律研究 [J]. 西南林业大学学报, 2017, 37(1): 138-143.
[13] 周素婷, 任兵杨, 李欲轲, 等. 链格孢致病机制、防治措施与开发利用研究进展 [J/OL]. 分子植物育种, 2021. [2021-10-15]. https://kns.cnki.net/kcms/detail/ 46.1068.S.20211014.1715.008.html.
[14] 文增叶, 李定华, 代梦瑶, 等. 三七根腐病病原菌尖孢镰刀菌的生物学特性分析 [J]. 中药材, 2019, 42(9): 1978-1984.
[15] 赵永强, 徐振, 杨冬静, 等. 甘薯黑痣病菌的生物学特性研究 [J]. 北方农业学报, 2018, 46(5): 89-92.
[16] 董锦艳, 李铷, 张克勤. 粘帚霉属真菌代谢物的研究进展 [J]. 微生物学通报, 2006, 33(2): 124-131.
[17] 唐保宏. 玉米叶斑病拮抗性毛壳属内生真菌的筛选 [A] // 中国菌物学会第七届全国会员代表大会暨2017年学术年会摘要集 [C]. 宜昌: 中国菌物学会, 2017: 202.
[18] Silva R N, Monteiro V N, Steindorff A S,. Trichoderma/pathogen/plant interaction in pre-harvest food security [J]., 2019, 123(8): 565-583.
Fungal community and diversity of endophytic fungi in different tissues of
WU Ting1, 2, WANG Meng-di1, HUANG Qing-er1, ZHU Bo1, QIN Lu-ping1
1. School of Pharmaceutical Sciences, Zhejiang Chinese Medical University, Hangzhou 311402, China 2. Department of Pharmacy, Lishui People's Hospital, Lishui 323000, China
To clarify fungal community and diversity of endophytic fungi in different tissues of.Endophytic fungi in tubers, leaves, stems and roots ofwere isolated, identified and analyzed by morphological and molecular methods.A total of 2162 endophytic fungal strains were isolated and divided into 106 morphological types. Through molecular systematical analysis, they were divided into 57 taxons, belonging to 41 genera, whileandwere the exclusive genera of tubers,andwere the exclusive genera of leaves, andandwere the exclusive genera of stems,is a proprietary genus in roots. Similarity analysis showed that there were significant differences in the endophytic fungal community in different tissues. Shannon Wiener diversity index and Simpson index analysis showed that the diversity index and abundance of endophytic fungi in roots and stems were significantly higher than those in tubers and leaves.There are abundant endophytic fungi in varioustissues. The paper provides basic information and scientific reference for the development and utilization of endophytic fungal resources in.
W. T. Wang; endophytic fungi; tissues; fungal community; diversity
R286.2
A
0253 - 2670(2023)16 - 5372 - 07
10.7501/j.issn.0253-2670.2023.16.025
2023-03-03
浙江中医药大学人才专项(2021ZR09);浙江省医药卫生青年创新人才支持计划(2022RC052)
吴 婷(1983—),女,在读研究生,研究方向为植物内生菌。E-mail: 513303983@qq.com
朱 波,博士,副研究员,硕士生导师,研究方向为植物内生菌。E-mail: zhubo@zcmu.edu.cn
秦路平,博士,教授,博士生导师,研究方向为中药资源及品质评价。E-mail: lpqin@zcmu.edu.cn
[责任编辑 时圣明]
