睡眠剥夺对豚鼠离焦性近视及视网膜多巴胺代谢的影响
2023-07-26杨雪莉陈尧关宇欣袁超群毛俊峰
杨雪莉 陈尧 关宇欣 袁超群 毛俊峰
作者单位:中南大学湘雅医院眼科中心 眼科学湖南省重点实验室,长沙 410008
青少年近视率在全球范围内呈逐年增长趋势,已成为影响青少年身心成长的重大公共卫生问题,造成的社会经济负担不容忽视[1]。2018年7 月,教育部发布我国首份《中国义务教育质量检测报告》,四、八年级学生视力不良检出率为36.5%、65.3%,部分地区四年级超过60%、八年级超过80%,其中近视眼占比90%以上。研究发现,近视对青少年的生理功能及情感功能具有负向影响,生存质量显著下降,不利于身心健康发展[2]。关于睡眠不足与近视的关系,宋逸等[3]对我国7~22岁261 832名汉族青少年进行调查研究,发现睡眠时间不足的青少年更易出现近视;Jee等[4]在韩国进行的一项横断面研究也证实,睡眠时间与青少年近视呈负相关,睡眠时间每增加1 h、近视风险降低10%。可见,睡眠不足与青少年近视的形成密切相关,但睡眠不足是否影响近视形成及具体机制尚不清楚。因此,本研究利用已建立的豚鼠透镜诱导性近视(Lens-induced myopia,LIM)模型,每天给予睡眠剥夺(Sleep deprivation,SD)干预,通过观察3,4-二羟基苯乙酸(3, 4-dihydroxyphenylacetic acid,DOPAC)和酪氨酸羟化酶(Tyrosine hydroxylase,TH)研究SD对豚鼠LIM模型中近视及视网膜近视相关信号因子多巴胺(Dopamine,DA)的影响,为探讨SD与青少年近视形成的关系提供科学依据。
1 材料与方法
1.1 材料
1.1.1 实验动物 2022 年3 月选取30 只21 日龄健康三色豚鼠(中南大学实验动物学部提供),体质量140~170 g,性别不限。所有豚鼠饲养在中南大学实验动物中心,保持室温25 ℃,维持12 h光照/12 h循环(7:00AM—7:00PM),光照在480~520 Lx之间,予以正常视觉环境或LIM处理,可自由进食饮水。本研究获中南大学实验动物学部伦理委员会批准(批号:2020sydw0084),实验动物的使用和喂养遵循中国科学技术委员会颁布的《实验动物管理条例》。
1.1.2 主要试剂及仪器 DA标准品(#H8502,美国Sigma-Aldrich公司)、DOPAC标准品(#850217,美国Sigma-Aldrich公司)、兔抗TH多克隆抗体(ab137721,美国Abcam公司)、兔抗β-actin多克隆抗体(ab8227,美国Abcam公司)、BCA蛋白定量试剂盒(#C05-02001,北京博奥森生物技术有限公司)。AUT-SD-1S动物睡眠剥夺仪(南京奥腾工程技术有限公司,原理:在实验笼内采用间歇性剥夺杆旋转进行触觉刺激干扰自由行为的豚鼠实现SD)、-6.00 D光学树脂凹透镜片(直径为12 mm,内弧曲率为9.61 mm,香港明达眼镜镜片有限公司)、ARK-510A电脑验光角膜曲率计(日本尼德克公司)、MD-2400S眼科A/B型超声诊断仪(天津迈达医学科技股份有限公司)、艾沃斯-V8 数字式照度计(武汉中测宏图测量仪器有限公司)等。
1.2 方法
1.2.1 分组处理 采用随机数表法将豚鼠分为LIM组和LIM+SD组,每组15只。所有豚鼠右眼为离焦眼,配戴-6.0 D凹透镜片14 d,制作LIM模型,对侧未处理眼为自身对照眼。LIM+SD组豚鼠在配戴凹透镜的同时,给予每天持续SD20 h(7:00AM—3:00AM)处理。研究SD对豚鼠离焦眼屈光发育和视网膜DA代谢的影响。
1.2.2 豚鼠LIM模型的制作 按照本课题组豚鼠LIM模型的建立方法[5]制作LIM模型,以右眼为实验眼,把自制镜框缝合固定于眶周软组织,-6.00 D凹透镜片插入固定于镜框之中,每日擦拭镜片以保持其清洁。配戴凹透镜片14 d后,采用电脑验光角膜曲率计测定角膜曲率半径(精确到0.01 mm)和眼球屈光度,A型超声仪测量眼轴长度(精确到0.01 mm),每眼测量3次,取平均值,评估眼球屈光状态变化。随后,腹腔注射过量戊巴比妥钠处死豚鼠,摘除眼球。
1.2.3 SD干预方式 LIM+SD组豚鼠放入AUT-SD-1S动物睡眠剥夺仪中,每个剥夺仪箱体内放入5只豚鼠。箱体材料是透明有机玻璃,圆柱形,直径50 cm,高40 cm,底部是46 cm长的旋转金属杆,金属杆转速可以调节。箱体内照明达到LIM造模条件,每天用数字式照度计测量箱体内豚鼠眼水平面的光照度,控制在480~520 Lx之间,箱内可以自由饮水及进食。金属杆转动模式控制在每间隔10 min旋转3圈,转速为1圈15 s,制作豚鼠SD模型。在配戴凹透镜期间每天持续SD20 h,结束后移入正常饲养箱。
1.2.4 神经视网膜DA、DOPAC含量的测定 每组各取8只豚鼠16个眼球,用于高效液相色谱电化学法测定神经视网膜DA和DOPAC的含量(ng/mg),具体方法参见本课题组以前的研究[6],并计算DOPAC/DA比值。操作大致如下:Hypersil ODS2色谱柱(5 μm,250 mm × 4.6 mm),流动相流速20 mmol/L,柠檬酸三钠(含5 mmol/L庚烷磺酸和0.1 mmol/LEDTA,pH值为3.7),甲醇(体积比90:7),流速1.0 ml/L,柱温箱温度35 ℃,电化学检测器工作电压+750 mV,进样量20 μL。
1.2.5 视网膜TH蛋白分布的检测 每组各取2只豚鼠4 个眼球进行视网膜TH蛋白检测。将眼球置于10%中性甲醛中固定,石蜡包埋,制作5 μm厚切片,用于免疫组织化学染色,一抗是兔抗TH多克隆抗体(1:200)。染色后切片,采集图像。
1.2.6 视网膜TH蛋白表达的检测 每组各取5 只豚鼠10个眼球用于Western blot检测。取视网膜神经上皮层标本,称质量,剪碎。加入细胞裂解液匀浆,离心后取上清液冻存。BCA蛋白定量试剂盒进行蛋白定量。取蛋白样品25 μg,进行Western blot检测,一抗是兔抗TH多克隆抗体(1:1 000)。利用Bandscan5.0 图象分析软件对目的条带进行灰度分析,以β-actin作内参,计算TH蛋白的相对表达量。TH蛋白相对表达量=TH条带灰度值/β-actin条带灰度值。
1.3 统计学方法
实验研究。通过SPSS 22.0统计学软件进行数据处理。所有数据经正态性和方差齐性检验,符合正态分布,以±s表示。豚鼠左右眼角膜曲率半径、眼球屈光度、眼轴长度、视网膜DOPAC含量、DA含量及DOPAC/DA比值、视网膜TH蛋白相对表达量的比较采用配对样本t检验,LIM组与LIM+SD豚鼠组间角膜曲率半径、屈光度、DOPAC含量和DA含量等屈光参数和神经递质含量比较采用独立样本t检验。离焦眼视网膜DA含量分别与眼球屈光度、眼轴长度进行相关分析,计算Pearson相关系数。以P<0.05为差异有统计学意义。
2 结果
2.1 SD对豚鼠离焦性近视眼屈光状态的影响
L I M 组豚鼠自身对照眼出现轻度远视(1.30±0.65)D,眼轴长度为(7.98±0.08)mm,角膜曲率为(3.56±0.05)mm。与自身对照眼比较,LIM组离焦眼眼轴延长为(8.36±0.08)mm(t=70.33,P<0.001),形成近视眼为(-1.87±0.72)D,相对近视度数为3.17 D(t=-30.04,P<0.001)。LIM+SD组和LIM组之间离焦眼相比较,LIM+SD组眼轴延长更大(8.48±0.09)mm、近视程度更深(-3.47±0.83)D,差异均有统计学意义(t=-3.87,P=0.001;t=5.63,P<0.001)。LIM+SD组和LIM组之间自身对照眼相比较,眼轴长度(8.01±0.06)mm和屈光度(+1.23±0.68)D,差异均无统计学意义(均P>0.05)。角膜曲率半径在LIM组和LIM+SD组的组间及组内比较,差异均无统计学意义(均P>0.05)。
2.2 SD对豚鼠离焦性近视眼视网膜DA、DOPAC含量的影响
LIM组豚鼠自身对照眼视网膜DA、DOPAC含量分别为(1.55±0.32)ng/mg、(0.92±0.17)ng/mg,与LIM+SD组自身对照眼比较,差异均无统计学意义(P>0.05)。与自身对照眼相比,LIM组及LIM+SD组离焦眼的视网膜DA、DOPAC含量均降低,差异有统计学意义(LIM组:t=-8.64,P<0.001;t=-13.03,P<0.001;LIM+SD 组:t=-11.72,P<0.001;t=-16.32,P<0.001)。LIM组和LIM+SD组组间的DA、DOPAC差异均有统计学意义(t=5.35,P<0.001;t=3.80,P=0.002)。豚鼠离焦眼视网膜DA含量与屈光度(r=0.93,P<0.001)、眼轴长度(r=-0.77,P=0.001)均有相关性。
视网膜DOPAC/DA比值在LIM组和LIM+SD组自身对照眼之间比较,差异无统计学意义(P>0.05)。LIM组和LIM+SD组离焦眼视网膜DOPAC/DA比值均低于自身对照眼,差异有统计学意义(LIM组:t=-9.55,P<0.001;LIM+SD组:t=-6.58,P<0.001)。与LIM组比较,LIM+SD组离焦眼视网膜DOPAC/DA比值升高,差异有统计学意义(t=-3.60,P=0.003)。
2.3 SD对豚鼠离焦性近视眼视网膜TH蛋白表达的影响
免疫组织化学染色结果显示,TH阳性反应细胞主要分布在视网膜神经纤维层、神经节细胞层、内丛状层和内核层,外丛状层可见少量着色;与自身对照眼相比,LIM组和LIM+SD组离焦眼视网膜TH蛋白均减弱(见图1)。Western blot检测发现,LIM组和LIM+SD组视网膜TH蛋白表达均低于自身对照眼,差异有统计学意义(LIM组:t=-40.25,P<0.001;LIM+SD组:t=-17.64,P<0.001);但2组离焦眼之间视网膜TH蛋白的差异无统计学意义(P=0.333),自身对照眼之间的差异也无统计学意义(P=0.246)(见图2)。
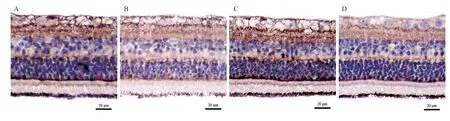
图1.免疫组织化学染色法检测豚鼠视网膜TH蛋白表达Figure 1.Immunohistochemical staining of TH protein in the retina of guinea pigs
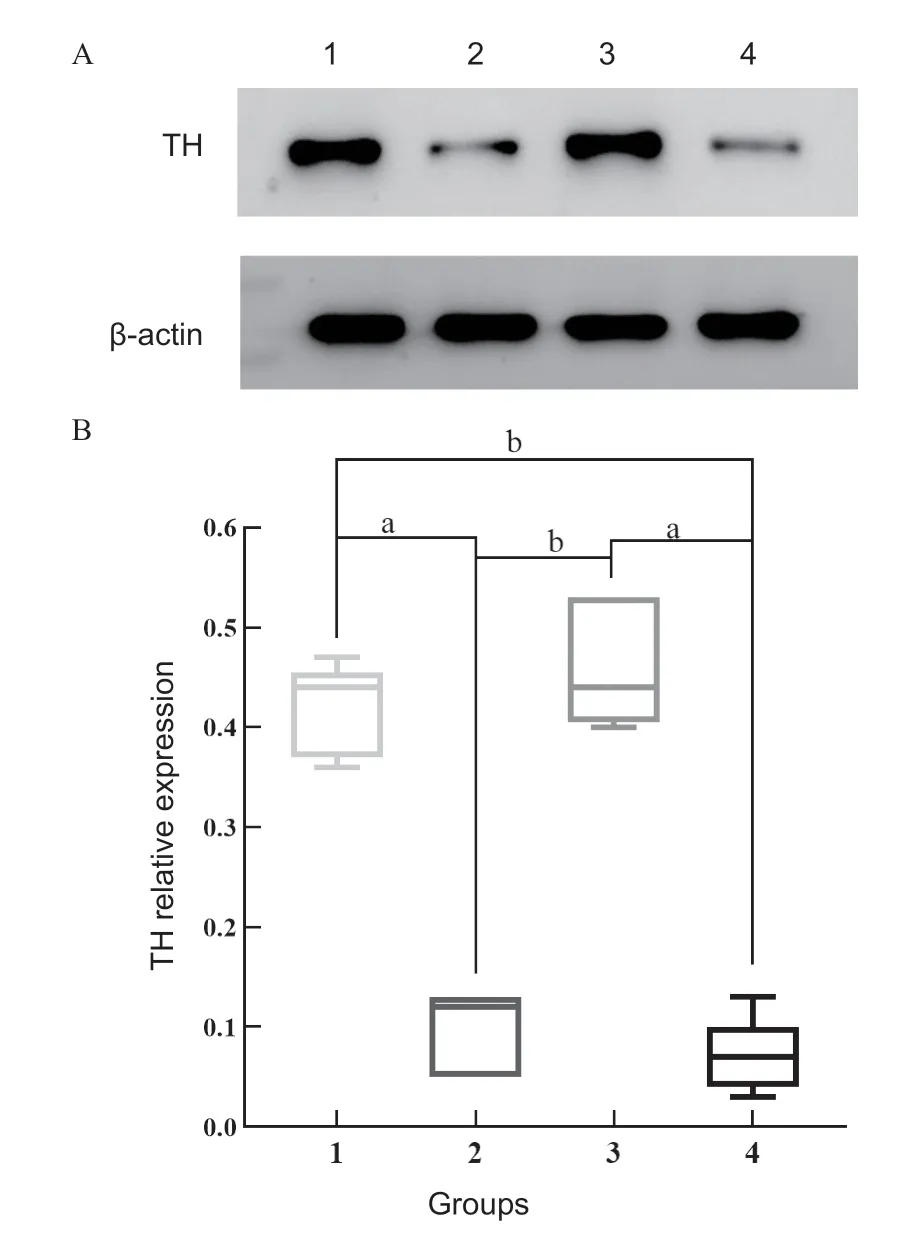
图2.SD对豚鼠视网膜TH蛋白含量的影响Figure 2.Effects of SD on TH proteins in the retina of guinea pigs
3 讨论
目前啮齿类动物广泛应用于睡眠研究当中,豚鼠出生时其神经系统发育相对完善,1 周后睡眠-觉醒周期即呈现成年豚鼠的睡眠模式,常用于睡眠的神经生理学及睡眠-觉醒机制研究。予以豚鼠SD 6 h即可表现为明显的非快速眼动睡眠增加,慢波睡眠增强,睡眠碎片化程度增加[7]。我们在已有的LIM模型基础上,每天给予SD 20 h,建立豚鼠LIM+SD模型,发现SD可显著加深离焦眼近视程度,延长眼轴,但对于自身对照眼的屈光状态并无影响,表明SD在豚鼠离焦眼中加速近视进展,对正常屈光状态眼的屈光发育无影响,即单独SD并不能诱导豚鼠正常眼形成近视。在随后的近视相关因素研究中,我们发现SD会加快LIM形成,LIM+SD组离焦眼的近视程度大于LIM组的离焦眼。因此,我们推测单纯SD并不足以诱导豚鼠正常眼形成近视,但能增加豚鼠离焦眼对负透镜引起的光学离焦信号的敏感性,从而促进LIM形成。
DA是一种公认的屈光发育调节神经递质,与抑制近视形成密切相关[8]。体内DA由酪氨酸合成,TH和芳香族L-氨基酸脱羧酶参与催化辅助。其中TH是DA合成的限速酶,常被用作评估DA合成的重要指标。在细胞质中合成的DA被2 型囊泡单胺转运蛋白转运,通过胞吐作用从DA能神经元轴突末端释放,并与靶受体作用,随后被突触间儿茶酚-O-甲基转移酶和单胺氧化酶(Monoamine oxidase,MAO)降解为DOPAC或被突触前膜上的多巴胺转运蛋白(Dopamine transporter,DAT)再摄取,通过2 型囊泡单胺转运蛋白转运进入囊泡中以重复利用[9]。DOPAC是DA的主要代谢产物,DOPAC/DA比值常作为评估DA分解代谢的敏感指标,DOPAC/DA增加表示该组织区域DA能神经元分解代谢旺盛、活性增强。在形觉剥夺性近视研究中发现,视网膜DA及DOPAC水平减低[10-11],外源性予以DA或激动/拮抗DA受体可有效抑制形觉剥夺性近视形成[12-14];而减少小鼠视网膜内DA含量可使正常视觉发育小鼠产生近视屈光[15]。LIM中同样如此,离焦眼视网膜DA含量显著下降[16],Thomson等[17]发现无论是给予玻璃体腔注射还是局部滴眼给药,左旋多巴均可明显延缓眼轴生长并呈现剂量依赖性显著抑制LIM发展。
本研究中我们发现LIM组和LIM+SD组豚鼠视网膜DA含量均低于自身对照眼,且LIM+SD组视网膜DA含量较LIM组更低,离焦眼视网膜DA含量与屈光度、眼轴长度均具有相关性,表明视网膜DA含量降低不但参与豚鼠LIM形成过程,还在SD促进豚鼠LIM形成中发挥作用。我们进一步检测了视网膜TH蛋白和DOPAC含量的变化,观察豚鼠视网膜DA合成及分解代谢的变化,分析豚鼠LIM视网膜DA含量变化的原因,发现LIM组和LIM+SD组之间离焦眼视网膜TH蛋白表达无显著性差异,但LIM+SD组DOPAC/DA比值高于LIM组,表明SD对视网膜DA的合成代谢无明显影响,但能引起视网膜DA分解代谢加快,是SD引起豚鼠LIM视网膜DA含量进一步下降的原因,促进LIM的进展。
但是SD引起视网膜DA含量下降,继而促进豚鼠LIM形成及进展的具体调控机制还需进一步研究。一方面可能与下丘脑-垂体-肾上腺轴激活和各种神经递质系统[18]相互作用有关,如SD可改变去甲肾上腺素能、5-HT能传递等,增加神经元放电率,增加DA传递[19-20];另一方面,DAT定位于多巴胺能神经元突触前膜,参与维持神经元内外DA稳态,也参与了睡眠-觉醒稳态调节,SD可显著降低DAT可用性[21-22],另外SD已被证明可降低纹状体D2受体活性,减弱对DAT转运功能的促进作用[23-24],其对视网膜多巴胺受体活性调节作用还有待研究;MAO是单胺类代谢的关键酶,DA主要由MAO转化DOPAC,研究发现SD还可影响MAO活性,提高对DA激动剂敏感性,促进DA能神经元传递[25]。由此可见,SD可通过降低DAT功能抑制DA向神经元内转入以及改变MAO活性促进DA转换,增强DA能神经元活动。此外,SD引起的细胞氧化应激、线粒体功能障碍和细胞内Ca2+稳态失衡等也可能在其中发挥作用[26]。除DA外,近视信号通路中转化生长因子-β、类胰岛素生长因子、视黄酸等[27]也扮演着重要角色,其与SD在近视的发生发展的相关性还有待进一步关注和研究。
关于SD和近视进展之间存在复杂关系的证据越来越多,本研究强调了SD对近视形成及进展的促进作用,可能通过提高DA代谢水平,增强DA能神经传递降低DA含量,促进近视进展。本研究中存在以下几点不足:第一,该项研究应用豚鼠造模,在短时间内发生近视改变,然而人的眼睛生长发育较慢,其屈光改变和视网膜神经回路不尽相同,甚至更加复杂,因此还需要更多的动物实验模型和体外研究;第二,我们并没有检测DAT和MAO等相关因子浓度变化,尚需进一步研究昼夜节律在近视中的作用机制及信号通路,有助于阐明以上观察到的SD、近视和DA之间的关联。此外,SD与视黄酸、转化生长因子-β等其他近视相关信号因子关联也将是今后关注的内容,我们将进一步探讨SD在近视进展中的作用和机制,为近视的预防和干预提供更多实验依据。
利益冲突申明本研究无任何利益冲突
作者贡献声明杨雪莉:收集数据,参与选题设计、资料分析及解释,撰写论文,根据编辑部的修改意见进行修改。陈尧:参与选题设计和修改论文的结果结论。关宇欣、袁超群:参与收集数据,修改论文并参与编辑部修改意见的修改。毛俊峰:参与选题设计、资料分析解释,修改论文中关键性结果、结论
