不同含油量花生品种中二酰甘油酰基转移酶基因的表达分析
2023-07-17潘丽娟许静王秀贞姜骁陈娜王通殷祥贞杨伟强迟晓元
潘丽娟,许静,王秀贞,姜骁,陈娜,王通,殷祥贞,杨伟强,迟晓元
(山东省花生研究所,山东青岛 266100)
在油料作物中,油脂主要以三酰甘油(triacylgycerol,TAG)的形式贮存在种子中。 二酰甘油酰基转移酶(diacylglycerol acyltransferase,DGAT)是催化二酰甘油合成TAG 的关键限速酶[1,2]。 目前报道的DGAT 存在四种类型——DGAT1、DGAT2、DGAT3 和DGAT/WS,彼此之间基因序列同源性很低,具有不同的亚细胞定位和表达模式[3-5],但都具有催化在二酰甘油加上酰基脂肪酸生成甘油三酯的作用[6-8]。
过表达DGAT1基因可促进种子中油脂的积累。 拟南芥DGAT1在烟草中过表达可使转基因烟草种子中TAG 含量显著上升[9]。 拟南芥突变体AS11的种子中DGAT1 活性降低,TAG 含量降低,脂肪酸组成发生变化,种子皱缩,发育迟缓,平均重量降低[10];而在野生型拟南芥中过量表达AtDGAT1后,种子含油量和千粒重得到明显提高[11]。 过量表达高油玉米DGAT1的转基因玉米种子含油量和油酸含量分别提高26.1% 和84.5%[12]。DGAT2在油料作物中过量表达,也能调控种子油脂的含量和组成[13,14],如蓖麻DGAT2过表达可促进蓖麻油中蓖麻油酸含量的增加[15]。DGAT3 和DGAT/WS 发现较晚,目前研究仅限于少数植物或微生物中,DGAT3 在TAG 合成中的功能已经在花生和产油酵母(oleaginous yeast)中被验证[16]。
在花生中,Saha 等(2006)从未成熟种子的子叶中首先克隆到AhDGAT3基因[8]; 王龙龙(2010)从花生中克隆获得了AhDGAT2的全长序列,在叶和花中表达量都较高[17];Chi 等(2014)通过构建的花生种子全长cDNA 文库,首次克隆获得AhDGAT1基因[18]。 目前花生中发现的这三种酶,AhDGAT1 和AhDGAT2 均有跨膜结构,属于膜蛋白,而AhDGAT3 为可溶性蛋白;基因序列同源性很低;AhDGAT1和AhDGAT2在种子发育前中期表达量较高,而AhDGAT3则主要在种子发育后期表达[18]。 说明它们在种子生长发育、储藏物质积累等方面所起的作用并不相同,可能在特定组织或发育阶段由主效酶发挥作用[7,17]。
花生是我国重要的油料作物,近年来我国花生总产中约52%用于榨油,年产花生油近300 万吨,占国产植物油产量的25%以上,是国产植物油的第二大来源[19]。 目前提高花生品种的含油量及改善其脂肪酸组成已成为育种专家努力的方向。 随着分子生物学和基因工程技术的发展,对花生油脂合成代谢途径及关键酶基因的认识也在不断深入。 DGAT 作为Kennedy 途径中唯一的限速酶[4],深入研究其在TAG 合成和积累过程中的作用对于阐明花生油脂及其脂肪酸组成的形成机理具有重要意义。 本研究分析了前期从花生中分离的3 个DGAT 基因AhDGAT1-1、AhDGAT1-2和AhDGAT3-3在含油量和油酸含量不同的3 个花生品种中的表达情况,以期为深入探究花生油脂合成和培育高油、高油酸新品种提供理论依据。
1 材料与方法
1.1 试验材料
试验所用3 个花生栽培品种分别为低油低油酸品种花育17 号、高油高油酸品种花育910 和高油低油酸品种花育918(表1),均为本课题组培育。 2020 年5 月初种植于山东省花生研究所莱西试验站大田,田间水肥管理等同于常规大田,及时进行病虫害防治。
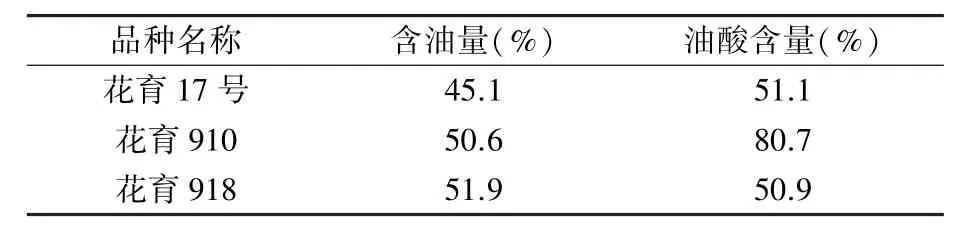
表1 供试花生品种油脂和油酸含量
1.2 取样方法
花生开花下针后,分别采集下针后30、40、50、60 d 的荚果,剥取籽仁(图1),用铝箔纸包裹,经液氮速冻后立即放入-80℃冰箱保存备用。

图1 供试材料4 个发育时期的籽仁
1.3 RNA 提取与cDNA 合成
采用北京天根生化科技有限公司试剂盒(RNeasy Mini Kit)提取样品总RNA,详细方法参考使用说明。 用DNaseⅠ处理提取的RNA 以去除DNA 污染。 利用Promega 的M-MLV 反转录酶进行cDNA 合成,每25 μL 反应体系中加入2 μg RNA;反转录体系于42℃反应1 h,之后置于冰上冷却5 min,然后将反转录产物放于-20℃备用。
1.4 实时荧光定量PCR 分析
用SYBR Green PreMix 试剂盒(购自MDBio台湾生工)按使用说明进行。 先将反转录所得的cDNA 稀释到8 ng/μL,每反应体系中加2 μL 稀释后的cDNA。 荧光定量PCR 仪采用Roche 的LightCycler 2.0。 反应程序:95℃10 s;95℃5 s,60℃30 s,72℃10 s,40 个循环;绘制熔解曲线,温度每10 s 升高0. 5℃。 所用内参基因为Actin11,所用引物见表2。每个样品设置3 次生物学重复。 采用delta-delta Cp 方法分析数据,误差线为3 次重复的标准偏差。
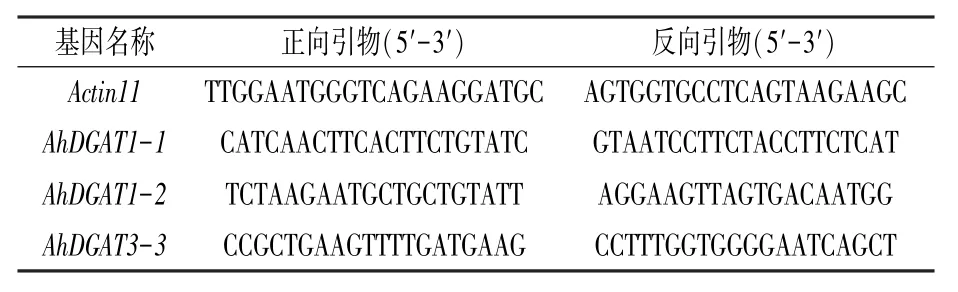
表2 荧光定量PCR 所用引物
2 结果与分析
2.1 AhDGAT1-1 基因的表达分析
随着荚果发育,AhDGAT1-1基因在3 个花生品种籽仁中的表达模式各有不同(图2)。 花育17 号与花育910 的AhDGAT1-1基因表达量均在下针后30 天(荚果发育初期)的籽仁中最高,在下针后40 天的籽仁中最低,下针后50 天又有小幅回升。 花育918 中,AhDGAT1-1基因表达量随荚果发育显著升高,在荚果发育中期(下针后40天)的籽仁中表达量最高,之后表达量逐渐降低。
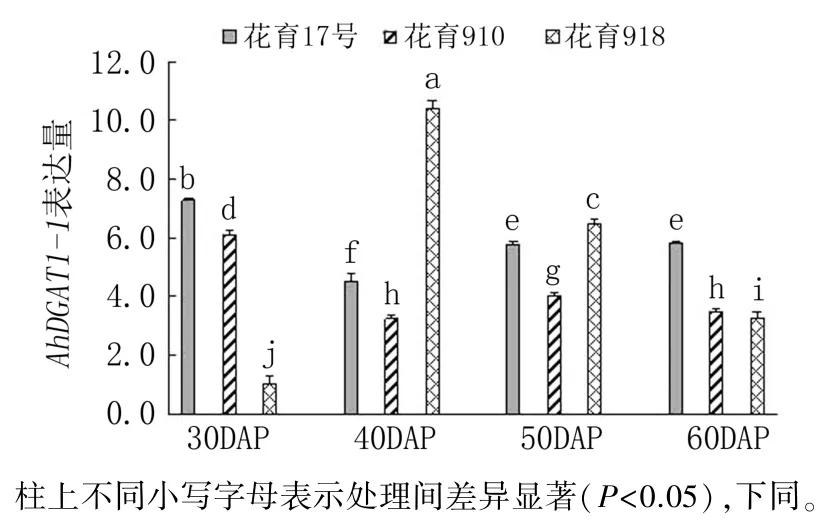
图2 花生品种间AhDGAT1-1 基因表达差异分析
AhDGAT1-1基因表达量在下针后30 天和60天表现为花育17 号>花育910>花育918,而下针后40 天和50 天则表现为花育918>花育17 号>花育910。AhDGAT1-1在花育918 下针后40 天的籽仁中表达量最高,显著高于其他时期各品种中的表达量;在花育17 号和花育910 下针后30天的籽仁中表达量也较高,与花育918 下针后50天的表达量相差较小。 说明AhDGAT1-1可能主要在低油低油酸品种和高油高油酸品种的荚果发育初期起作用,但在高油低油酸品种中则主要在籽仁油脂积累快速期。
2.2 花生AhDGAT1-2 基因的表达分析
由图3 可见,随着荚果发育,花育17 号中Ah-DGAT1-2基因的表达量呈降低-升高-降低的变化趋势,最高值出现在下针后30 天。 花育910 中AhDGAT1-2基因的表达峰值出现在下针后50天,其次为下针后30 天,下针后40 天的表达量最低。 花育918 中AhDGAT1-2基因的表达量随荚果发育先逐渐升高,下针后50 天达到峰值,之后急剧下降。 3 个品种间,下针后30 天和60 天,Ah-DGAT1-2基因的表达量均表现为花育17 号>花育910>花育918,品种间差异显著;下针后40 天和50 天,AhDGAT1-2基因均在花育17 号中的表达量最高,其次为花育918,花育910 中的表达量最低,品种间差异也达显著水平。 综合来看,Ah-DGAT1-2基因的表达峰值出现在低油低油酸品种的籽仁充实初期,而出现在高油高油酸品种和高油低油酸品种的籽仁油脂积累快速期。
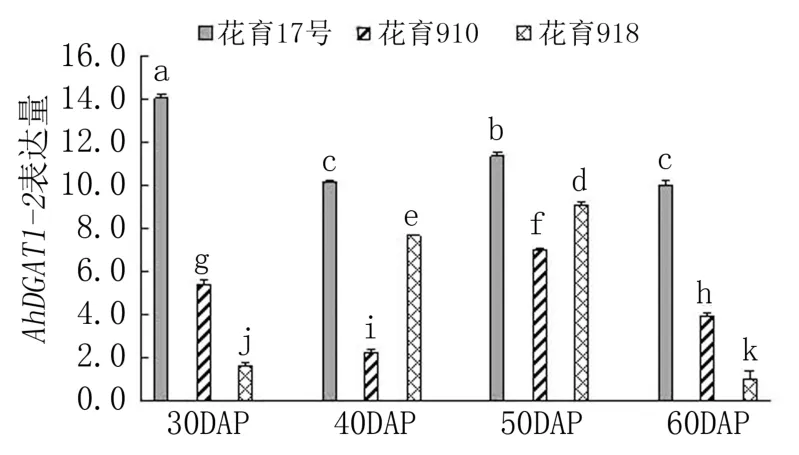
图3 花生品种间AhDGAT1-2 基因表达差异分析
2.3 花生AhDGAT3-3 基因的表达分析
由图4 可见,下针后30 天,AhDGAT3-3基因在花育910 中表达量最高,依次高于花育17 号和花育918;下针后40 天和50 天,花育17 号中Ah-DGAT3-3基因的表达量最高,花育918 中最低;下针后60 天,花育918 中AhDGAT3-3基因的表达量急剧升高,分别是花育17 号和花育910 表达量的4.65 倍和7.56 倍。 随着荚果发育,AhDGAT3-3基因表达量在花育17 号和花育910 中均呈先升高后降低趋势,均在下针后50 天达到峰值;而在花育918 中则呈持续升高趋势,在下针后60 天达到峰值。 可见,AhDGAT3-3基因主要在3 个花生品种荚果发育中后期表达,说明其可能在种子储藏物质积累方面起作用。
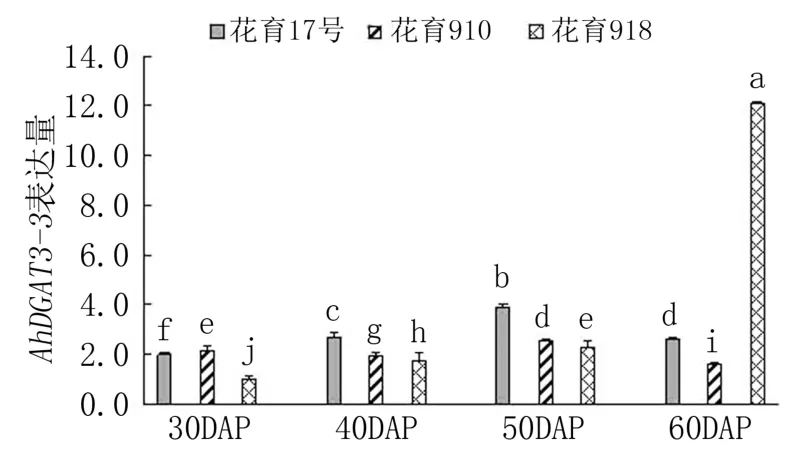
图4 花生品种间AhDGAT3-3 基因表达差异分析
3 讨论与结论
花生是我国重要的油料作物与经济作物,提高含油量是花生品种改良的主攻方向之一。 由于花生基因组测序研究起步较晚,目前从分子水平研究花生油脂合成代谢机制的还较少。 越来越多的研究表明,DGAT 在真核生物TAG 生物合成的最后一步和限速步骤中起着至关重要的作用[20-22]。 对DGAT基因的研究近几年来在一些油料作物如高油玉米、大豆、油菜、油桐(Vernicial fordii)和向日葵(Helianthus annuus) 中相继开展[12,23-26]。
本研究对前期从花生中分离的3 个DGAT基因(AhDGAT1-1、AhDGAT1-2和AhDGAT3-3)在不同油脂含量的花生品种间的表达差异进行比较,结果表明,AhDGAT1-1基因表达峰值出现在低油低油酸品种和高油高油酸品种的荚果发育初期(下针后30 天),而出现在高油低油酸品种荚果发育中期(下针后40 天);AhDGAT1-2基因的表达峰值出现在低油低油酸品种的荚果发育初期及在高油高油酸品种和高油低油酸品种的籽仁油脂积累快速期;AhDGAT3-3基因则主要在3 个品种的荚果发育中后期表达。
研究发现不同植物的DGAT 对酰基脂肪酸底物选择存在偏好性差异,致使不同物种的油脂成分不同,使特定脂肪酸在某些植物中富集,例如亚麻DGAT1 对亚麻酰CoA、油桐DGAT2 对桐酰CoA、可可(Theobroma cacao)DGAT1 对硬脂酰CoA 和花生DGAT3 对油酰基CoA(18 ∶1 CoA)的优先选择[8,27-29]。 本研究结果显示,在高油高油酸品种花育910 中AhDGAT1-1基因表达峰值出现在荚果发育初期,对促进花生籽仁油脂的合成有重要作用,但之后随荚果发育表达量降低,这种降低是否与油酸含量升高有关,还需进一步的试验验证。AhDGAT1-2基因的表达峰值出现在低油低油酸品种花育17 号籽仁充实初期,而在高油高油酸品种花育910 和高油低油酸品种花育918的籽仁油脂积累快速期,说明该基因可能在种子发育中后期参与油脂与脂肪酸的合成与积累。DGAT3是一类组成型表达基因,于种子中优势表达,在拟南芥、大豆、花生种子发育中期开始大量积累,一直延续到接近成熟阶段[3,30]。 在本研究检测的3 个花生品种中,AhDGAT3-3基因的表达峰值出现在荚果发育的中后期,也再次验证了前人的试验结果,说明该基因可能在种子储藏物质积累方面起作用。
花生油脂合成途径中关键酶基因的筛选及功能研究,对利用分子育种手段提高花生油脂含量意义重大。 本研究通过实时荧光定量PCR 检测了3 个DGAT基因在高、低油花生品种中的表达,发现AhDGAT1-1主要在荚果发育早期起作用,而AhDGAT1-2和AhDGAT3-3主要在荚果发育中后期起作用。 本研究结果对深入理解花生油脂合成积累机制和调控网络及选育高油花生品种具有一定的参考价值。
