三叶青两个受体蛋白基因的克隆及功能分析
2023-07-17林国卫武思梦张悦蔡红闻静
林国卫,武思梦,张悦,蔡红,闻静
(1. 上饶师范学院生命科学学院,江西上饶 334000;2. 玉山县红日农林农民专业合作社,江西玉山 334700)
三叶青(Tetrastigma hemsleyanuDiels et Gilg.)在我国有着悠久的民间药用历史,块根是其重要的药用部位,富含黄酮类[1]、多糖类[2]、酚酸类[3]及脂肪酸类[4]等多种次生代谢物质,其提取物具有消炎、抗病毒、解热及抗肿瘤等功效[5,6]。 由于三叶青在自然环境中生长缓慢,仅靠自然繁殖已不能满足市场需求,开发快繁及人工栽培技术对于提高产量、保护种质资源及促进其生物学研究具有重要意义。 但三叶青组织培养中存在严重的再分化障碍,通过愈伤组织途径很难形成再生芽或胚状体,目前只能通过芽原基无菌培养增殖实现快速繁殖[7,8],这严重制约了转基因技术在三叶青分子生物学基础研究中的应用。 另外,三叶青块根从纤维根到营养储藏器官的转变过程也涉及到体细胞的再分化。 研究表明胚状体受体类激酶(somatic embryogenesis receptor kinases,SERKs)是参与植物营养储藏器官形成的关键受体类蛋白[9],在体细胞胚发生及体细胞的脱分化中发挥重要作用[10,11]。 因此,本研究从三叶青转录组数据库中筛选到与SERKs 相似度最高的两个受体蛋白基因ThRLP1及ThRLK1,通过生物信息学分析、原核表达分析、块根发育不同时期及器官特异性表达分析等,初步明确其分子结构特征及表达特征,为进一步研究该类基因所涉及的生物学过程及其功能鉴定提供分子生物学依据。
1 材料与方法
1.1 试验材料
1.1.1 植物材料 本研究所用三叶青为栽培品种“怀玉2 号”,由玉山县红日农林农民专业合作社提供。
1.1.2 试剂 针对富含多酚多糖植物组织的RNA 提取试剂盒Quik RNA Isolation Kit GK,购自北京华越洋生物科技有限公司;反转录试剂盒(TransScript®One-Step gDNA Removal and cDNA Synthesis SuperMix)、基因克隆PCR 酶(2×TransTaq High Fidelity PCR SuperMixⅠ)及检测PCRTaq酶(2×EasyTaq®PCR SuperMix),均购自北京TRANSGENE 科技有限公司; 胶回收试剂盒(MiniBEST Agarose Gel DNA Extraction Kit Ver.4.0)、克隆载体(pMDTM18-T Vector Cloning Kit)及大肠杆菌DH5α 感受态,选用大连TAKARA 公司的产品;琼脂糖、β-巯基乙醇、NaCl、Tris、EDTA等化学试剂采购于百化商城。
1.1.3 试验仪器 MC nexus PCR 仪(德国,Eppendorf),实时荧光定量PCR 检测仪器(西安,天隆),DYY-6E 型电泳仪(北京,六一),Tanon 3500R 全自动数码凝胶图像分析系统(上海,天能),微量蛋白测定仪(日本,Shimadzu),高速离心机(德国,Sigma 2K15),JY96-IIN 型超声波细胞破碎仪。 由上海尼桑生物工程股份有限公司提供测序服务。
1.2 试验方法
1.2.1 样品采集与处理方法 2022 年5 月15 日上午从怀玉山红日农林农民专业合作社的温室中(E117°58′17″,N28°53′51″)采集三叶青的健康功能叶、匍匐状分枝(枝蔓)、膨大块根、受精子房及顶芽,然后取叶片正中间1/4 大小作为叶片样品,取两片叶子中间的枝截取1/4 长度作为枝蔓样品,取块根中间部位约1/4 大小作为块根样品;受精子房取样以子房膨大及花冠萎蔫为标准,取样时去掉花托及萎蔫的花冠部分;顶芽取样尽量去掉周围的幼叶。 每种样品取3 ~5 株,剪碎后混合,称取100 mg,重复3 次。
不同发育时期块根采样参照闻静等[12]的方法,样品称量后立即液氮速冻,于-80℃保存备用。
1.2.2 总RNA 的提取及cDNA 的合成 以顶芽为样本,液氮研磨后提取总RNA,操作步骤参照RNA 提取试剂盒说明书。 对提取后的RNA 进行纯度及定量分析,然后用70 ng/μL 的总RNA 作为样本进行反转录合成cDNA,操作步骤参照反转录试剂盒说明书。
1.2.3ThRLP1和ThRLK1基因的克隆和测序根据前期从三叶青块根及叶片转录组获得的基因序列及注释信息分析,设计ThRLP1及ThRLK1全基因扩增引物、开放读码框扩增引物及荧光定量引物(表1)。 PCR 反应体系为50 μL,反应成分及用量参照试剂盒说明书;反应程序为98℃变性5 s,56℃退火5 s,72℃延伸1.5 min,共35 个循环。 PCR 产物用1.2%~1.5%琼脂糖TAE 凝胶电泳检测,纯化后与克隆载体连接,转化大肠杆菌,菌落PCR 检测后测序。

表1 用于ThRLP1 和ThRLK1 基因克隆及荧光定量分析的引物
1.2.4ThRLP1和ThRLK1基因的生物信息学分析 采用在线软件进行相关分析,具体见表2。采用MEGA 7 进行多重比对后利用neighbor-joining 算法构建进化树。

表2 生物信息学分析在线软件名称、用途及网址
1.2.5ThRLP1和ThRLK1基因表达的RT-qPCR分析 相对荧光定量采用染料法,反应不同组分用量及反应温度条件参照荧光定量试剂盒说明书两步法操作,反应体系为20 μL。 器官特异性表达检测中采用ELF-α(MT731970)[12]为内参基因,块根发育不同时期的引物设计参考闻静等[12]的方法,内参基因为RPⅡ(MW307822)。 试验设置生物学重复、技术重复各3 次。 数据分析采用2-△△Ct法[13],差异显著性分析采用SPSS 软件的LSD 法。
1.2.6ThRLP1和ThRLK1基因的原核表达分析以ThRLP1及ThRLK1基因的开放读码框序列设计引物并进行PCR 扩增。 扩增片段回收后与原核表达载体连接,具体连接及转化方法参考pEASY®-Blunt 载体说明书;转入DH5α 抗生素筛选,经扩大培养后提取质粒测序,选择正确的重组原核表达载体序列转化DE3 感受态,涂布在含有50 μg/mL 氨苄西林的固体LB 培养基上培养12 h;挑取单菌落扩大培养,菌液浓度达到OD600nm为0.45~0.60 时加入1 mmol/L IPTG 诱导蛋白表达,摇菌4 h 后取1~5 mL 菌液,4℃、12 000 r/min离心,收集菌体沉淀后冰上超声波破碎,4℃、12 000 r/min 离心20 min,取上清进行SDSPAGE 检测。
2 结果与分析
2.1 目的基因克隆
以怀玉山三叶青顶芽的cDNA 为模板扩增ThRLP1和ThRLK1基因的全长序列,凝胶电泳结果(图1)显示,扩增产物大小与预期接近。 经测序得到的碱基序列与前期获得的转录组序列相似度大于99%,推测的氨基酸序列与由转录组原序列推测的氨基酸序列相似度为100%。
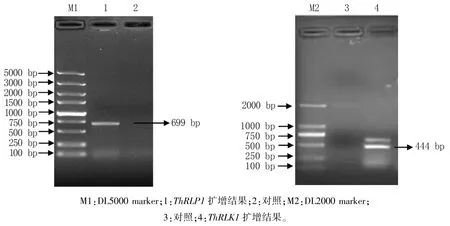
图1 ThRLP1 和ThRLK1 基因PCR 电泳检测结果
2.2 ThRLP1 和ThRLK1 基因的生物信息学分析
2.2.1ThRLP1和ThRLK1基因推测蛋白质序列的生物信息学分析ThRLP1基因开放读码框长度为657 bp,预测编码218 个氨基酸。 经在线比对,蛋白序列与河岸葡萄(Vitis riparia)的富含亮氨酸的重复蛋白亚型1(leucine-rich repeat protein 1-like isoform X1,XP_034694208.1)相似度最高,为94.50%,序列覆盖率为100%。 特异匹配保守域分析结果(图2)表明在第26 ~67 个氨基酸位点有N 端的亮氨酸重复序列(leucine rich repeat N-terminal domain,LRRNT_2,pfam08263),在第95~154 个氨基酸位点有亮氨酸富含区(leucine rich repeat,LRR_8,pfam13855)。

图2 ThRLP1 基因推测氨基酸序列的信号肽、保守氨基酸及保守域分析
ThRLK1基因开放读码框长度为399 bp,预测编码132 个氨基酸;蛋白质序列与葡萄的SERK2(XP_002276414.2)相似度最高,为96.97%,序列覆盖率为100%;保守域分析结果(图3)表明在第1~85 氨基酸位点有丝氨酸/苏氨酸激酶的催化结构域(catalytic domain of the serine/threonine kinase,STK_BAK1_like)。
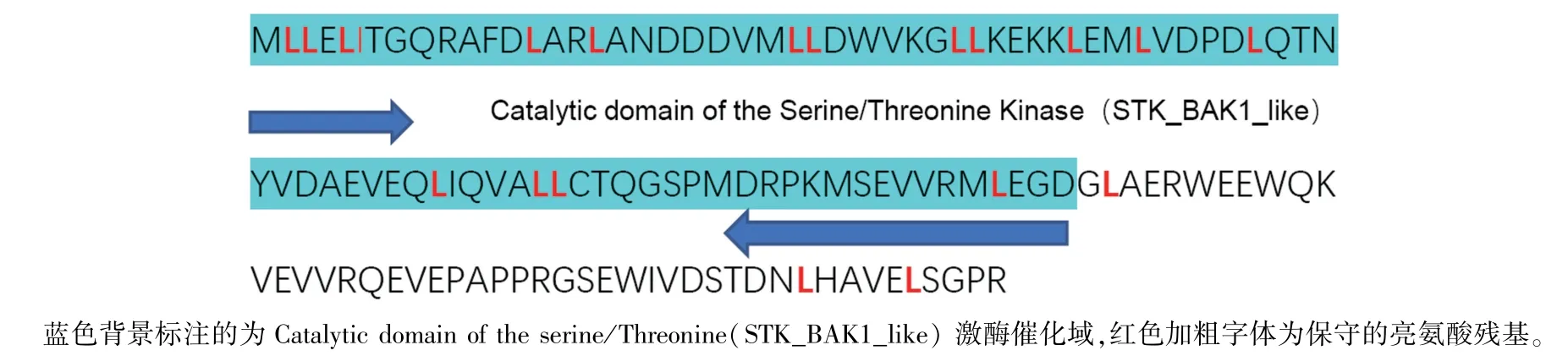
图3 ThRLK1 基因推测氨基酸序列的保守氨基酸及保守域分析
ThRLP1和ThRLK1基因预测蛋白的分子量、等电点、分子功能及亚细胞定位等见表3。

表3 ThRLP1 和ThRLK1 蛋白的生物信息学分析结果
2.2.2ThRLP1和ThRLK1基因的进化分析 利用NCBI 数据库的Blastp 比对,将ThRLP1 和ThRLK1及与其同源性较高的46 条蛋白通过neighbor-joining 构建系统进化树。 由图4 可见,所有蛋白被分为3 类,ThRLP1 和ThRLK1 分处于不同的类群,其中,ThRLP1 与葡萄的VvSERK1 和河岸葡萄的VrLRP1处于同一进化分枝,序列相似性最高;ThRLK1 与葡萄的VvSERK2 和河岸葡萄的VrBAK1 处于同一进化分枝,在系统发育中序列相似性最高。
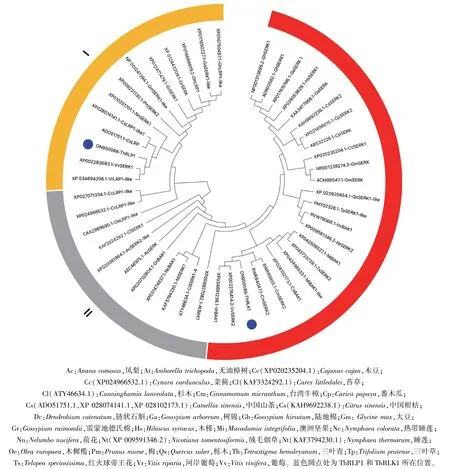
图4 ThRLP1 和ThRLK1 及其同源蛋白序列的系统进化树
2.3 ThRLP1 和ThRLK1 基因的表达特征分析
2.3.1 两基因在三叶青不同器官中的表达ThRLP1在三叶青块根中的表达量显著低于其他器官,而叶片、顶芽、受精的子房及枝蔓中该基因的表达差异不显著(图5A)。 在5 种器官中都检测到了ThRLK1基因的表达,受精的子房中表达量最低,显著低于其他器官;顶芽中的表达量略高于受精的子房,但明显低于叶片、块根中的表达水平;该基因在枝蔓中的表达量最高,显著高于其他4 种器官(图5B)。
2.3.2 两基因在块根发育不同时期的表达分析
由图6 可见,ThRLP1在具有块根的纤维根(Ⅳ)中表达量最高,且与其他发育状态的根中的表达量差异显著;其次为在纤维根期(Ⅰ)与块根期(Ⅲ),两者间差异不显著,但显著高于块根的初始膨大期(Ⅱ)。ThRLK1基因在具有块根的纤维根(Ⅳ)中表达量最高,显著高于纤维根期(Ⅰ)和块根初始膨大期(Ⅱ),但与块根期(Ⅲ)差异不显著;在纤维根期(Ⅰ)该基因的表达量最低,显著低于块根期(Ⅲ)与具有块根的纤维根(Ⅳ),但与块根初始膨大期(Ⅱ)的表达量差异不显著。

图6 三叶青块根发育不同时期ThRLP1(A)和ThRLK1(B)基因的相对表达量
2.4 ThRLP1 和ThRLK1 的原核表达分析
原核诱导表达分析结果(图7)显示,含有重组表达载体pEASY- Blunt E1ThRLP1与pEASYBlunt E1ThRLK1的受体菌经过4 h 的ITPG 诱导后,分别诱导表达出29 kD 与20 kD 左右的蛋白,与预测的目的蛋白大小相符。
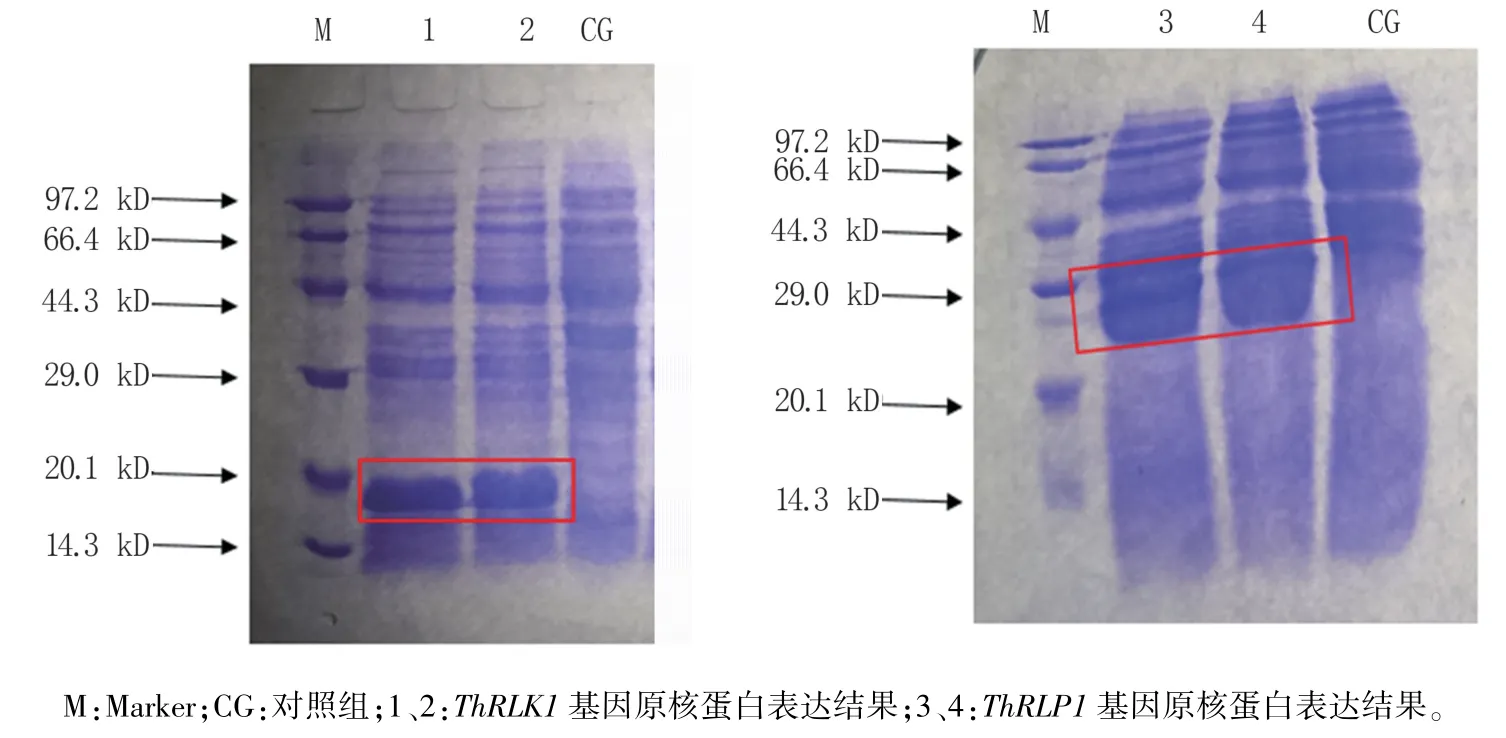
图7 ThRLK1(A)及ThRLP1(B)的原核表达SDS-PAGE 电泳结果
3 讨论与结论
SERK 类蛋白已经被证明在植物愈伤组织向体细胞胚发育过程中起到重要作用,如在唇兰的组织培养中,在体细胞胚发育初期,ClSERK基因在由原球茎产生的胚性愈伤组织中高度表达[14];Pérez-Núñez 等证实CnSERK表达与体细胞胚胎发生的诱导有关,是调控体外培养的椰子组织中形成体细胞胚胎的关键基因[15]。 典型的SERK类蛋白包含一个富含丝氨酸或脯氨酸的胞外结构域、一个跨膜结构域和一个胞内激酶结构域[16,17]。 本研究结果表明ThRLP1基因所编码的氨基酸序列富含亮氨酸残基,具有类似LxxLxx-LxxLxLxN 的保守结构域及细胞外LRR 蛋白的特征结构基元Lt/sgxIP;该蛋白序列具有信号肽序列,但缺少质膜内的激酶区域,所以ThRLP1 是一类细胞外受体蛋白[18]。 ThRLK1 不具有信号肽序列,缺少细胞外区域及跨膜区的保守域,但具有激酶特征的丝氨酸/苏氨酸激酶的催化结构域,因此被称为质膜受体类激酶[19]。 与已知胚状体受体类激酶分子结构对比, ThRLP1 和ThRLK1 的蛋白序列只具有部分胚状体受体类激酶保守域,通过进一步对三叶青块根及叶片转录组中序列的筛选比对,并没有发现同时具有细胞外LRR 氨基酸结构域及质膜受体结构域的SERK 类蛋白。 另外,ThRLP1具有SERK 的胞外结构及跨膜结构,而ThRLK1具有SERK 的膜内激酶结构,项目组初期怀疑两个基因可能来源于一个完整的SERK类基因片段,但是通过设计特异性引物扩增,并没有得到预想的结构完整的SERK 类蛋白序列。 而与ThRLP1 和ThRLK1 蛋白序列相似性较高的葡萄中确实存在具有类似结构的同类蛋白质,因此本研究克隆的ThRLP1和ThRLK1基因可以作为进一步研究三叶青SERK 类蛋白质的对象。
本研究在器官特异性表达分析中发现ThRLP1在三叶青叶片、顶芽、受精后的子房、块根及枝蔓中均有表达,与Sultana 等[20]发现拟南芥的细胞外LRR 类受体蛋白在器官中的表达较为普遍的结果类似;但块根中的表达量显著低于其他器官,推测该基因与块根发育呈负相关性或相关性较小。 目前受体类蛋白参与根或地下储藏器官发育的研究并不多见,Chen 等[21]研究发现富含亮氨酸受体蛋白(leucine-rich repeat RKs,LRRRKs)可以直接与根部顶端生长因子RGF(root meristem growth factor)作用,控制根顶端分生组织的发育。 而甜薯贮藏根组织mRNA 印迹分析表明,其受体类激酶SRF6 的mRNA 定位于木质部的初生形成层和分生组织周围,控制贮藏根增厚[22]。 但细胞外受体类蛋白直接参与植物地下储藏器官形成的报道还未见,目前植物胞外受体蛋白的研究主要集中在植物与病原互作的免疫反应过程[23,24]。
ThRLK1在三叶青各器官中也均有表达,且在顶芽及受精的子房中表达量较低,推测该基因与芽的发生过程相关性较小。 三叶青匍匐状分枝是其主要的运输器官,主要分布着疏导组织,Zhang等研究表明拟南芥的SERK家族受体类激酶与CLE41/TDIF-PXY 作为共同的受体调节植物维管束的发育[25],而ThRLK1在三叶青枝蔓中的表达量最高,初步推测其可能与维管组织发育相关。Blümke 等研究表明拟南芥质膜受体激酶MAZZA与CLV1f 共同控制的信号转导网络控制根的发育[26],但目前质膜类激酶参与到三叶青块根发育的研究还未见报道。 本研究发现,虽然ThRLK1基因在三叶青块根中的表达量低于枝蔓,但是在块根和具有块根植株的纤维根中表达量较高,推测其可能在块根发育过程中起一定作用,具体作用机制还需深入研究。
植物器官再分化及体细胞胚发育是一个复杂的生物学问题,受到植物基因型及培养环境中激素、养分等诸多因素的调节。 本研究克隆的两个三叶青受体类蛋白虽然与SERKs 蛋白序列相似度较高,但在芽与受精的子房中并无显著的高丰度表达,在块根中表达量也低于枝蔓。 因此,有关怀玉山三叶青SERK 类蛋白的鉴定及功能还有待进一步研究。
