宏病毒组测序检测豆蚜田间种群病毒种类
2023-07-08靳道然朴敬爱季英华
朴 君,靳道然,,朴敬爱,季英华,孙 枫,李 硕
(1.辽宁师范大学 生命科学学院,辽宁 大连 116081;2.江苏省农业科学院 植物保护研究所,江苏 南京 210014)
蚜虫属于半翅目(Hemiptera)蚜总科(Aphidoidea),种类繁多,已知超过4 700种,繁殖快,适应能力强,主要分布在北半球的温带和亚热带地区[1]。蚜虫的寄主范围十分广泛,寄主植物涉及267个科、2 120个属,主要包括禾本科、豆科、兰科、杨柳科等[2-3]。蚜虫是农业主要害虫之一,其为害主要包括直接为害和间接为害两方面:直接为害是指蚜虫借助口针刺吸植物嫩芽和新叶中的汁液来补充自身的营养物质,常导致植物生长缓慢;间接为害是指蚜虫传播植物病毒,蚜虫在刺探和取食植物的过程,口针中结合的或唾液腺外泌的病毒粒子会被释放进入植物体内[4],引起严重的植物病毒病害,致使农作物产量和品质降低,给农业生产造成严重的经济损失。蚜虫是最大的一类媒介昆虫,其传播的植物病毒超过虫媒植物病毒种类的二分之一[3-4]。控制蚜虫种群及其传毒危害是保障农作物安全生产的有效手段。
病毒是一种具有细胞感染性的亚显微粒子,个体微小,结构简单,只能寄生在活细胞内并依赖宿主细胞因子以复制方式增殖[5]。自然界中几乎所有的具有细胞结构的生命体都可以被病毒感染,病毒严重影响到了人类健康、社会安定以及地球生态系统的稳定[6]。在农业领域,对农业生产造成严重危害的同时,病毒也是重要的生物防治资源,其中昆虫病毒杀虫剂应用最广,具有无污染、毒力高、防效长、靶标害虫不易产生抗药性等优点[7]。在我国,棉铃虫核型多角体病毒和松毛虫质型多角体病毒是研究和应用较多的病毒,在害虫生防中发挥了重要作用[8]。目前,已鉴定出涉及15个科600多种昆虫致病性病毒[9]。蚜虫具有聚集取食和转主迁移习性,这为使用昆虫病毒对其进行生物防治提供了有利条件。
豆蚜(Aphiscraccivora),也称花生蚜,豆科作物重要害虫,在长江流域年发生20代以上,豆蚜春季在蚕豆和紫云英上繁殖,5月中下旬以后,产生有翅蚜迁向刀豆、豇豆、花生等豆科植物上繁殖危害,11月以后迁向蚕豆、冬豌豆等心叶或叶背处繁殖越冬。目前,对豆蚜自然种群携带的植物和昆虫病毒种类尚缺乏系统认识,为此,本研究通过宏病毒组测序对江苏豆蚜田间种群所携带的病毒种类进行分析鉴定,不但可以掌握豆蚜传播植物病毒的种类,利于植物病毒病防控,而且可以挖掘其体内昆虫病毒资源、发现新病毒,为利用昆虫病毒进行蚜虫生防提供参考。
1 材料和方法
1.1 试验材料
供试虫源:蚜虫自然种群于2021年12月采自江苏省南京市江宁区3块蚕豆田,带回实验室后,根据相关文献[10]进行形态鉴定,初步判定为豆蚜,蚜虫置于-70 ℃保存。
主要试剂:TRIzol试剂购自Invitrogen公司;NEBNext Ultra Ⅱ RNA文库制备试剂盒购自NEB公司;高保真DNA聚合酶、pMD18-T载体和反转录试剂盒(PrimeScript 1st Strand cDNA Synthesis Kit)购自TaKaRa公司。
1.2 试验方法
1.2.1 蚜虫种类的分子鉴定 5头蚜虫为1组,设5组生物学重复,采用CTAB法提取蚜虫总DNA。使用蚜虫近缘种鉴定通用引物[11](LCO1490:5′-TCAACAAATCATAAAGATATTGG-3′,HCO2198:5′-TAAACTTCAGGCTGACCAAAAAATCA-3′)PCR扩增蚜虫COI基因。分离纯化扩增产物,连接至pMD18-T载体,经转化大肠杆菌,选取阳性克隆测序。利用Blast(http://www.ncbi.nlm.nih.gov/BLAST/Blast.cgi)比对测序结果,明确蚜虫种类。
1.2.2 豆蚜总RNA的提取 鉴定蚜虫种类后,取豆蚜500头液氮研磨,使用TRIzol试剂提取总RNA。通过1%琼脂糖凝胶电泳和Agilent 2100 bioanalyzer检测RNA质量和完整性,NanoDrop 2000微量紫外分光光度计测定RNA纯度(OD260/280及OD260/230比值),Qubit2.0 Fluorometer测定RNA浓度。
1.2.3 文库构建和测序 RNA质控合格后,使用NEBNext Ultra Ⅱ RNA文库制备试剂盒构建文库。将总RNA除去核糖体RNA得到mRNA,随后在NEB Fragmentation Buffer中用二价阳离子将mRNA随机打断成短片段(250~300 bp),随机引物将短片段反转录为第1链cDNA,PCR扩增cDNA,即得到测序文库。文库质检合格后,进行Illumina测序,由测序公司完成。
1.2.4 数据处理与病毒注释 过滤测序原始数据,去除带接头、含N(无法确定碱基信息)、低质量(Qphred≤20的碱基数占整个read长度50%以上)reads,得到高质量Clean reads,然后将其与蚜虫基因组比对去除宿主的reads。获得无宿主污染的数据后,使用Trinity软件[12]进行转录本拼接,再利用DIAMOND软件[13]将拼接结果与GenBank Virus RefSeq、NR和CDD数据库比对,获得候选序列。对病毒序列进行ORF预测、物种注释、基因功能注释,评估其有效性,鉴定出病毒种类。
1.2.5 新病毒分子检测和序列分析 在豆蚜体内发现1种与细胞质弹状病毒属(Cytorhabdovirus)同源性较高的病毒,推测是一种新的植物病毒,对其进行基因检测。根据Contigs序列,设计2对特异引物,Crhv1-F:5′-ATGAGGGGATTTACTGACCCGCCCT-3′,Crhv1-R:5′-CGGACACGCAATAGAGGGCCAT-3′,Crhv2-F:5′-GTTTTAGAAGAAATTCTTGCT-3′,Crhv2-R:5′-TTATAACATAACAAGGAAGAAC-3′。以蚜虫总RNA为模板,利用反转录试剂盒合成第1链cDNA,进行PCR扩增。产物经基因克隆和测序,获得病毒基因片段信息。使用Blast和DNAstar软件MegAlign程序进行序列比对和同源性分析,利用MEGA-X软件[14]采用最大似然法(Maximum Likelihood)完成氨基酸序列聚类分析,使用1 000次Bootstrap Replications验证进化树可信度。
此外,在豆蚜体内新发现了昆虫病毒(HimetobiPvirus,HiPV),该病毒尚未在蚜虫中报道,故对其进行核酸和蛋白检测验证。根据灰飞虱来源的HiPV结构蛋白VP1序列(GenBank登录号:AB017037)设计引物,vp1-F:5′-GCTAACTTTGCGTCTACTG-3′和vp1-R:5′-CTAAACTTTGTCAAAGGGAT-3′,对VP1基因进行RT-PCR检测和序列分析。同时,取30头蚜虫提取总蛋白,12.5%SDS-PAGE电泳后,使用实验室前期制备的VP1抗体[15]进行Western Blot分析。
2 结果与分析
2.1 豆蚜的基因鉴定
田间采集的蚜虫根据形态鉴定,初步判定为豆蚜。5头蚜虫分为1组,共5组,提取总DNA扩增COI片段,得到符合预期大小的特异条带(图1),测序显示这些片段大小为707 bp且序列一致,经BlastN比对,其核苷酸序列与豆蚜COI基因片段(AB506718.1)相似性达99.58%,确定本研究所用蚜虫为豆蚜。

M.DNA标准分子量;1—5.蚜虫样本。M.DNA Marker;1—5.Aphid samples.
2.2 宏病毒组测序获得蚜虫体内病毒序列信息
豆蚜文库经宏病毒组测序得到35 867 014条原始序列,总碱基数为5.2 G,经质控后得到34 672 404条高质量的Clean reads,拼接后获得127 863条Contigs。经GenBank、NR和CDD数据库比对和分类注释,筛选出病毒候选序列329条,ORF预测和基因注释后,共获得病毒序列177条,与24种病毒序列一致或同源性较高。
2.3 豆蚜体内病毒种类鉴定
鉴定的24种病毒中,有5种植物病毒,来自5个病毒科(表1),马铃薯Y病毒科(Potyviridae)、帚状病毒科(Virgaviridae)、泛欧尔密病毒科(Botourmiaviridae)、弹状病毒科(Rhabdoviridae)和白纤病毒科(Phenuiviridae),显示测试的豆蚜群体携带该5个病毒科的病毒。其中匹配序列最多的病毒为一种与水稻条纹花叶病毒(Ricestripemosaicvirus,RSMV)有一定同源性的病毒,说明有一种弹状病毒在豆蚜体内有较高的含量。鉴定了19种专性寄生的昆虫病毒(表2),涉及9个病毒科,属于DNA病毒的有杆状病毒科(Baculoviridae)和昆虫痘病毒亚科(Entomopoxvirinae),属于RNA病毒的有楚病毒科(Chuviridae)、黄病毒科(Flaviviridae)、双顺反子病毒科(Dicistroviridae)、复制酶置换四体病毒科(Permutotetraviridae)、Artoviridae、幻影病毒科(Phasmaviridae)以及呼肠孤病毒科(Reoviridae),此外,还包括6种暂未分类RNA病毒。这些昆虫病毒,除黄病毒科和双顺反子病毒科病毒之前已在蚜虫体内发现[16-17],其余在蚜虫中鲜有报道。

表1 蚜虫体内经宏病毒组测序鉴定的植物病毒Tab.1 Identified plant viruses in Aphis craccivora via virome sequencing
表2 蚜虫体内经宏病毒组测序鉴定的昆虫病毒Tab.2 Identified insect viruses in Aphis craccivora via virome sequencing
2.4 新植物弹状病毒的检测和序列分析
根据获得的与RSMV同源的Contigs设计引物,对豆蚜总RNA进行RT-PCR检测,检出2个符合预期大小的条带(图2)。测序显示,2个片段分别为558,460 bp,核苷酸序列经BlastN在NCBI数据库中没有比对到结果。氨基酸序列BlastX比对结果显示,其与RSMV L蛋白(RNA聚合酶)(GenBank 登录号:AZB50424.1)同源性最高,分别为51.61%,34.35%。另外,与该序列相似性较高的11种病毒分离物均来自细胞质弹状病毒属,2个片段与这11种弹状病毒的L蛋白进行氨基酸序列比对,可以观察到该序列与这些弹状病毒具有相同的保守区(图3)。这些结果说明,扩增的病毒基因片段来源于一种新的弹状病毒,将其暂命名为Aphiscraccivoraassociatedrhabdovirus(AcARV)。将获得的2个片段氨基酸序列合并,利用MEGA-X软件将AcARV与上述11种弹状病毒分离物进行系统进化分析,结果显示,这些病毒L蛋白在系统进化树中可聚类到2个大的分支中,AcARV与RSMV构成最小分支(图4),说明这2种病毒在进化上最为近缘。在病毒聚类的两大分支中,AcARV所在分支中含有9种病毒,其寄主主要为禾本科植物,暗示AcARV的植物寄主可能为禾本科作物。

A.病毒基因片段1的扩增;B.病毒基因片段2的扩增。A.PCR amplification of viral gene segment 1;B.PCR amplification of viral gene segment 2.

A.AcARV片段1与细胞质弹状病毒的比对;B.AcARV片段2的比对;比对所涉及病毒序列信息:AcARV.本研究所获序列;BARV.菜豆伴随弹状病毒(GenBank登录号:QAU20941.1);BYSMV.大麦黄条点花叶病毒(YP_009177231.1);CARV.柑橘伴随弹状病毒(QMS92530.1);CBDAV.香芋波邦内病伴随病毒(YP_009362280.1);MARV.玉米伴随弹状病毒(ARS22495.1);MSSV.玉米不育矮化病毒(QBJ27595.1);MYSV.玉米黄条纹病毒(ATN96453.1);NCMV.北方禾谷花叶病毒(NP_597914.1);PARV.番木瓜伴随弹状病毒(YP_010087157.1);RSMV.水稻条纹花叶病毒(AZB50424.1);RVR.玫瑰病毒R(QQZ02079.1)。A.Alignment among AcARV segment 1 and other Cytorhabdoviruses;B.Alignment of AcARV segment 2;Viral sequence information used in alignment is as follows:AcARV.Obtained in this study;BARV.Bean-associated rhabdovirus(GenBank No.QAU20941.1);BYSMV.Barley yellow striate mosaic virus(YP_009177231.1);CARV.Citrus-associated rhabdovirus(QMS92530.1);CBDAV.Colocasia bobone disease-associated virus (YP_009362280.1);MARV.Maize associated rhabdovirus(ARS22495.1);MSSV.Maize sterile stunt virus(QBJ27595.1);MYSV.Maize yellow striate virus(ATN96453.1);NCMV.Northern cereal mosaic virus(NP_597914.1);PARV.Papaya associated rhabdovirus(YP_010087157.1);RSMV.Rice stripe mosaic virus(AZB50424.1);RVR.Rose virus R(QQZ02079.1).

系统树所用病毒序列信息与图3一致。Viral sequence information used in phylogenetic tree is consistent with Fig.3.
2.5 豆蚜体内昆虫病毒HiPV的检测
为了检测豆蚜体内昆虫病毒HiPV的感染情况,根据HiPV灰飞虱分离物VP1基因设计引物,经RT-PCR扩增出目的片段(图5-A),测序结果显示,所扩条带为774 bp的VP1基因。Blast比对显示,其与日本灰飞虱分离物VP1序列(AB017037)核苷酸同源性为95.5%,氨基酸序列相似性为95.3%,与江苏省农科院植保所病毒实验室保存的灰飞虱群体中HiPV VP1相似度极高,核苷酸和氨基酸序列同源性分别为99.0%,98.1%,这说明不同昆虫寄主之间HiPV的相似性很高。提取豆蚜总蛋白,经12.5% SDS-PAGE电泳后,用VP1抗体进行Western Blot检测,结果显示,可以从豆蚜中特异性检测出一条约28 ku的蛋白条带(图5-B),符合预期大小,即为HiPV VP1蛋白,进一步从蛋白水平证明了豆蚜体内存在HiPV的感染,这是首次在蚜虫体内发现HiPV。
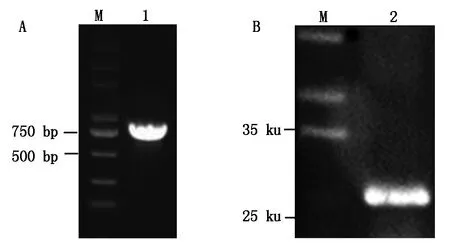
A.HiPV VP1的PCR扩增;B.HiPV VP1的Western Blot检测。A.PCR amplification of HiPV VP1 gene;B.Western Blot detection of HiPV VP1.
3 结论与讨论
蚜虫传播病毒造成的危害已经极大程度超过了蚜虫本身所带来的危害[18]。目前,对蚜虫的控制仍以化学防治为主,但大量使用杀虫剂不仅污染环境,而且还会引起蚜虫抗药性,不符合农业绿色、可持续的发展趋势。过去十多年,以天敌资源(主要是寄生蜂)为核心的蚜虫生物防治也得到了长足的发展[19],而利用病原微生物控蚜的研究与应用仍然较少。昆虫病毒是一种天然杀虫剂,可以在宿主昆虫种群中大规模传播,具有杀虫毒力高、环境污染小等优点,且不易诱导昆虫产生抗性[7]。我国在20世纪60年代初开始将昆虫病毒用于生物防治,目前,已有30多种昆虫病毒被用于农林害虫防治[20]。当前,人们仍掌握不清蚜虫群体中昆虫病毒种类,制约了利用病毒进行蚜虫生防的研究工作。为此,本研究通过宏病毒组测序对江苏豆蚜种群所携带的病毒种类进行了分析鉴定,以期在掌握其携带植物病毒的同时,挖掘其体内昆虫病毒资源。
宏病毒组,又称病毒转录组,是一种新兴的以宏转录组为理论基础的病毒分子检测技术,可直接以组织或环境样本为对象,快速鉴定样本中全部病毒种类,为病毒病快速诊断、新病毒发现和病毒流行预警提供了有力支撑。与传统病毒检测技术相比,高通量测序技术(主要包括宏病毒组和小RNA深度测序)极大提高了病毒检测的效率和准确性,具有无法比拟的优势。随着该技术不断成熟和成本降低,其已成为临床和实验室研究中的常用技术,利用该技术鉴定的新病毒种类日趋增多[21-22]。作者之前在半翅目昆虫中首次发现类复制酶置换四体病毒,便是基于高通量测序技术完成鉴定[23]。
本研究对豆蚜携带的病毒种类进行了全面分析,共鉴定出24种病毒,其中植物病毒5种,昆虫病毒19种。对植物病毒中Contigs数目最多的病毒进行基因检测和序列分析,发现其与RSMV的RNA聚合酶氨基酸序列同源性最高(51.61%),且与其他细胞质弹状病毒属成员具有共同的保守区,这提示在豆蚜体内检测到一种新的弹状病毒,暂命名为AcARV。细胞质弹状病毒属病毒的分布是世界性的,单个病毒的寄主范围较窄,但整个属的寄主范围较广。该属成员可由蚜虫、叶蝉、飞虱等媒介昆虫传播,病毒在植物和介体昆虫体内均可增殖,少数病毒也可经汁液传播。前期有研究在烟粉虱体内也鉴定出类似的弹状病毒(Bemisiatabaciassociatedvirus1)[24]。本研究发现的AcARV由蚜虫传播,但其植物寄主尚不清楚,根据目前掌握的氨基酸序列系统进化分析结果,AcARV与其他8种病毒聚类成一个大的分支,该分支中的成员其寄主要为禾本科植物,推测AcARV的寄主可能是禾本科作物。由于豆蚜主要取食危害豆科植物,因此,也不排除AcARV的植物寄主为豆科作物。当然,其真实寄主的确定还需后续对相关植物进行病毒病害调查和检测鉴定。另外,AcARV全基因组序列尚不清楚,也需进一步完成测序。
在鉴定的昆虫病毒中,HiPV较为引人注意。HiPV为双顺反子病毒科Triatovirus属成员(原为蟋蟀麻痹病毒属Cripavirus成员),是一种低致病性、可持续性感染宿主的病毒[25]。该病毒最早由日本学者在灰飞虱中发现,之前报道的HiPV寄主仅为几种稻飞虱[25],本研究证明了豆蚜体内存在HiPV的感染,这是首次在蚜虫体内发现HiPV,该病毒能否侵染其他蚜虫种类有待进一步明确。作为正单链RNA病毒,HiPV基因组结构简单[26],通过侵染性克隆“拯救”策略大量制备该病毒相对较为容易,昆虫寄主范围的扩大使其具备作为生防病毒的潜力。HiPV对稻飞虱的致病性较弱,是其用作生防病毒的不足之处,对其毒力基因进行改造增加致病力,将是决定其生防应用前景的关键。同为双顺反子病毒科的Rhopalosiphumpadivirus(RhPV)可感染蚜虫,并缩短其寿命和抑制繁殖力[17],HiPV对蚜虫的致病性程度决定着其在防治蚜虫方面的应用潜力,后续值得关注。病毒与昆虫在长期进化过程中形成了复杂多样的关系。多数昆虫病毒对寄主产生致病性不利于其生存,而少数病毒则有利于寄主昆虫的生存和扩散[27]。前期研究发现,HiPV可与灰飞虱传播的水稻条纹病毒互作并参与调控病毒增殖[28],蚜虫作为重要的媒介昆虫,HiPV在其体内是否影响蚜虫对植物病毒的传播,这也是一个令人感兴趣且有意义的研究方向。此外,还在蚜虫体内发现了复制酶置换四体病毒科病毒,之前本实验室在灰飞虱中也发现了这类病毒[23],这说明自然界中该类病毒在昆虫体内广泛存在。
本研究鉴定了江苏地区越冬豆蚜群体中携带的病毒种类,结果发现,蚜虫体内含有多种植物病毒,说明其对周边农作物有潜在的传毒威胁,因此,生产上需要加强对蚜虫传播病毒病的监测和防控。同时,发现豆蚜体内昆虫病毒资源较为丰富,证实HiPV可感染蚜虫,为今后生防研究和应用提供了候选病毒资源。
