不同抗旱基因对小麦千粒质量的影响
2023-07-08程小虎黄冀楠于翠红胡梦芸孙丽静王清涛张颖君
杨 锴,程小虎,赵 杰,黄冀楠,于翠红,张 丽,胡梦芸,孙丽静,李 辉,王清涛,张颖君
(1.河北工程大学 园林与生态工程学院,河北 邯郸 056038;2.河北省农林科学院 粮油作物研究所,河北省作物遗传育种实验室,河北 石家庄 050035;3.定西市农业科学研究院,甘肃 定西 743000)
水是限制农作物生产力的主要因素[1],目前,全球超过三分之二的淡水资源被用于农业生产。在水资源日益紧张的现在,农业用水引起的社会矛盾也越来越突出。我国黄淮麦区,尤其是河北省,水资源非常匮乏。作为河北省的小麦科研单位,培育抗旱、节水小麦品种的任务尤为突出,使其在干旱条件下可以获得相当的成活率和收益,而在水分相对充足的条件下得到更高的产量。为达到此目标,鉴定和应用与抗旱相关的遗传资源是提高小麦抗旱性的关键[2],挖掘小麦抗旱基因、探究抗旱机理、提高植株水分利用效率是必由之路。
近年来,涉及植物非生物胁迫反应和耐受的信号网络已经得到了较好研究。已证实转录因子家族包括WRKY、NAC、AP2/ERF、bHLH和MYB等在植物应答非生物胁迫的表达调控中起着重要作用[3-8]。例如,在拟南芥中,NAC转录因子ATAF1通过活性氧信号通路影响非生物应激反应[9],它还通过影响DREB2A,调控水胁迫应答基因表达,调节植物的抗旱性[10]。在小麦中,过表达TaPIMP1(1个MYB基因)可以改变胁迫应答基因的响应强度,提高转基因小麦的耐旱能力[11]。在水稻中,已证明干旱诱导基因OsWRKY47通过调节CaBP和CRRSP11的表达,来提高植株的抗旱性[12]。
植物碳吸收与蒸腾作用强度密切相关。在植物干旱响应过程中,气孔发挥着核心作用,它可以调节蒸腾作用和CO2的吸收[13]。光合作用时,CO2和水蒸气在气孔中进行交换混合,这个过程受气体压力梯度和扩散系数影响[14-15]。在干旱条件下,植物蒸腾作用降低,为了使碳的同化吸收保持在较高水平,必须增加叶片中CO2的浓度梯度使其得到补偿,从而维持植株得到较高的生物学产量,这样也就提高了水分利用效率(WUE)。脱落酸(ABA)在提高植物水分利用效率中的核心作用是大家公认的。首先,水分利用效率受气孔密度和大小的影响,而气孔孔径由ABA信号控制。在干旱条件下,在植物中产生水势变化信号,此信号在植株内长距离快速传递,诱导ABA合成[16]。ABA含量的增加可以启动控制保卫细胞抗旱反应的信号通路[17],激活OPENSTOMATA1(SnRK2.6)。SnRK2.6是ABA信号转导通路中的核心调控元件,它通过磷酸化激活SLOW ANION CHANNEL ASSOCIATED 1因子,导致气孔关闭,减少蒸腾作用,提高植物的抗旱性[18-19]。有研究证实,提高ABA受体响应可以使植株在相同的水蒸发量下获得更高的生物产量,如在植物中同时表达ABA受体基因RCAR6/PYL12可以提高植株40%的水分利用效率[20]。其次,ABA还会促进叶片衰老,使营养优先供应新生组织和器官,提高植物在干旱胁迫条件下的存活率[21-22]。例如,异源表达RCARs[23-24]和降低PP2Cs的表达[25]可以减少植物蒸腾作用,提高转基因植株在干旱胁迫下的存活率。
本研究利用黄淮麦区主栽小麦品种或优异品系为试验材料,利用KASP标记检测技术对已报道的3个抗旱主效基因1-fehw3、TaDREb和CWI-4A进行等位基因分型,分析抗旱基因在不同环境下对小麦千粒质量的影响,以明确抗旱基因对产量性状的贡献率,为进行分子标记辅助选育抗旱小麦品种提供数据支撑。
1 材料和方法
1.1 供试材料
通过对黄淮麦区小麦种质材料的地理分布、系谱、表型和基因型等数据进行分析,筛选出352份有代表性的小麦品种作为试验材料。田间试验在河北省农林科学院粮油作物研究所堤上试验站进行,设2个处理,①正常灌溉条件:足墒播种,在小麦拔节期、灌浆期各浇1水,灌水量为750 m3/(hm2·次);②干旱处理:足墒播种,小麦全生育期不浇水。4 m行长,110粒,随机区组设计,3次重复。干旱处理的试验地与周边试验地设5 m的保护区,以减少周边试验地灌溉对其的影响。小麦千粒质量数据调查采用万深SC-G型自动种子考种分析仪进行。
1.2 KASP标记检测
挑选了3个抗旱主效基因,分别是1-fehw3[26]、TaDreb-B1[27]和Cwi-4A[28],利用KASP标记[29]对试验材料进行基因分型,引物序列如表1所示。以CTAB法提取供试材料叶片基因组DNA,加400 μL TE溶解。DNA经1%琼脂糖凝胶电泳进行质量检测,所提DNA要求无明显杂质、条带清晰、无降解。利用NanoDrop 2000(Thermo Fisher)检测DNA浓度,统一稀释为28.3 ng/μL,以稀释后的小麦基因组DNA为模板,进行PCR扩增。

表1 抗旱基因及KASP标记引物序列Tab.1 Drought resistant gene and KASP marker sequence
KASP标记引物工作液制备:取2条上游引物(100 μmol/L)各12 μL、下游引物(100 μmol/L)30 μL,用无菌超纯水补充至100 μL,作为KASP标记的引物工作液,-20 ℃保存备用。PCR扩增体系为:模板DNA 1.5 μL,引物工作液0.041 7 μL,2× KASP Master Mix(LGC公司,Lot No.13426773)0.75 μL,用无菌超纯水补充反应体系至3 μL。PCR反应程序为:94 ℃预变性15 min;94 ℃变性20 s、复性20 s(第1次的复性温度为61 ℃,每个循环降温0.6 ℃)共10个循环;94 ℃变性20 s、55 ℃复性1 min,共26个循环;72 ℃延伸3 min,4 ℃保存。
PCR反应完成后利用荧光信号阅读仪(Omega)和荧光检测系统(Araya)将荧光信号转变为可分析的数值对反应产物进行荧光数据读取。荧光扫描结果利用R软件包进行图形化展示,抗旱基因类型带有FAM荧光,分布于x轴附近;不抗旱基因类型带有HEX荧光,分布于y轴附近。对抗旱基因型和不抗旱基因型品种的千粒质量数据进行t-test检测,以明确这3个抗旱基因对小麦千粒质量性状的影响。
2 结果与分析
2.1 KASP标记检测效果
3个小麦抗旱基因(1-fehw3、TaDreb-B1和Cwi-4A)的KASP标记检测结果如图1所示。抗旱基因型带有FAM荧光,分布于x轴附近。不抗旱基因类型带有HEX荧光,分布于y轴附近。从结果看,每个基因2种不同基因型的小麦材料各分为1群,差异显著。利用KASP标记能够将不同基因类型的小麦材料进行有效区分,1-fehw3和Cwi-4A基因的分型结果比TaDreb-B1基因更优。
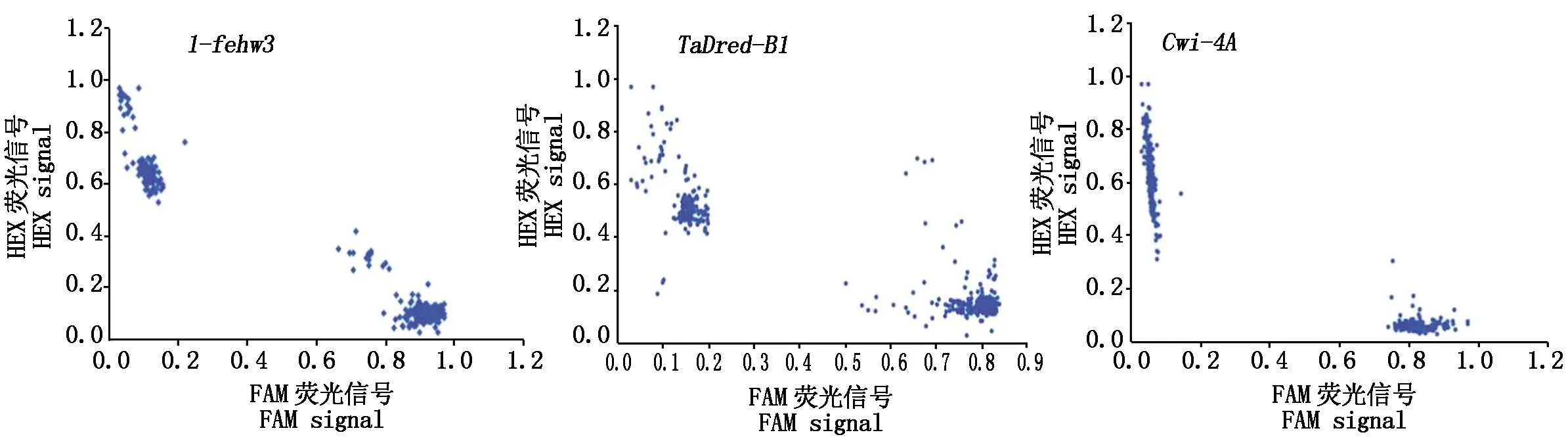
图1 小麦抗旱基因KASP标记检测结果Fig.1 KASP marker result of wheat drought resistance gene
2.2 单基因检测结果
通过连续3 a(2019—2021年)在正常灌溉和干旱条件下检测抗旱基因对小麦千粒质量的影响。结果分析显示,1-fehw3、TaDreb-B1和Cwi-4A3个基因的优势等位基因型分布频率分别为36.9%,41.1%,35.1%。但利用1-fehw3、TaDreb-B1和Cwi-4A3个基因单独检测时,在不同年份和不同条件下,抗旱基因型与不抗旱基因型品种间千粒质量均未达到显著水平(表2—4)。
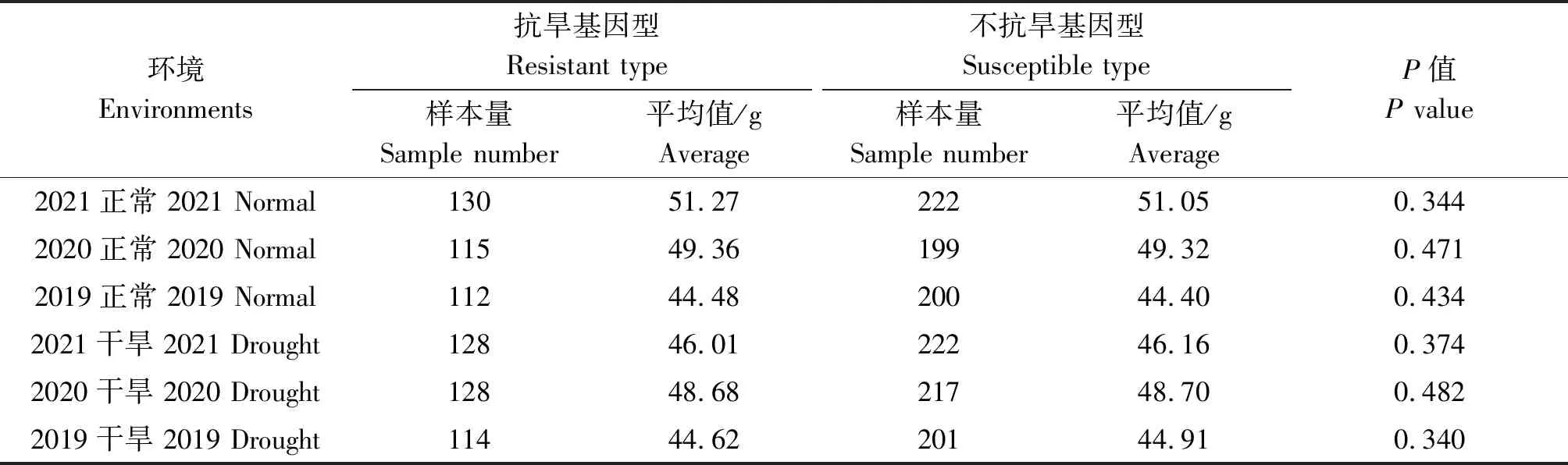
表2 不同环境下1-fehw3基因对小麦千粒质量的影响Tab.2 Effect of 1-fehw3 gene on thousand grains weight of wheat under different environments
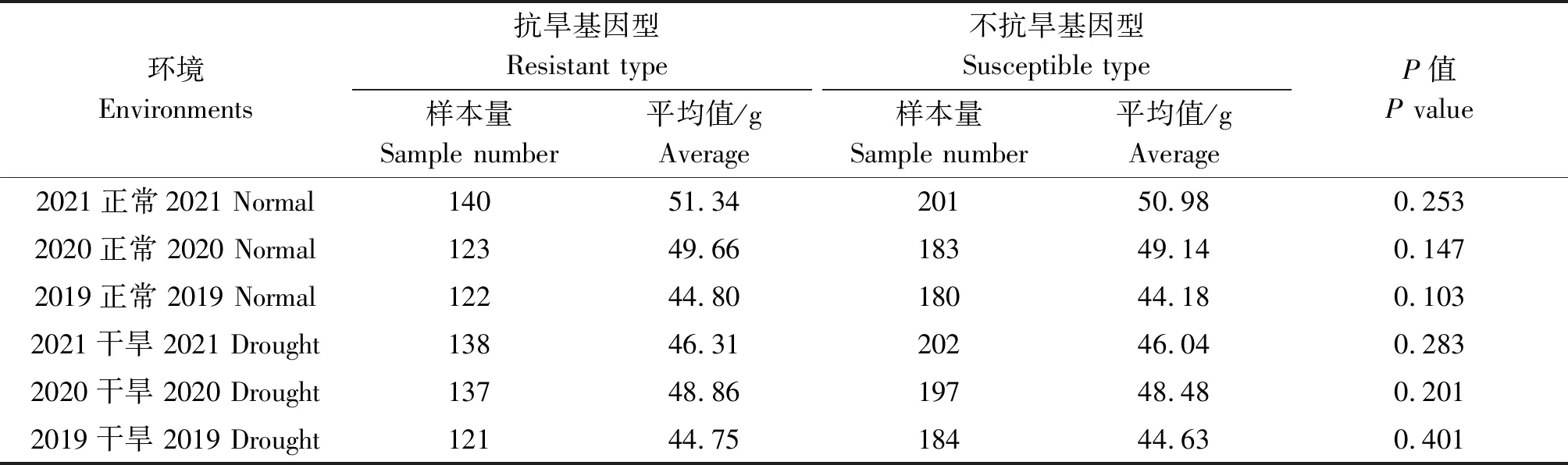
表3 不同环境下TaDreb-B1基因对小麦千粒质量的影响Tab.3 Effect of TaDreb-B1 gene on thousand grains weight of wheat under different environments
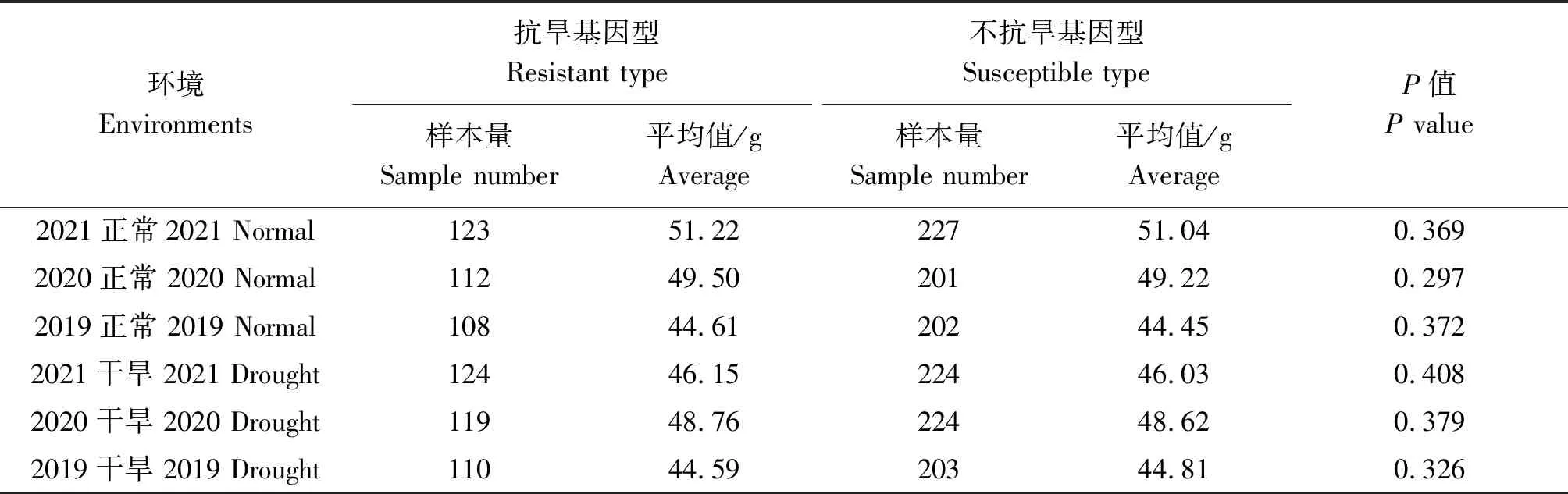
表4 不同环境下Cwi-4A基因对小麦千粒质量的影响Tab.4 Effect of Cwi-4A gene on thousand grains weight of wheat under different environments
2.3 双基因检测结果
当以2个基因进行联合检测时,显示1-fehw3+TaDreb-B1在2019年正常灌溉条件下抗旱基因型平均千粒质量为46.46 g,不抗旱基因型平均千粒质量为44.36 g;2020年干旱条件下抗旱基因型平均千粒质量为49.21 g,不抗旱基因型为47.30 g,2个环境下千粒质量均达到显著水平(表5)。当利用1-fehw3+Cwi-4A两基因进行联合检测时,在2019干旱、2020干旱两环境下抗旱基因型较不抗旱基因型千粒质量达到显著水平(表6)。利用TaDreb-B1+Cwi-4A两基因检测时,在2020干旱环境下抗旱基因型较不抗旱基因型千粒质量达到显著水平(表7)。
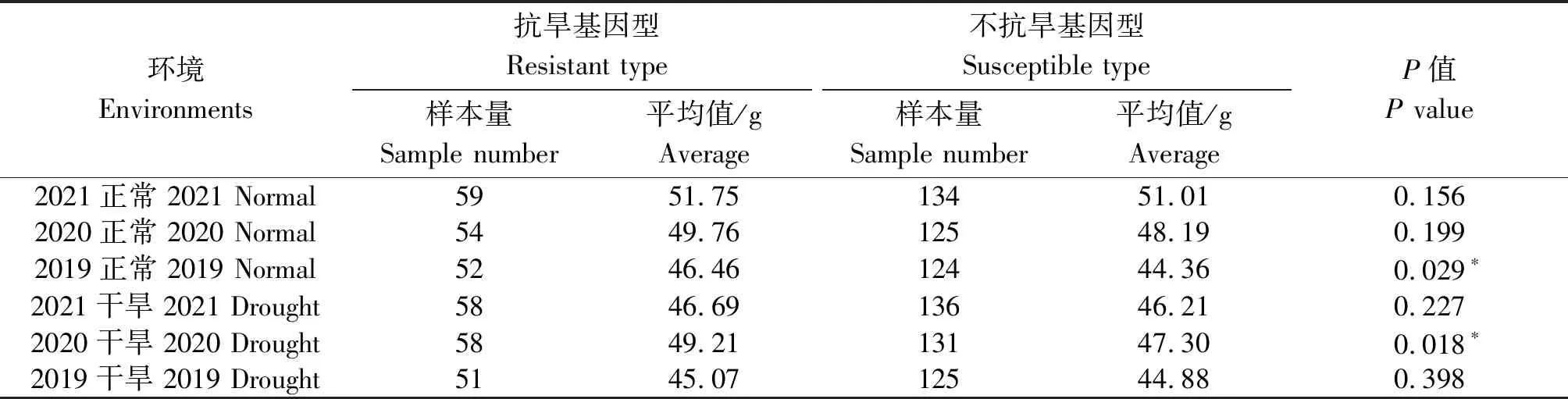
表5 不同环境下1-fehw3+TaDreb-B1基因对小麦千粒质量的影响Tab.5 Effect of 1-fehw3+TaDreb-B1 gene on thousand grains weight of wheat under different environments
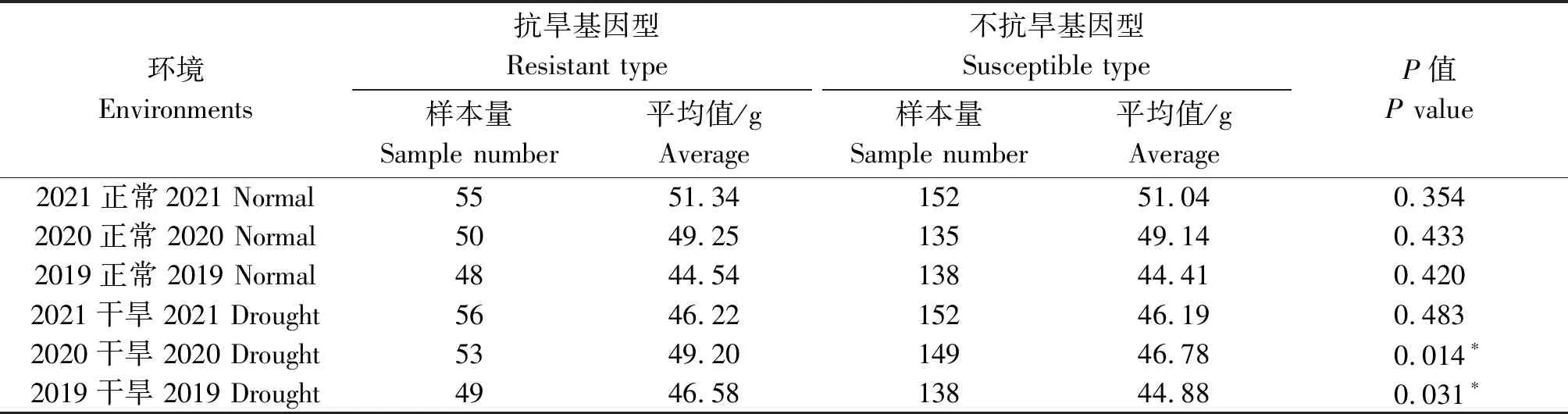
表6 不同环境下1-fehw3+Cwi-4A基因对小麦千粒质量的影响Tab.6 Effect of 1-fehw3+Cwi-4A gene on thousand grains weight of wheat under different environments
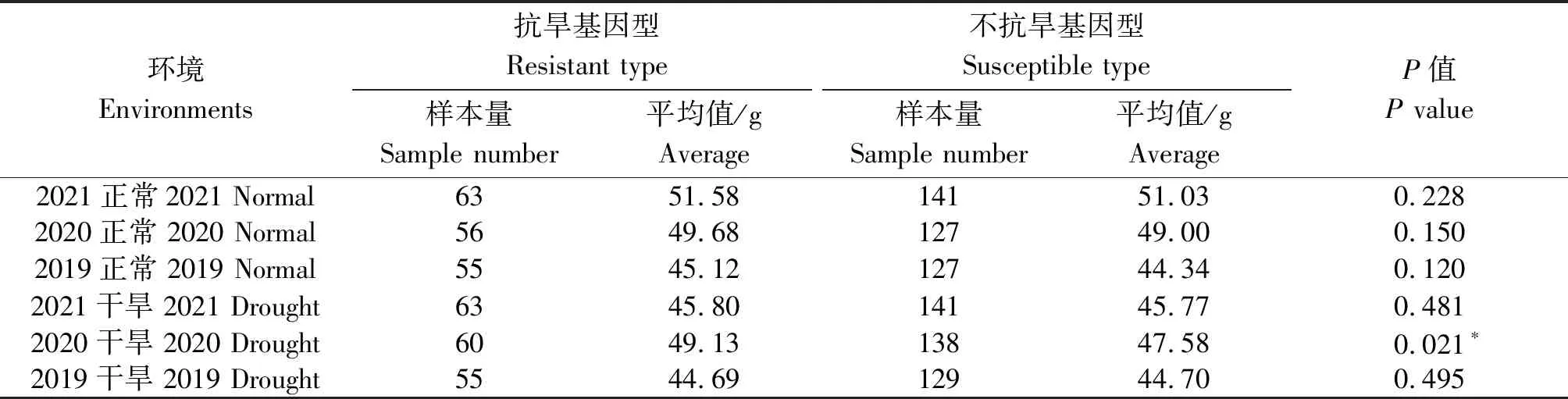
表7 不同环境下TaDreb-B1+Cwi-4A基因对小麦千粒质量的影响Tab.7 Effect of TaDreb-B1+Cwi-4A gene on thousand grains weight of wheat under different environments
2.4 三基因联合检测结果
当以1-fehw3、TaDreb-B1和Cwi-4A3个基因进行联合检测时,优势等位基因类型分布频率约占8.2%。除2020年正常灌溉条件之外,其余5个环境(2019年正常灌溉、2019年干旱、2020年干旱、2021年正常灌溉、2021年干旱)下,抗旱基因型平均千粒质量均高于不抗旱基因型,达到显著水平(表8)。
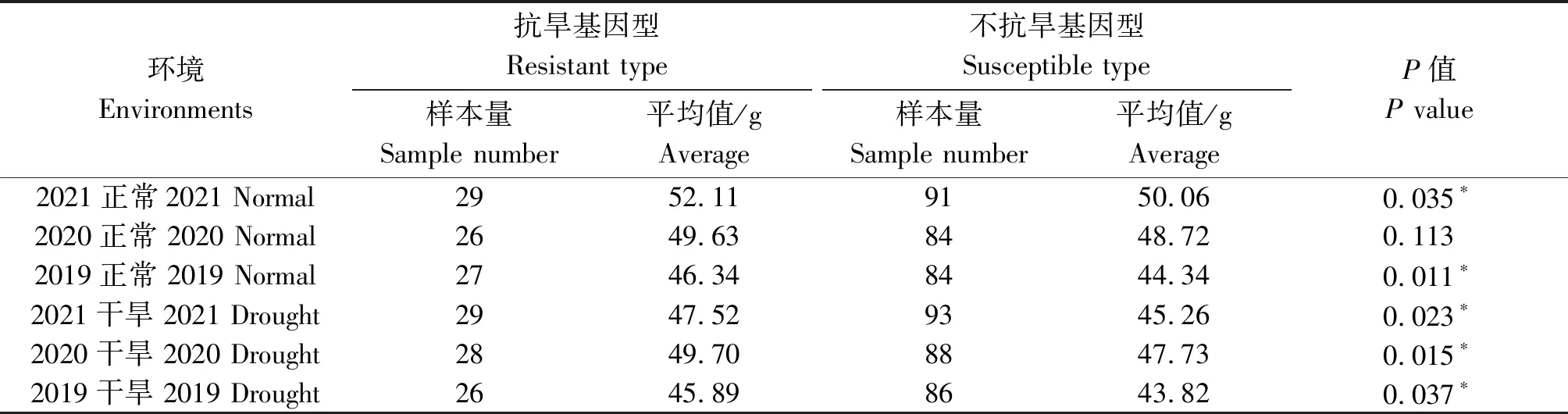
表8 不同环境下1-fehw3+TaDreb-B1+Cwi-4A基因对小麦千粒质量的影响Tab.8 Effect of 1-fehw3+TaDreb-B1+Cwi-4A gene on thousand grains weight of wheat under different environments
3 结论与讨论
小麦是我国三大作物之一,与我国粮食安全息息相关。为了满足人们不断增长的粮食需求,在过去几十年中育种工作者对小麦产量、品质、抗性、适应性等性状进行了改良,不仅使小麦表型产生明显变化,而且对重要基因的优异等位变异进行了强烈选择。我国黄淮麦区地下水资源严重缺乏,小麦的抗旱性对于小麦高产、稳产至关重要。小麦粒质量是小麦产量构成的三要素之一,提高粒质量是现代小麦育种的主要目标之一。通过分子生物学手段,鉴定小麦抗旱基因分布、研究抗旱基因对粒质量的贡献率,对于培育抗旱、高产小麦品种和多基因聚合育种都具有较高的应用价值和指导意义。
随着基因组学的迅速发展,特别是高通量检测技术的应用,利用小麦功能标记解析品种关键基因或优异等位变异成为可能,分子标记在小麦遗传改良中应用越来越广泛。目前,SNP标记由于其在基因组中的丰富性、共显性遗传、位点特异性、能够进行高通量基因分型等优点得到大家的认可,提高了小麦分子标记辅助育种的效率。KASP标记也逐渐取代了以往的RFLP、RAPD、AFLP、CAPS、SSR等分子标记,成为主要的分子标记类型。很多学者利用开发的KASP标记对小麦抗旱功能基因进行检测,促进了对抗旱小麦遗传基础解析和基因聚合育种的发展。如国审小麦品种漯麦18抗旱能力突出,袁谦等[30]为了深入了解漯麦18抗旱的分子机制,采用KASP标记分析了漯麦18含有的抗旱功能基因。结果表明,漯麦18同时含有1-fehw3、CWI-4A和CWI-5D3个抗旱基因。以上信息为深入认识漯麦18的特征性状的分子机制提供了理论支持,对品种的进一步应用和未来小麦遗传改良具有较强的实用价值。高振贤等[31]利用KASP标记检测了河北省153份审定小麦品种的光周期、春化、株高、粒质量、穗发芽、抗旱和抗病相关基因。结果表明,2个与抗旱相关的基因TaDREB-B1和1-fehw3优异等位变异占比分别是49.0%和39.9%。王志伟等[32]利用3个抗旱基因的KASP标记对云南育成的42份小麦品种(系)进行检测,显示23份材料为TaDreb-B1基因的耐旱单倍型,频率为54.76%;24份材料为1-fehw3基因的抗旱单倍型,频率为57.14%;35份材料为TaCwi-4A基因的抗旱单倍型,频率为83.33%。有11份材料为3个抗旱基因的抗性单倍型组合,频率为26.19%。本研究利用1-fehw3、TaDreb-B1和Cwi-4A3个抗旱基因的KASP标记对352份黄淮麦区小麦种质材料进行检测,1-fehw3、TaDreb-B1和Cwi-4A3个基因的优势等位基因型分布频率分别为36.9%,41.1%和35.1%,与高振贤等[31]的结果相似,低于王志伟等[32]的结果。这可能是由于试验材料的区域不同的原因,高振贤等[31]检测的河北省审定小麦品种,本研究所用试验材料来自黄淮麦区,包括河北省、河南省和山东省,所以结果相似。而王志伟等[32]的试验材料来自云南省,与黄淮麦区小麦材料差异较大。
本研究中,利用1-fehw3、TaDreb-B1和Cwi-4A3个基因单独检测时,试验结果均未达到显著差异,但同时利用3个基因联合检测时差异显著,说明作物的抗旱性是由多基因控制的复杂数量性状,单个基因起的作用较小,在利用单个基因进行检测时结果存在较大偏差,而同时利用2个或3个基因进行联合检测,可以大大提高分子标记选择的准确性。前人也报道过相似情况,如张海萍等[33]为探索我国小麦微核心种质及地方品种籽粒休眠的遗传基础,利用已报道的5个分子标记对107份我国小麦微核心种质及31份地方品种进行籽粒休眠的分子标记鉴定。结果表明,单个基因效应存在一定差异,而利用5个标记联合则可解释95.9%的性状变异。利用2个标记联合检测时的准确率也远远高于单个基因,如Vp1-b2和Xbarc294的组合可解释表型变异的89.1%。小麦抗白粉病基因Pm6具有良好的成株期抗性,但在苗期对一些小种的抗性较差,采用传统的方法难以很快鉴定Pm6基因的存在。Xwg996为共显性标记,与Pm6基因连锁而且专一性很强。KSUK948为互斥相标记,与Pm6基因连锁专一性较强。王瑞等[34]联合使用这2个标记有效地检测了Pm6基因的存在。同样,单独使用一个分子标记很难检测小麦抗白粉病基因Pm4,王俊美[35]利用2个标记STS463和STS410进行联合检测,即能够有效地进行Pm4基因的分子检测,有助于将Pm4基因准确导入农艺性状优良的感病品种。
