人为噪声对动物的非听觉影响*
2023-06-20邹建文金宝玲陈其才付子英
邹建文 金宝玲 陈其才 付子英
(华中师范大学生命科学学院,遗传调控与整合生物学湖北省重点实验室,武汉 430079)
工业化的发展,在给人类带来繁荣和进步的同时,也产生了越来越多的噪声。人为噪声(anthropogenic noise)主要是由人类活动或设施产生的,多为低频声音,如交通噪声、工业噪声、船舶噪声等,主要与4个相互关联的因素有关,分别是人口增长、城市化、交通工具和资源开采,预计这些因素的影响将会在未来几十年继续显著增加。如今噪声污染已经被列为主要的环境问题之一[1]。以往关于人为噪声污染的研究主要关注对象是人类。有很多证据表明,长期暴露在超过一定强度的噪声下,会对人类的健康产生负面影响,造成听力损失和一系列的非听觉影响[2]。比如:噪声会干扰睡眠,并由此产生慢性应激反应而最终导致病理生理变化,使心血管或代谢相关疾病的患病风险增大[3];环境噪声对儿童的学习和认知能力有负面影响,长期接触飞机、铁路、道路交通噪声的儿童,其阅读理解、记忆力和认知能力较差[4-5]。
最近,研究人员们意识到噪声也可能影响野生动物,并进行了相关的研究[6]。随着人类对海洋不断地探索和利用,人们还发现噪声污染不仅影响陆地上动物的生存,对于海洋生物的生理反应和行为等也会产生显著影响[7]。强烈的水下噪声,可能导致动物与压力相关的生理指标发生变化,造成身体损伤、行为变化,甚至死亡[8]。噪声在陆地生态系统和水生生态系统中普遍存在,大多数动物对噪声都比较敏感,所以噪声影响着各种动物的生存,包括两栖动物、节肢动物、鸟类、鱼类、哺乳动物、软体动物和爬行动物等[6]。其中研究最多的动物为鸟类,其次是水生哺乳动物和鱼类[9]。本文总结了人为噪声对动物造成的一系列非听觉影响,并对未来的可能研究方向进行了展望。
1 人为噪声对动物应激水平和生长发育的影响
噪声直接影响动物听觉系统导致噪声性听力损失[10],也会对动物的生理和发育等多个方面造成影响。
1.1 人为噪声对动物应激水平的影响
噪声是一种应激源,可能改变动物体内的生理状态而影响体内平衡。城市化进程不断推进的同时,野生动物的生活区域不断缩小,也使其更容易受到人类活动的影响[11]。已有许多研究证明,长期的噪声暴露会使动物产生应激反应,可能会影响下丘脑-垂体-肾上腺(hypothalamic-pituitaryadrenal,HPA)轴的发育,比如,暴露在100 dBA噪声(0~26 kHz)下的大鼠,其肾上腺皮质的束状带和网状带出现亚细胞结构上的损伤[12]。糖皮质激素在应激时从肾上腺分泌,是HPA 轴激活的结果[13-14](图1)。动物在应激后血清中的糖皮质激素会迅速升高,糖皮质激素包括皮质醇(cortisol)和皮质酮(corticosterone,CORT),在不同生理和应激状态下,皮质醇和皮质酮的变化趋势一致,都能作为应激的指标[15]。糖皮质激素的分泌对于HPA 轴有负反馈作用,主要作用于下丘脑和垂体。当HPA 轴的发育受到影响后,可能会使负反馈作用失效,从而使糖皮质激素长期处于高水平状态。暴露于重复噪声4周的猪,表现出血浆皮质醇和皮质类固醇结合球蛋白水平升高,以及促肾上腺皮质激素释放激素(corticotropin releasing hormone)和促肾上腺皮质激素(adrenocorticotrophic hormone)浓度升高,表明噪声可能通过HPA 轴上调皮质醇水平[16]。
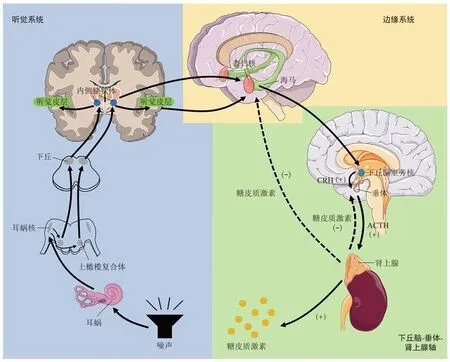
Fig. 1 Diagram of activation process of HPA axis图1 HPA轴的激活过程示意图
长期处在噪声环境中会使动物的皮质醇水平维持在较高的水平从而影响动物的生理机能,尤其是免疫能力和抗氧化防御能力。强烈的水下噪声会在短时间内对近距离的鱼类造成巨大伤害,长期的低声压级噪声同样会对鱼类的生存造成威胁[17]。长期暴露于压力源可能会影响鱼类的下丘脑-垂体-肾间组织(hypothalamus-pituitary-interrenal axis)轴,导致血浆皮质醇水平慢性升高、活性氧(reactive oxygen species,ROS)的积累和抗氧化分子的代偿性反应[17]。工业循环水产养殖系统(recirculation aquaculture system, RAS)中设备的运行噪声会影响鱼类生活。研究人员根据血液、肝脏和肠道样本的分析结果,探究了长期的RAS 噪声对幼年大口黑鲈(Micropterus salmoides)的生理影响。结果显示,长期的RAS 噪声环境下生长的大口黑鲈,其抗氧化酶指标和血清免疫指标都显著降低[18]。皮质酮可以通过影响血浆中白细胞和抗体的数量而对免疫能力产生影响[19],有研究发现交通噪声暴露会使学习敏感期内的斑胸草雀(Taeniopygia guttata)免疫能力下降[20]。
ROS 水平和抗氧化酶活性是反映动物是否处于逆境状态的实用生物学指标。 黑鲷(Acanthopagrus schlegelii)暴露于噪声环境中两周,表现出比对照组更高的ROS 水平,同时影响了肝脏抗氧化酶——过氧化氢酶(catalase,CAT)和谷胱甘肽过氧化物酶(glutathione peroxidase,GPx)的基因cat和gpx的表达;在噪声暴露24 h后,cat的mRNA 表达水平下降,在噪声暴露1 周后,gpx的mRNA 表达水平上调[21]。若抗氧化防御不足以平衡ROS的产生,就会发生氧化损伤。
噪声对鸟类的应激反应的影响也受到了研究者的关注。靠近公路的白冠带鹀(Zonotrichia leucophrys oriantha)雏鸟比远离公路的雏鸟有更高的生理应激反应,皮质酮水平随着与公路距离的减小而增加[22]。但在Crino 等[23]的研究中,与处于环境噪声中的对照组相比,白冠带鹀雏鸟暴露于交通噪声后15 min 的糖皮质激素水平显著降低。Kleist 等[24]进一步探究了噪声梯度对西蓝鸲(Sialia Mexicana)、山蓝鸲(Sialia currucoides)和灰喉蝇霸鹟(Myiarchus cinerascens)三种鸟类雌性个体与雏鸟的影响。结果显示,在噪声梯度下,无论是雌鸟还是雏鸟,皮质酮水平随着噪声梯度的增加而降低,两者之间呈负相关,而雏鸟急性应激源诱导的皮质酮水平升高,与噪声梯度之间呈正相关。目前尚不清楚产生这种截然相反结果的原因。
慢性噪声暴露对发育中的鸟类应激生理的影响仍有争议。Flore等[25]通过实验评估了交通噪声对早成鸟日本鹌鹑(Coturnix japonica)雏鸟皮质酮水平和应激反应的影响,结果显示暴露于交通噪声组((70.05±5.11) dB SPL (sound pressure level))与农村噪声组((63.33±6.54)dB SPL)的雏鸟皮质酮水平没有显著差异。有些依赖于城市生存或高度适应城市生活的动物可能会适应城市噪声,从而不受这些噪声暴露的影响,或是对刺激的反应不太强烈。Angelier 等[26]研究发现,从出生后第3 天开始,将麻雀(Passer domesticus)雏鸟每天在交通噪声(约61 dBA)下暴露6 h,在出生后第9 天时,其皮质酮水平并没有发生显著变化,且HPA 轴的发育也没有受到影响。粉红噪声暴露下的成年鹪鹩(Trogloytes aedon),其皮质酮水平在噪声暴露前后也没有发生明显变化[27]。同样,树燕(Tachycineta bicolor)雏鸟暴露于65 dB SPL的白噪声中,免疫水平和应激水平并没有发生显著改变[28],这与之前的许多研究结果不同,研究人员推测可能是由于噪声类型不同或是研究中可能存在其他影响因素,比如石油设备产生噪声的同时可能产生了刺激性气味或是化学污染。之后,又有研究人员通过交通噪声回放的方法测试了树燕在长期交通噪声下,其应激诱导的皮质酮水平和适应性指标受到的影响[29]。结果显示,交通噪声与自由生活的成年雌性树燕以及雏鸟的皮质醇水平改变有关。
噪声的强度、持续时间以及噪声类型都会影响动物对噪声的生理反应,所以在研究过程中不同的噪声暴露后可能会得到不同的结果,如糖皮质激素水平不变、下降或上升[29]。声压、时间和频率不规则的噪声会更容易使动物产生应激反应。最近,Grunst 等[30]研究了较为恒定的交通噪声和多种人为噪声组合的噪声对成年大山雀(Parus major)的影响,结果表明噪声暴露并没有对大山雀的后代的繁育行为产生显著影响。但噪声敏感性存在个体差异,一些个体对噪声暴露比其他个体敏感,特别是在暴露于多种噪声类型且持续时间不可预测的城市噪声情况下。因此,类似于高速公路噪声一样,较为一致的噪声产生模式的破坏性可能更小。在未来的研究中,系统的研究噪声对代谢的长期影响和动物的世代适应性将有助于了解相关的噪声反应的生理模式。
1.2 人为噪声影响雏鸟端粒的长短
噪声暴露能够在分子水平上影响动物。研究发现,处在城市环境的大山雀雏鸟的端粒长度更短,在发育过程中的噪声暴露会增加树燕雏鸟的端粒磨损程度[31-32]。端粒及其相关指标可反映生物体的生理状态[33],生命早期的端粒长度与寿命间存在显著的相关性,端粒的缩短可能会对动物个体的寿命产生不可逆的影响[34]。对城市生活高度适应的麻雀,在交通噪声暴露下饲养的雏鸟与未暴露的雏鸟相比,端粒长度显著缩短[35]。然而,以斑胸草雀为实验动物的研究发现,在孵化后的第21~120天之间,端粒的缩短更大,而非雏鸟阶段[36]。在经过噪声暴露的雌性克氏副杜父鱼(Paracottus knerii)中,其内耳组织中的端粒酶活性显著降低,大脑延髓中的端粒相对长度显著缩短,而在雄性中却没有显著的差异。该实验进行于产卵后的夏季末和秋季,研究人员推测可能与克氏副杜父鱼雄性独特的照顾后代的策略有关[37]。
生命早期的噪声暴露也会影响动物其他相关的特征。类胡萝卜素与鸟类羽毛的着色有关,而且作为抗氧化剂,可能与抗氧化应激有关。因此,与噪声暴露相关的压力水平增加可能会降低基于类胡萝卜素的着色强度。研究人员对噪声暴露下大山雀的端粒长度和类胡萝卜素着色进行了检测,发现类胡萝卜素的色调与端粒长度之间存在正相关关系,可以作为生命早期端粒长度的信号[38]。
1.3 人为噪声影响大脑发育和学习、认知能力
动物会根据噪声实时改变发声。雄性白冠麻雀(Zonotrichia leucophyrs)雏鸟在没有噪声存在情况下的鸣曲学习没有偏好,而在噪声掩蔽的情况下,绝大多数(82%)的雄性雏鸟倾向学习高频、较少受到掩蔽的鸣曲。这表明避免噪声掩蔽有强烈的选择作用,且雄性雏鸟可能会遵循认知倾向,模仿最容易检测到的鸣曲[39]。
大量研究观察到城市栖息地的鸟类发声的适应性变化,这些变化增加了声信号的传递效率。噪声对发声学习相关的大脑区域——X 区(area X)相对体积以及高级发声中枢(high vocal center,HVC)相对体积发育影响的研究表明,在早期暴露于交通噪声中的雄性斑胸草雀的这些区域要小一些[40]。tau蛋白在学习和记忆中起重要作用,中等强度的噪声(80 dB SPL)暴露会造成小鼠下丘(inferior colliculus,IC)、听皮层(auditory cortex,AC)和海马(hippocampus)的氧化损伤和tau 蛋白的过度磷酸化,损害小鼠的学习和记忆能力,且暴露时间越长,损害越大[41]。连续20 d 暴露于110 dB SPL 白噪声1 h 的C57BL/6 小鼠的空间学习和记忆能力受损,其海马中存在更多过度磷酸化的tau 蛋白和脂褐质,且脂褐质不会被降解,它的累积会降低细胞的正常功能和存活率[42]。噪声污染可能干扰生活在城市地区或道路附近动物的认知功能,研究人员通过研究交通噪声对成年斑胸草雀完成觅食任务的影响,发现交通噪声会降低斑胸草雀的认知能力,包括抑制控制能力、运动学习能力、空间记忆能力和社会学习能力等,但不会降低其色彩联想学习能力[43]。
2 人为噪声对动物行为反应的影响
噪声对动物行为的影响主要包括觅食效率的降低、反捕食行为的变化、干扰信号交流、声信号的改变、防御行为的变化等。
2.1 人为噪声对动物觅食行为的影响
人为噪声会影响多种动物的觅食活动。目前存在3 种噪声干扰觅食的潜在机制假说,即声掩蔽、减少注意和噪声回避[44]。如果是因为声掩蔽,在噪声与猎物的声信息重合时,会降低动物觅食效率;如果是因为减少注意,则无论是噪声是否与猎物的声信息重合,都会降低动物觅食效率;如果是噪声回避的影响,动物会避开噪声觅食。
交通噪声会降低大多数回声定位蝙蝠的觅食效率。即使没有与猎物回声的频率重叠,回声定位蝙蝠也会避开噪声源并且减少觅食活动,这可能是因为噪声作为厌恶刺激使蝙蝠产生回避反应[44]。同样,黑冠山雀(Poecile atricapillus)也会回避噪声,改变捕食行为[45]。Allen 等[46]利用苍白洞蝠(Antrozous pallidus)为实验对象的研究发现噪声会分散觅食蝙蝠的注意力,降低其觅食效率。人工夜间照明和噪声的增加会对脊椎动物产生负面影响,比如增加被捕食风险和减少觅食行为,但由于光照带来的无脊椎动物食物资源和觅食机会的增加,使得增添光照在一定程度上可以抵消噪声带来的负面影响[47]。
人为噪声的掩蔽可能会影响捕食者与被捕食者之间的相互平衡。许多无脊椎动物通过声音信号来检测捕食者,有些会产生防御信号来避开或是阻止捕食者[48]。对长鳍近海乌贼(Doryteuthis pealeii)的研究发现,暴露于回放噪声的乌贼通常猎物捕获率较低,而且如果在追捕过程中出现噪声,乌贼可能会放弃追逐猎物[49]。鱼类也会需要通过获得捕食者发出的声音信息来躲避捕食者,而且许多鱼类会在觅食地与产卵地之间迁徙,噪声的存在也会使其因回避反应而偏离原本的迁徙路线[50]。Halfwerk 和van Oers[51]研究了人为噪声对大山雀觅食行为的影响,结果显示噪声会干扰觅食期间大山雀的视觉处理,延长其捕食过程。
2.2 人为噪声改变动物发声,影响声交流
许多动物会使用声信号来传递信息,噪声的存在可能会掩蔽重要的声信号,影响动物的交流,进而对动物的配偶选择、领地占据、育雏以及躲避捕食者等行为产生影响。在实验中暴露于噪声中的动物会改变发声信号,但调整的幅度和方向因物种而异[52]。
东方蝙蝠(Vespertilio sinensis)会增加发声幅度并降低声信号的复杂性(即单音节呼叫的频率增加)以降低社交过程中的噪声干扰[53]。鸟类在人为噪声存在时会改变其鸣曲的频率、幅度、持续时间、节奏等。灰胸绣眼鸟(Zosterops lateralis)在低频城市噪声下以较高的频率鸣唱且每秒的音节数量会下降[54]。红翅黑鹂(Agelaius phoeniceus)在交通噪声下会改变它们的鸣曲结构和鸣曲时间[55]。雄性家雀(Carpodacus mexicanus)会随着低频噪声水平的增加而改变其鸣曲的最低频率[56]。然而,Zollinger 等[57]的研究结果却发现,在大山雀发育期间的低频噪声暴露不会导致鸣曲的频率低于成年鸟的频率。
与安静环境中的雏鸟相比,在噪声暴露的地区,树燕雏鸟的体型减小,反映身体状况的相关指标下降,对雏鸟的成活率产生负面影响[29]。当亲鸟带来食物时,在嘈杂环境下的雏鸟会有更大的可能无法得到食物,一方面可能是噪声掩盖了亲鸟到来的声音,使得雏鸟很难将亲鸟产生的声音与其他无关刺激区分开;另一方面,噪声也可能使雏鸟惊慌或分心,这些因素都可能影响到雏鸟的生存[58]。
船舶是常见的人为噪声的来源,生活在船只经过区域的鱼类,长期暴露在低强度的噪声中,可能会对其行为产生影响。 大西洋白姑鱼(Argyrosomus regius)在繁殖季节会发出响亮的合唱促进群体的聚集和交配,Vieira 等[59]研究了在实验室条件下,船只噪声对鱼类发声行为的影响。结果表明,船只噪声可以掩盖特定的叫声,并可能干扰船只通过期间大西洋白姑鱼的合唱行为。因为声信号无法被有效感知,噪声的掩蔽会导致呼叫者的通信空间(一个特定物种可以检测到另一个物种的范围)的减少。布氏鲸(Balaenoptera edeni)的通信空间在船只通过期间减少了近87.4%,在觅食时使用接触呼叫来保持群体凝聚力的大眼鲷(Pempheris adspersa)在船只通过期间的通信空间减少了61.5%[60]。
2.3 人为噪声影响动物的防御行为
噪声的存在会影响许多水生生物的水下活动。白尾雀鲷(Pomacentrus chrysurus)在摩托艇和船舶噪声下的日常游泳和逃生反应会受到影响,增加了个体的反应延迟[61]。欧洲鲦鱼(Phoxinus phoxinus)在复杂性不同的噪声下会出现惊吓反应并降低它们的群体平均游泳速度[62]。在对大口黑鲈的研究中,RAS 噪声使鱼群游动的不规则性增加,增大了鱼群的能量消耗,进而增加了这些鱼类的生存成本[18]。暴露于摩托艇噪声的多刺棘光鳃鲷(Acanthochromis polyacanthus)的防御行为会增加,同时减少了对幼崽的喂养和互动,这不利于后代的成长,降低了后代的存活率[63]。
鸣叫声在鸟类中不仅只有交流的作用,还起着传递捕食者信息的防御作用。许多动物依靠警报声来阻止捕食者,向同种动物寻求帮助或是给予同伴危险警告。人为噪声的存在可能会削弱动物的反捕食行为。黑冠山雀能够很好地适应城市地区,在交通噪声下会发出持续时间更长、峰值频率更低的警报声[64]。稀树草鹀在检测到捕食者时,会发出连续的警报声,其他个体为避免暴露巢穴位置,返回巢穴的时间会延长,也就是延长觅食时间,而噪声的存在则会降低它们的警惕度[65]。
2.4 人为噪声影响动物睡眠
睡眠在体能恢复、细胞修复和记忆巩固等过程中起着至关重要的作用。交通噪声对许多动物的睡眠行为产生负面影响。交通噪声减少了大山雀的睡眠时间并诱导鸟类在早上更早离开巢箱,而且对于老年个体的影响更强烈[66]。在正常情况下,黑背钟鹊(Crcticus tibicen)的非快速眼动(non-rapid eye movement) 睡 眠 和 快 速 眼 动(rapid eye movement)睡眠分别占夜间的(72.7±1.2)%和(12.2±1.0)%,其余(15.1±2.0)%处在清醒状态[67]。而夜间暴露于城市噪声中的黑背钟鹊会有更多的时间处在清醒中,清醒时间翻倍,睡眠碎片化,快速眼动睡眠减半至(5.6±1.3)%、非快速眼动睡眠减少至(61.6±3.2)%。
3 结语与展望
噪声会对许多动物的生理状态、行为反应等产生广泛影响(表1)。交通噪声、工业噪声和环境噪声是主要的人为噪声源。噪声对动物的影响与噪声的特征有关,比如频率、强度、持续时间等。以往的研究发现,野生动物对强度为40 dB SPL的噪声开始产生反应[9]。长期或频繁的噪声会影响动物对声音的检测,而间歇性的、不可预测的噪声对动物来说是一种恐吓或是威胁,二者都会直接或间接影响动物的健康[68]。那么,噪声特征与其对动物的影响存在怎样的对应关系?在某些特殊时期,如冬眠、繁殖期内,噪声是否会对动物产生更大的影响?这些问题的探究会帮助人类更有效地控制噪声。

Table 1 Non-auditory effects of anthropogenic noise on animals表1 人为噪声对动物的非听觉影响
噪声对动物产生的影响是多样而复杂的,可能会直接影响动物的生理应激水平、免疫功能、大脑的发育以及某些基因的表达。此外,噪声也会间接影响动物的行为,比如,强烈的噪声使动物感到威胁而逃跑或躲避、改变动物的觅食效率等。多数行为的改变都是适应性的,主要是因为噪声的存在对动物所造成的干扰、注意力分散和掩蔽效应等[68]。行为的变化和生理状态的改变是互相影响的,最终都会影响动物个体或群体的健康[69]。然而人为噪声所引起的生理改变与行为变化之间的关系,即行为变化的生理机制,目前仍缺乏认识。了解背后的机制,可能会更有助于了解各种动物行为受噪声的影响存在哪些差异?动物的哪些活动更容易受到噪声影响?从而采取有效措施减少对动物的不利影响。
目前有关人为噪声对动物影响的研究多集中于鸟类、鱼类等脊椎动物,而对陆地无脊椎动物和海洋无脊椎动物的关注则相对较少。地球上的无脊椎动物占动物物种总体的97%左右,在生态系统中发挥重要作用。人为噪声对于无脊椎动物的影响可能是广泛而复杂的,但只有4%关于噪声与野生动物的研究工作涉及到无脊椎动物[9]。未来的研究可以关注不同特征的噪声对各种无脊椎动物的各种影响[48],尤其是自然条件下的噪声是如何影响无脊椎动物的,这对于更全面地了解噪声污染对于动物的潜在影响,维护生态环境的平衡与稳定至关重要。
此外,现有研究显示,不同物种的听觉系统对噪声的敏感度存在较大差异,即存在噪声性听力损失的物种差异[10]。那么,噪声敏感度不同的物种,噪声诱发的生理改变,如应激水平,是否存在差异?这些差异与噪声性听力损失物种差异是否存在因果关系?一系列有趣而重要的问题仍有待进一步的研究去揭示。

续表1
