草地贪夜蛾β-Actin基因的原核表达及多克隆抗体制备
2023-06-14罗海玲夏顺超卢琴李海银
罗海玲 夏顺超 卢琴 李海银

摘 要: 草地贪夜蛾是一种重要的农业害虫,其迁飞性强、寄主广、繁殖力高,给我国农业经济带来了严重损失。从分子水平探究草地贪夜蛾生长、发育、繁殖、抗药性形成的内在分子机理,能为虫害的防治、防控提供重要参考。 β-Actin作为一种高度保守的看家蛋白常在分子生物学中用作内参去定量目标蛋白的表达水平,因而获得高质量的β-Actin抗体是从事相关分子研究的基本前提。因此,本研究克隆了草地贪夜蛾β-Actin基因部分编码序列,长度为579 bp。利用原核表达系统诱导获得了约37 kD的β-Actin重组蛋白,将纯化后的重组蛋白免疫新西兰大白兔制备得到β-Actin多克隆抗体血清。经ELISA检测,获得的抗血清效价达1∶ 256000。利用制备的β-Actin抗血清进行Western blot试验,结果显示在42 kD处出现强且单一的蛋白条带, 说明制备的β-Actin抗血清能有效识别草地贪夜蛾β-Actin蛋白。综上,本研究制备的β-Actin多克隆抗体效价高、特异性较强,能为草地贪夜蛾后续相关分子研究提供基本条件。
关键词:草地贪夜蛾;β-Actin;原核表达;多克隆抗体
中图分类号:Q965
文献标识码:A
文章编号:1008-0457(2023)02-0081-05
国际DOI编码:10.15958/j.cnki.sdnyswxb.2023.02.012
草地贪夜蛾 Spodoptera frugiperda ,属鳞翅目Lepidoptera,夜蛾科Noctuidae,是一种重要的杂食性农业害虫,其幼虫可危害玉米、水稻、高粱等禾本科作物[1]。草地贪夜蛾原产于美洲大陆[2],2018年底从缅甸入侵我国云南地区,随后在贵州各地陆续发现草地贪夜蛾[3],并逐渐向我国其他地区扩散[4]。草地贪夜蛾具有迁飞扩散性、寄主广泛性、高繁殖性、暴食为害性等特点,严重损害了我国农业经济[5-6]。目前,草地贪夜蛾的防治主要使用化学防治、农业防治、生物防治等,其中化学防治方法见效快,使用方法也比较简单方便[7],但随着化学药剂的大量使用和草地贪夜蛾的快速演进,草地贪夜蛾的抗药性也逐渐增强,急需开发更新型的绿色杀虫药物用以防治。伴随着分子生物学技术的发展,从分子水平探究草地贪夜蛾生长、发育、繁殖、抗药性形成的内在分子机制,以此为理论基础挖掘防治草地贪夜蛾的潜在分子靶标,开发新型绿色农药,能为草地贪夜蛾的防治提供新思路[8-9]。
在分子生物学技术中,蛋白质印迹(Western blot)是蛋白層面研究的一种基础且重要的技术,具有高敏感性和特异性的特点,可以从混合蛋白质中利用抗体检测出目的蛋白,并能进行定性或定量分析[10]。然而,该技术需要一个稳定表达的蛋白作为参考,以减少试验误差, 使结果更为信服。Actin,即肌动蛋白,几乎存在于所有真核细胞中,是许多真核细胞中含量最为丰富的蛋白质[11],作为细胞骨架的重要组分,参与细胞分裂、运动、生长等重要的生理活动[12-14]。根据Actin蛋白等电点的不同可以将其分为α、β、γ三种,其中β-Actin由于其高表达量和高稳定性,常被用作内参来比较不同来源目的蛋白的表达差异[15]。而在试验中检测β-Actin蛋白则需要抗体的特异性识别,因此一个高效、稳定的β-Actin抗体在蛋白分子研究中不可或缺。
本研究对草地贪夜蛾β-Actin基因部分编码序列进行克隆,通过体外重组表达、纯化得到草地贪夜蛾β-Actin重组蛋白,免疫新西兰大白兔获得草地贪夜蛾β-Actin多克隆抗体血清,并对抗体进行了效价检测,Western blot结果显示制备的抗体能特异性识别草地贪夜蛾β-Actin蛋白。 本研究制备的抗体能为草地贪夜蛾后续相关分子生物学研究提供必要条件。
1 材料与方法
1.1 材料及主要试剂仪器
1.1.1 供试材料
试验中使用的草地贪夜蛾采自贵州省黔西南望谟县(N 25°10′28.53″,E 106°05′40.64″),并经实验室人工饲养8代,饲养条件:温度:25 ℃,湿度:75%,光照:2000 lux,L∶ D=14∶ 10。
1.1.2 主要试剂
总RNA提取试剂盒(碧云天);反转录试剂盒 (Thermo);T4DNA连接酶(Thermo)、限制性内切酶EcoRⅠ和HindⅢ(Takara)、Taq DNA聚合酶(Takara); DNA胶回收试剂盒(上海生工)、质粒提取试剂盒(上海生工);His-tag蛋白纯化试剂盒(碧云天)。
1.1.3 主要仪器
低温超速离心机(Sigma)、PCR仪(Bio-Rad)、垂直蛋白电泳仪(Bio-Rad)、超声破碎仪(新芝)、超低温冰箱(捷盛)、酶标仪(Thermo)、超微量分光光度计(Thermo)。
1.2 试验方法
1.2.1 总RNA提取及cDNA合成
取草地贪夜蛾3龄幼虫整虫液氮研磨,使用RNA抽提试剂盒提取草地贪夜蛾的总RNA,经1%琼脂糖凝胶电泳验证草地贪夜蛾总RNA的完整性,经分光光度计检测总RNA浓度。取1 μg总RNA作为模板,Oligo d(T)18为引物,参照反转录试剂盒步骤进行逆转录合成cDNA。
1.2.2 PCR扩增与产物回收纯化
参照NCBI网站中草地贪夜蛾的β-Actin基因序列(登录号:HQ008727.1)设计原核表达引物(表1),以草地贪夜蛾cDNA为模板,利用原核表达引物进行PCR扩增。PCR程序:98 ℃变性2 min; 98 ℃变性15 s,60 ℃退火30 s,72 ℃延伸30 s,34个循环;72 ℃延伸5 min。将PCR产物经1%琼脂糖凝胶电泳,并利用试剂盒切胶回收纯化PCR产物。
1.2.3 原核表达载体的构建与鉴定
将纯化后的PCR产物和表达质粒pET-32a用内切酶EcoRⅠ和HindⅢ分别进行双酶切,纯化双酶切产物。在T4 DNA连接酶作用下将纯化后的基因片段连入pET-32a质粒,转化DH5α感受态细胞,并涂布于含有氨苄抗生素的LB固体培养基上37 ℃培养14~16 h。PCR鉴定阳性菌落,并测序验证序列正确性。挑选测序正确的阳性菌扩大培养,并抽提重组质粒。
1.2.4 重组蛋白诱导表达及纯化
将鉴定正确的重组质粒转化BL21感受态细胞,挑选单菌落入5 mL LB液体培养基,37 ℃培养至对数期(OD600值0.6~0.8),加入IPTG诱导表达6 h,同时设置不加IPTG诱导的菌液作为对照。用SDS-PAGE电泳检测蛋白表达情况。 将生长旺盛的种子菌接种到500 mL的LB液体培养基培养至对数期后,根据小规模表达的条件,加IPTG诱导6 h。收集大规模表达的菌液,利用裂解液和超声破碎细胞,参照碧云天His-tag蛋白纯化试剂盒进行蛋白纯化。测定纯化后蛋白浓度,并进行SDS-PAGE电泳检测蛋白纯度。-20 ℃保存纯化后蛋白。
1.2.5 抗体的制备及效价检测
采用皮下免疫法,将纯化后的β-Actin重组蛋白免疫新西兰大白兔获得抗体血清,该部分由武汉金开瑞生物公司完成。
将获得的抗体血清进行ELISA抗体效价检测。用包被液稀释抗原到2 μg/mL,以100 μL/孔加入96孔酶标反应板中,4 ℃过夜包被;洗涤液洗涤4次,加入封闭液,37 ℃温育2 h;洗涤液洗涤4次,用PBS按1∶ 2000,1∶ 4000,1∶ 8000,1∶ 16000等比例稀释血清,免疫前血清同比稀释做为阴性对照,每孔加入100 μL,37 ℃孵育1 h;洗涤液洗涤4次,加入HRP标记的羊抗兔IgG抗体,100 μL/孔,37 ℃孵育40 min;洗涤5次后拍干,加新鲜配制的底物溶液100 μL/孔,37 ℃遮光放置10~20 min;加50 μL/孔终止液,拍照并用酶标仪测定OD450。将抗体血清OD450/免疫前血清OD450的比值≥2.1的最高稀释度作为该抗血清的效价。
1.2.6 Western blot
提取草地贪夜蛾六龄幼虫总蛋白,Brandford法测定蛋白浓度,并制备蛋白样品。以3 μg、5 μg、8 μg总蛋白为浓度梯度进行SDS-PAGE电泳;湿转法在100 V,1 h条件下将蛋白转印到PVDF膜上;封闭液37 ℃孵育1 h;一抗(1∶ 3000)37 ℃孵育1 h;TBST洗4次,加入二抗(1∶ 10000)37 ℃孵育1.5 h;洗膜4次,ECL显影曝光,扫描光密度信号。
2 结果与分析
2.1 原核表达载体的构建
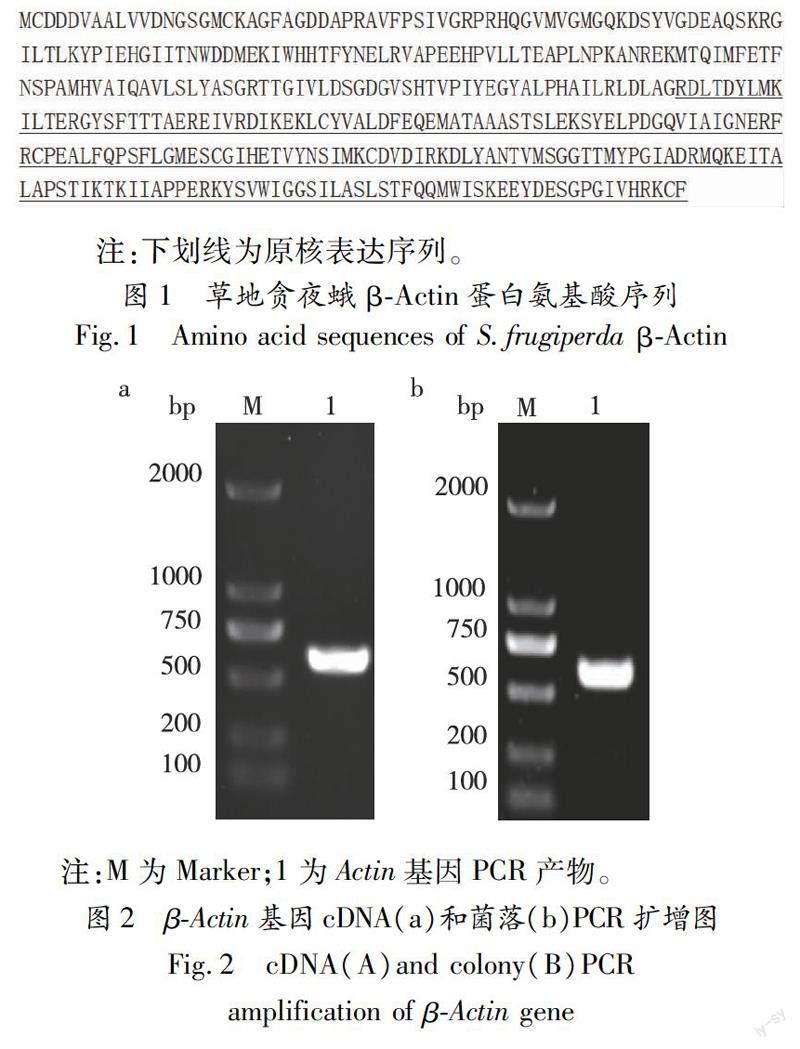
根据草地贪夜蛾基因组推定的β-Actin蛋白序列(登录号:AEK82131.1),选择184~376位氨基酸序列作为免疫原(图1),根据免疫原对应的编码序列设计引物进行PCR扩增,经1%琼脂糖凝胶电泳显示,在500~750 bp之间出现一条与理论长度(579 bp)大小一致的阳性条带(图2-a)。将所得的目的片段双酶切连入pET-32a表达载体,并转化DH5α感受态细胞。挑选单菌落进行PCR检测(图2-b),测序鉴定。
2.2 重组蛋白的诱导表达及纯化
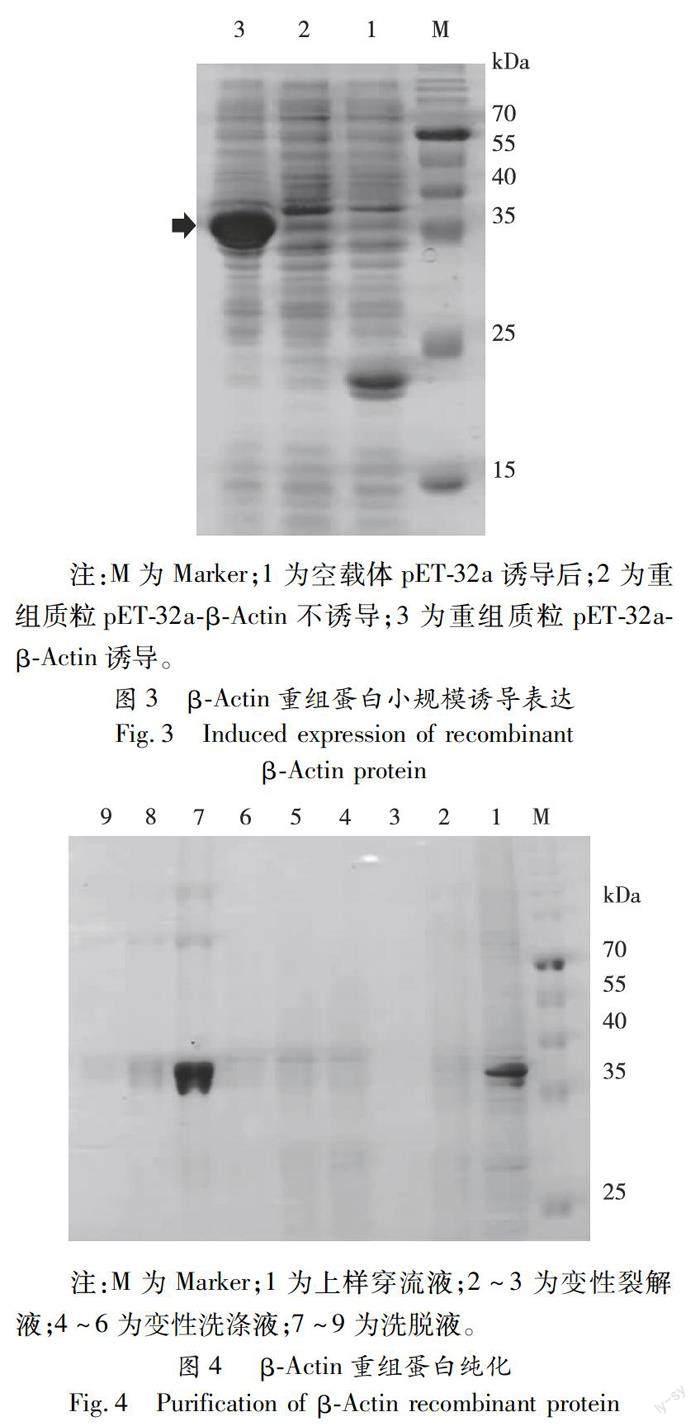
将构建正确的pET-32a-β-Actin重组质粒转化BL21感受态细胞,阳性菌株用终浓度为0.2 mM IPTG诱导重组蛋白表达。SDS-PAGE电泳结果表明,诱导表达后在37 kD处有一条明显增强蛋白条带,与预期重组蛋白(含载体自带硫氧还原蛋白和His标签)大小一致(图3),说明目的蛋白诱导表达成功。 将小规模诱导表达成功的菌液进行大规模诱导,收集诱导后的菌液,利用His-tag蛋白纯化试剂盒进行蛋白纯化,SDS-PAGE 电泳结果显示经高浓度咪唑洗脱的重组蛋白纯度较高(图4),符合免疫要求,将洗脱的重组蛋白免疫新西兰大白兔制备抗体。
2.3 ELISA检测抗体效价
将采集获得的抗体血清进行ELISA抗体效价检测。结果表明,随着血清稀释倍数的增加,OD450依次降低,β-Actin抗血清以1∶ 256000稀释后OD450值是阴性对照免疫前血清OD450的2.65倍(>2.1),由此判定制备的β-Actin抗血清效价为1∶ 256000(图5)。
2.4 Western blot检测草地贪夜蛾β-Actin蛋白
从草地贪夜蛾幼虫中提取总蛋白进行Western blot试验,结果显示在42 kD左右检測到一条明显的特异性蛋白条带,大小与预测的草地贪夜蛾β-Actin蛋白大小一致(图6),表明制备的多克隆抗体可以特异性识别草地贪夜蛾β-Actin蛋白。
3 结论与讨论
近半个世纪以来,分子生物学高速发展,从微观分子水平探寻生物体的发生机制已成为当前研究的主体[16],其中蛋白表达分析在发现新的调控分子机制方面发挥着重要的作用[17]。比较生命体不同细胞或组织的蛋白表达差异是研究其生物学现象和功能的重要基础,要准确判断蛋白表达的水平,就需要一个稳定表达的内参,使定性定量分析达到标准化[18]。β-Actin是生物进化过程中最保守的蛋白之一,在蛋白水平具有95%的一级结构保守性[19]。此外,β-Actin蛋白几乎在所有真核细胞中稳定高量表达,因此常被用作内参来标定其他蛋白的表达情况。在昆虫的分子生物学相关研究中,β-Actin是最常用的内参之一[20-22],但β-Actin蛋白的检测需要相应特异性抗体,目前市面上大多 数商品化的抗体主要针对哺乳动物细胞及组织,在昆虫(如草地贪夜蛾)体内特异性不强,且商品化内参抗体价格也较为昂贵[23],因而制备草地贪夜蛾β-Actin多克隆抗体是检测其他蛋白表达情况的前提条件。
本研究选用大肠杆菌原核表达系统进行β-Actin重组蛋白表达,该表达系统具有条件需求简单、使用安全、试验周期短、重组蛋白表达量高等优点[24-25]。本研究使用的表达载体质粒为pET-32a(+),该质粒自带硫氧还原蛋白,与目的蛋白融合表达可增加重组蛋白的水溶性,有利于提高蛋白表达量。此外,pET-32a(+)携带了6×His标签,在不影响目的蛋白结构和功能前提下,可与Ni+稳定螯合,为纯化目的蛋白提供了有利的条件。鉴于以上优势,本研究成功诱导表达并纯化了草地贪夜蛾β-Actin重组蛋白,并以此为抗原免疫新西兰大白兔,制备得到了β-Actin多克隆抗体。
抗体可分为多克隆抗体和单克隆抗体。相较于单克隆抗体,多克隆抗体的制备程序简单,效价更高,但与其他非靶标蛋白发生交叉反应的风险也较高[26]。本研究制备的β-Actin多克隆抗体经ELISA檢测抗体效价达到1∶ 256000,而Western blot结果证明制备的β-Actin多克隆抗体能有效识别草地贪夜蛾β-Actin蛋白,且未见有其他明显杂交条带,说明制备的多克隆抗体特异性强,与其他蛋白并不发生强的交叉反应。因此,本研究制备的草地贪夜蛾β-Actin多克隆抗体不仅可以作为内参在Western blot中来标定其他蛋白的表达情况,还能应用于对抗体要求较高的免疫组化、免疫荧光、免疫共沉淀等多种研究。
综上所述,本研究通过重组表达草地贪夜蛾β-Actin蛋白,并免疫新西兰大白兔获得多克隆抗体血清,经效价测定和Western blot检测证明制备的β-Actin抗体效价高、特异性好,可用于后续试验,能为草地贪夜蛾的深入研究提供良好基础。
(责任编辑:段丽丽)
参 考 文 献:
[1]
黄润,易军,李其勇,等.草地贪夜蛾综合防治[J].现代农业科技,2022,50(7):87-89.
[2] Sparks A N.A review of the biology of the fall armyworm [J].Florida Entomologist,1979,62(2):82-87.
[3] 刘利,郭晓关,谈孝凤,等.贵州省铜仁市草地贪夜蛾越冬情况调查[J].湖南农业科学,2021(5):54-58.
[4] 常向前,吕亮,万鹏.草地贪夜蛾在湖北的越冬研究初探[J].植物保护,2022,48(1):116-120.
[5] 姜玉英,刘 杰,朱晓明.草地贪夜蛾侵入我国的发生动态和未来趋势分析[J].中国植保导刊,2019,39(2):33-35.
[6] 胡朝兴,杨茂发,郅军锐,等.重大入侵害虫草地贪夜蛾对贵州省农业生产的潜在危害分析及其防控[J].山地农业生物学报,2019,38(3):1-5.
[7] 王燕平,向娟,袁娇,等.草地贪夜蛾的防治技术综述[J].四川农业科技,2021(9):28-29.
[8] Franz L,Raming K,Nauen R.Recombinant expression of ABCC2 variants confirms the importance of mutations in extracellular loop 4 for Cry1F resistance in fall armyworm[J].Toxins,2022,14(2):157.
[9] Boaventura D,Martin M,Pozzebon A,et al.Monitoring of Target-Site mutations conferring insecticide resistance in Spodoptera frugiperda [J].Insects,2020,11(8):545.
[10] 陈莹.家蚕常用内参蛋白多克隆抗体的制备及其检测[D].河南:南阳师范学院,2018.
[11] Dominguez R,Holmes K C.Actin structure and function[J].Annual Review of Biophysics,2011,40:169-186.
[12] Bunnell T M,Burbach B J,Shimizu Y,et al.β-actin specifically controls cell growth,migration,and the G-actin pool[J].Molecular Biology Cell,2011,22(21):4047-4058.
[13] Dugina V B,Shagieva G S,Shakhov A S,et al.The cytoplasmic actins in the regulation of endothelial cell function[J].International Journal of Molecular Sciences,2021,22(15):7836.
[14] Ma H M,Mai K S,Liu F U,et al.Cloning and characterization of an actin gene of Chlamys ferrei and the phylogenetic analysis of mollusk actins[J].Chinese Journal of Oceanology and Linmology,2007,25(3):304-309.
[15] Zhu H N,Zhang S Z,Zhou Y K,et al.Cloning and sequence analysis of the coding sequence of β-actin cDNA from the Chinese alligator and suitable internal reference primers from the β-Actin gene[J].Genetics and Molecular Research,2015,14(4):12159-12167.
[16] 齐书莹,朱圣庚.分子生物学发展趋势浅析[J].生命科学,1993,5(6):49-52.
[17] 牙库甫江·阿西木,关波,张富春.植物基因表达转录分析中内参基因的选择与应用[J].生物技术通报, 2011(7):7-11.
[18] Sikand K,Singh J,Ebron J S,et al.Housekeeping gene selection advisory:Glyceraldehyde-3-Phosphate dehydrogenase (GAPDH) and beta-Actin are targets of miR-644a[J].PloS One,2012,7(10):e47510.
[19] Gunning P W,Ghoshdastider U,Whitaker S,et al.The evolution of compositionally and functionally distinct actin filaments[J].Cell Science,2015,128(11):2009-2019.
[20] 唐婷,柳峰松,任國栋 .谢氏宽漠王 β-Actin基因cDNA克隆、 序列分析及表达量检测[J].昆虫学报 ,2008,52(11):1210-1215.
[21] 郭长宁,李鑫,于建光,等.金纹细蛾β-Actin基因克隆及表达分析[J].西北农业学报,2015,24(3):157-162.
[22] 张敏,张古忍.大头金蝇β-actin基因克隆与多克隆抗体制备[J].环境昆虫学报,2018,40(1):180-186.
[23] 刘妍,潘丽,秦贵信.肌动蛋白结构及生物学功能的研究进展[J].中国畜牧杂志,2020,56(1):34-38.
[24] Rosano G L,Ceccarelli E A.Recombinant protein expression in Escherichia coli :advances and challenges[J].Frontiers in Microbiology,2014,5:172.
[25] Porowińska D,Wujak M,Roszek K,et al.Prokaryotic expression systems[J].Postepy Higieny I Medycyny Doswiadczalnej,2013,67:119-129.
[26] Lipman N S,Jackson L R,Trudel L J,et al.Monoclonal versus polyclonal antibodies:Distinguishing characteristics,applications,and information resources[J].ILAR Journal,2005,46(3):258-268.
Prokaryotic Expression and Polyclonal Antibody Preparation of the β-Actin in Spodoptera frugiperda
Luo Hailing,Xia Shunchao,Lu Qin,Li Haiyin*
(Institute of Entomology,Guizhou University/The Provincial Key Laboratory for Agricultural Pest Management of Mountainous Region of Guizhou,Guiyang,Guizhou 550025,China)
Abstract:
Spodoptera frugiperda is an important agricultural pest.This pest has wide range of hosts,high reproduction,and strong immigration,which cause serious losses to the agricultural economy in China.The exploring of molecular mechanism of the growth,development,reproduction,and drug resistance of S.frugiperda,which could provide important information for the control of S.frugiperda.β-Actin,a highly conservative housekeeper protein,is often used as an internal reference to quantify the target protein expression frequently.Therefore,a high-quality β-Actin antibody is a prerequisite for protein research.In this study,we cloned the partial coding sequences of the β-Actin gene (579 bp) from S.frugiperda.A prokaryotic expression system was used to induce the β-Actin recombinant protein of about 37 kD.And then purified β-Actin recombinant proteins were used to immunize New Zealand white rabbit for preparing the antiserum β-Actin.ELISA analysis showed that the titer of the β-Actin antiserum was 1∶ 256000.Western blot showed that the β-Actin antibody could specifically recognize β-Actin protein (the band was about 42 kD).In conclusion,the β-Actin polyclonal antibody with high potency and specificity was generated successfully in this study,which will provide the basic conditions for further molecular research in S.frugiperda.
Keywords:
Spodoptera frugiperda; β-Actin; prokaryotic expression; polyclonal antibody
