森林光环境特征及其生态作用
2023-06-14金鹏徐明冉富菊雷云蒙春灿张健
金鹏 徐明 冉富菊 雷云 蒙春灿 张健

摘 要:光是森林生态系统的关键能源输入。森林光环境的演变机制很复杂,我们通过对林缘、林内和林窗的光环境相关指标及监测方法进行了归纳总结,并在分析森林光环境特征的基础上,从时间尺度上对其日变化、季节动态和年际变化三个方面进行了总结。同时从个体、群落、生态系统等方面对森林光环境生态作用进行了探讨,旨在为深入了解森林植被时空动态与光环境间协同演变机制提供参考,为促进森林可持续管理提供依据。
关键词:森林光环境;光環境特征;时空变化;生态作用
中图分类号:S718.5
文献标识码:A
文章编号:1008-0457(2023)02-0038-10
国际DOI编码:10.15958/j.cnki.sdnyswxb.2023.02.006
光作为一种重要的生态因子,也是动植物必需的自然资源之一。地球上所有生物的生长、发育和繁殖所需的能量都直接或间接来自光能。其中,光对植物的生理功能、形态结构、演替和分布等方面具有重要的生理生态作用,如植物光合作用是由光而引起的电子迁移作用,消耗了水和二氧化碳,把太阳能转化为化学能,制造氧气和有机化合物[1],对植物的物质积累起着重要作用。光照也会直接或间接影响到其他的生态因子,如温度、湿度、土壤物理性质等,进而影响植物的生长发育。因此,光对植物的影响具有综合性。
近年来,全球气候变化剧烈、极端天气频发、臭氧层的破坏等气象因素变化,极大的影响着光环境的变化,进而影响着人类与其他生物的生存与发展。高等植物在其生活史中均会受到光调节的作用,这种现象在全球各地普遍存在[2]。对于大部分的森林生态系统而言,光是调控森林植物生长发育的重要生态因子,对森林树种分布、生存和发展产生重要影响,因此,有必要加强森林光环境的作用机制与变化规律的研究,如探究不同光强、光质和光周期对森林植被的萌发、更新、生长发育以及不同演替阶段的影响,揭示不同干扰类型对森林及其光环境的影响,阐明森林光环境的时空变化特征及其与森林物种分布规律间的耦合关系等[3]。这将有利于依据森林光环境特点对珍稀濒危物种进行有效保护、提高林下经济作物的存活率和利用率、选择合适的抚育经营措施和补植作业方式促进干扰后的次生林的恢复演替。可见,加强森林生态系统光环境的生态监测研究,揭示森林光环境时空演变特征及其生态作用,将有助于增加人类对森林植被更新之间的关联机制、植被演替机制等方面的深入了解,进而促进森林可持续经营与科学管理。
1 森林典型生境类型的光环境特征
1.1 林内光环境特征
光照是影响森林群落林下植物生长与分布的重要生态因素[4]。一般而言,到达森林上方的光照只有少量能进入林内[5]。当森林冠层发生变化之后,复杂的森林群落通过对光的吸收、反射和透射作用,经过林冠层、冠层结构和树种叶片的空间分布、大小、方向及其动态变化造成了林下植被光照的情况[6-7],能强烈的改变光质、光强和光照时间[8],导致林下光照的光谱成分和分配发生不同程度的改变。当冠层开度小于55%~60%范围时,林下光照主要以散射光为主,冠层开度越小,散射光占主要地位越明显;冠层开度大于此范围时,林下光照以直射光为主要光源,且随冠层开度增大,直射光占主要地位越明显[9]。
除了散射光外,在郁闭的森林底层,有时也会接收到从林冠层空隙射下来的光强较高的太阳光照,其时间短,连续几秒钟或更长一些时间,称为光斑[10]。光斑很容易受外界的影响,如地球自转、冠层高度、树冠结构、风条件、 叶子聚集、树枝和叶子大小、地形位置和云量等因素[11],森林中的光斑更多地出现在落叶季节和上层冠层,可使林内光照情况差异增大,光环境的异质性增强。由于太阳辐照度经常被上层植物拦截或散射,导致林下光斑扩散,到达森林地面的光斑时间和强度大打折扣[12]。但光斑的分布可能是植物有效利用光能力的重要决定因素,因为光斑对林下植物可用于光合作用的光子通量密度贡献值达到10%~80%[13]。如在夏威夷常绿林中,林下总光子通量约40%是由光斑造成的,在相对晴朗的日子里,光斑点的贡献可能高达80%[11];在温带硬木和针叶林中,短时间的光斑可以提供季节性光合有效辐射总量的37%至68%[14];在马来西亚龙脑香森林中,在较长时间的光斑条件下,光斑占每日光通量密度总量的45%[15]。因此,林冠空隙形成的光斑对林下植物的光环境有重要影响,对林下植物分布、生命过程的维持有重要作用。
1.2 林缘光环境特征
林缘是从森林群落过渡到林外其他群落或生态系统的典型生态过渡区[16]。林缘的形成,会引起林缘及附近区域光照因子的强烈变化,形成不同的光环境,导致温度和水分等生态因子发生变化[17],造成明显的边缘效应。
一般来说,林缘的太阳辐射往往介于林外和森林内部之间,且从林外到林缘再到林内逐渐降低[18]。林缘光强变化趋势与林窗和林内相似, 随时间变化曲线为单峰型[19]。林冠对林缘光照的影响与距离林缘远近有关,不同林冠高度、郁闭度形成的林缘,其对周围植物光能利用的影响不同,进而影响物种丰富度、植物的根生物量与叶片厚度、叶片净光合速率、土壤微量元素[20]。林冠越高、郁闭度越大、距离林缘越近,林缘对植物光能利用影响就越大[17,21-22]。兴安落叶松林更新木从林缘到林内多为聚集分布,且林缘区域的更新木聚集度和树木数量比林内大,其林缘效应对兴安落叶松更新的影响大致在距离林缘20 m范围左右。此外,不同林缘方向对其附近区域小气候和植物生长的影响不同,光照条件好、光照时间长的南向林缘,更新情况更好[23]。
1.3 林窗光环境特征
林窗,即由各种原因如风灾、火灾、病虫害、酸雨、树木衰老、气候变化和人为干扰等造成一棵或多棵树死亡引起的林冠空隙[24],会极大地改变光照、温度、土壤湿度、养分可用性、凋落物深度以及与折断或连根拔起的树木相关的再生微生境的变化[26],增加林内光环境的异质性,是植物快速繁殖和生长的理想地点和条件。
林窗大小引起光、温、水、气等生态因子的变化,强烈影响着林下光环境,造成小气候的变化。一般认为,林窗内的光质较接近全光谱[25],太阳辐射随着林窗面积的增大而增强,即大林窗>小林窗,且光合有效辐射所占的比例也较大。Chazdon与Fetcher[26]在哥斯达黎加热带雨林的研究表明,一般林下的光合有效辐射是全光照的1%~2%,而在200 m2林窗中心则为9%,在400 m2的林窗中心则为20%~35%;在林窗内不同位置,光合有效辐射变化也有所不同,如林窗中心的光照强度最大,朝边缘逐渐减弱[25]。对于不同的林窗,林窗的方位也会影响到林窗的光照分布,例如南北走向林窗的光强水平大于东西走向的林窗[27]。在同一林窗中,其光强在南北方向上会呈现不对称性[28]。这与在林窗中心的南缘和北缘之间的小气候存在显著差异一致[29]。
林窗的空间分布决定了时间和空间上的光照情况,增加了幼苗的萌发和出苗。新的林窗形成依赖于位于整个森林的衰老树木[25]。林窗总面积有明显的下降趋势,这是因为随时间变化较小的林窗逐渐消失,较大的林窗趋于缩小[24]。在同一林窗中,由于太阳高度角、方位角、天气状况、各演替阶段林窗边缘乔木高度、林缘乔木冠层结构不同,林窗内不同时期、位置光照强度、光谱成分和透射率也存在差异[30]。此外,在同一类型的森林中,不同经、纬度也会导致林窗光状况的不同,如温带林窗的最大初始光强低于热带林窗[31],林窗的存在导致光资源分布不均匀[32],有了更多的边缘效应和物种多样性。
2 森林光环境的时间变化
2.1 森林光环境的日变化
对于森林群落而言,复杂的冠层是影响光因子的重要因素,对森林光环境有显著影响,主要分为直接影响和间接影响。其中,林窗对林下光产生的影响属于直接影响[33],而冠层通过影响亚冠层从而影响林下光照属于间接影响[34]。在林窗和林下,光照在一天之中的变化,中午光照最强,早晚最弱。在热带雨林和常绿阔叶林中,在晴天下光照强度日变化为单峰形,在中午达到最大值。林下、林窗、旷地中光合有效辐射随着太阳的升起迅速增加,并均在12∶ 00左右达到最大值[35],由于林窗和林下受冠层的影响,在早上10∶ 00点后光合有效辐射才明显升高,林下和林窗环境达到最大光合有效辐射最大值均迟于旷地[36],且旷地>林窗>林下[37];此外,光强还受到天气和云层的影响,在阴天下光照强度变化有明显波动,受云层厚度的影响大,晴天的光强明显高于阴天[38]。
2.2 森林光环境的季节变化
在北半球温带地区,一年中以夏天光照最强,冬季最弱;由于太阳高度的变化所造成的昼夜长短在各地是不同的,日照长度随纬度变化而进行着周期性变化。纬度越低,最长日和最短日光照差距越小。随着纬度的增加,最长日和最短日的差距越来越大,即纬度越高日照长短的变化越明显。
一般而言,落叶阔叶林的光照波动幅度最大,常绿阔叶林最稳定。常绿阔叶林的冠层面积生长快,冠层对光的截获能力提高,林下的光照条件较差[39]。在稳定的常绿阔叶林中,林下光照年变化不大,一年中林下光照以1月份最低,4月份和7月份较高,林下总光照与林冠开度和上层光照呈极显著的相关关系,且与林冠上层光照的相关性大于林冠开度[40]。对于常绿松树而言,其在生长和非生长季节都有针叶,因此它们可以全年在森林树冠下保持低光照条件,包括非生长季节[41]。一年中橡胶林各层光合有效辐射月总量在7月达到最大值,在1月或12月为最小值[42]。在温带森林,随着季节的变化,林冠叶片抽芽、展叶,冠层开度降低,叶面积指数增加,冠层的消光能力增强,林下光照强度降低;当林冠变黄脱落时,光强逐渐恢复,导致林下光照具有明显的季节差异,春季萌芽期和秋季落叶期的林下光照强度要明显高于生长夏季林冠郁闭期[43]。陈梅等[28]发现辽东次生林林窗在整个生长季各个林窗中的平均光合有效辐射均表现为春季>夏季>秋季,春季的平均光合有效辐射与夏秋季有显著差异[28];孙思焱[44]对金丝楸人工纯林林分研究则表明,春夏秋三个季节的光合有效辐射是一种抛物线的趋势。三个季节的光合有效辐射日变化曲线均为单峰曲线,峰值相比是夏季>春季>秋季。周光等[43]发现原始阔叶红松林和次生杨桦林光合有效辐射月积累量的变化趋势大致相同,均为双峰曲线,并且两个林分月累积量的最大值均分别位于4月和10月。
落叶林中与冠层树木相比,林下树为会通过物候避免树冠阴影:拥有较长的叶寿命,在春季较早完成叶片萌发这些活动,在秋季的叶片衰老和掉落则较晚来避免部分冠层树的叶片所施加的弱光环境。但由于早春和晚秋的低温限制叶片光合作用速率、较老的叶子的光合能力低于幼叶[45]、林冠树的衰老叶子仍会投下阴影,导致他们合成的有机物的速率有限。一般来说,冠层树幼苗的叶物候学倾向于更类似于亚冠层树,而不是冠层树,这些生理和形态的变化有助于其在各个生长阶段最大程度地减少光的拦截并增加碳同化效率[47]。
2.3 森林光环境的年际变化
光是树木幼苗定居、生长和生存的重要影响因子。在演替过程中,光照条件线性变化导致物种不断更新,是促进物种替代的主要驱动因子之一[47]。森林群落上方的辐射总量在不同演替阶段的差别很小[48],但不同冠层树种的生物学特性,如叶片特性、分枝结构等决定着群落内部的光环境差异,如常绿阔叶林不同演替阶段林冠层、林下光照呈先上升后下降的趋势[49]。
森林不同演替阶段的光环境存在明显差异。森林演替早期阶段,由于缺乏植被的覆盖和缓冲,光直接到达地面,造成了近地表面高温、土壤含水量降低,演替早期物种在高光环境中生长良好,生长快速,有能力应对光环境的极端变化,形成健康的大型植物。而早期林下的演替物种能够改变对光强度的光合反应模式,使得它们在阴暗处生长时变得更像耐荫植物。草本植物紫草的个体在高光环境中失去了原來的叶子,并长出了比移植时植物上的叶子颜色浅得多的新叶[50]。在后期演替阶段郁闭的林分里,大部分光被林木冠层截获,光照强度明显降低,光斑和林窗成为影响林下植物完成光合、蒸腾、生长等生理过程的重要环境因子之一[48]。较强的垂直光梯度会限制光需求强的先驱物种的再生[47],而后期演替物种的耐荫程度通常较高,光合饱和所需的辐照度水平较低,具有较强的养分保持能力和耐荫能力,能在阴凉环境中生长良好,因此冠层下耐荫演替物种的比例增加。
3 森林光环境的空间变化
3.1 森林光环境的垂直梯度变化
在自然环境中,森林群落在垂直结构上通常可分为乔木层、灌木层、草本层和地衣苔藓层。森林冠层在森林群落中具有重要意义,因树冠结构和形状、健康状况等因素影响冠层对光的捕获效率,导致不同冠层光环境存在明显差异。随着林冠深度的增加,由于枝叶对光合有效辐射的明显削弱作用,光合有效辐射量明显减少[51]。随着森林演替的进行,这种光梯度通常会变得越来越陡峭[52]。林冠上方全光照条件下的光合有效辐射最高,乔木层获得大部分光合有效辐射,到达灌层的光合有效辐射仅有少数。其中包括林分类型、林分密度、树冠的冠形、树冠尺度(大小、面积以及体积等)、叶倾角等,都会影响光资源的再分配。一般来说,冠层叶面积指数越大,光截获率越大,林冠结构对林下散射光的影响比对林下直射光大。冠形的影响与林分密度有关,对低密度林分,窄形冠和平伸形冠日吸收的光合有效辐射较大,对高密度林分,平伸形树冠日吸收的光合有效辐射最大。
在热带雨林中,高大的常绿树冠强烈地消耗了光的可利用性并产生了强烈的垂直光环境梯度[47],这种垂直光环境梯度对树木和其他植物间的光竞争具有重要影响,进而影响着林下植物的生长策略。在亚热带常绿阔叶林中,栲树冠层光合有效辐射表现为树冠外部显著高于树冠内部,冠层上部叶片光合活性高,对强光利用能力强;而冠层中部和底部光合活性低但对弱光适应能力强,从而使冠层整体光能的利用效率最高[53]。
林下光照条件是限制草本植物生长的关键因子[54],光照度与草本植物的物种丰富度、底层植物的覆盖度以及幼苗的物种丰富度和覆盖度呈正相关[57]。一般沿林层向上物种多样性呈非线性变化趋势,灌木层最高,林冠上层最低,且各林层之间差异显著,一般灌木层中植物个体数量和丰富度最大[56]。
林窗下随冠层开度增加,灌木层植物物种丰富度和多样性指数均呈降低趋势,这是因为林下光照过强会引起光抑制,降低偏荫性灌木层植物的萌发和生长[57]。
3.2 森林光环境的水平梯度变化
森林群落作为一个具有镶嵌特性的异质体,水平结构为森林植物在林地上的分布状态与格局[58]。不同植物种群具有独特的分布格局,分布格局一般分为聚集分布、随机分布和均匀分布。其中,聚集分布是森林群落常见的水平分布格局,但理想的林木水平分布格局应为随机分布[59]。由于森林冠层上方光照条件变化不大,森林水平光环境能被林窗大小、深度、形状和分布、冠层郁闭度、林分混交度、角尺度、聚集指数、竞争指数、冠层树种和下层植物的叶面分布等因素影响[47]。林分水平的变化包括冠层高度增加和叶面积指数增加[60]。林下水平光照异质性影响着林分的更新过程,如种子萌发、生长以及幼苗和幼树的存活等,通过光生态位划分促进物种共存,从而促进物种多样性[47]。
微地形是指土壤坡面由于土壤侵蚀等作用形成的大小不等、性状各异的局部地形[61]。在林内微地形上,由于海拔、坡向、坡位、坡度等地形因子的影响[62],加上地表粗糙度、群落结构及物种分布等的不同,地表对太阳辐射的接受有较大影响,同时林窗内的微地形最明显的是树倒坑、树倒丘、树倒掘坑和地表隆起的树干等。地形差异影响光照、温度等环境因子的再分配,形成不同水平梯度变化,具有明显的空间分布特征[63]。如沟脊大、沟底小、阳坡大、阴坡小、坡度越大接受的太阳辐射量越少[64]。各微地形间树木的大小和密度有差异,特别是上部坡面和下部坡面各微地形单元之间差异明显[65]。由于各种微地形单元的存在,在空间上的分化提供的异质化的生境导致了区域丰富的物种和群落多样性的形成。
4 森林光环境的指标与测量方法
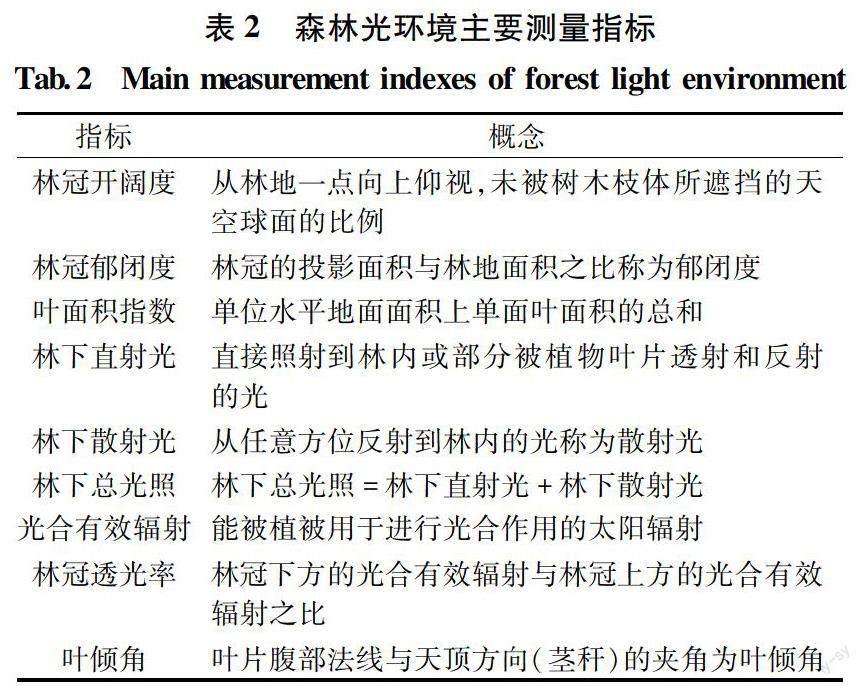
冠层作为与光接触的部位,其结构、形态、分布和动态变化对森林光环境有着明显的影响。由于冠层的复杂性,早期对森林冠层的描述多为定性描述,其效果并不理想,为了准确、定量的描述森林光环境,关于光环境的指标在不断的更新优化,近年来一系列的定量性的表述指标出现,如林冠开阔度、叶面积指数、郁闭度、林下直射光、林下散射光、林下总光照、光合有效辐射、林冠透光率、平均叶倾角等(表2),能够很好的记录冠层结构的特征。
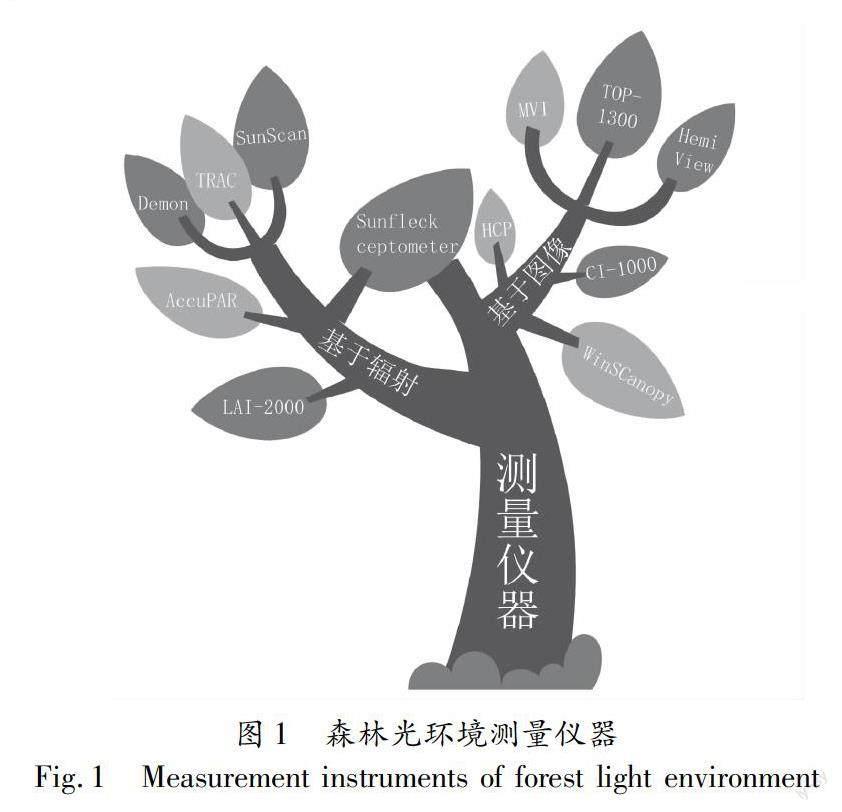
选择精确的测量仪器和合适的测量指标对于更有效的揭示森林光环境特征尤为重要。目前,根据森林光环境测量仪器原理可分为基于辐射测量的方法和基于图像测量的方法(图1)。基于辐射测量的方法仪器主要由辐射传感器和微处理器组成,能通过测定辐射透过率来计算相应指标。其次是基于图像测量的方法,通过鱼眼镜头和数码相机获取冠层图像,利用软件对冠层图像进行分析,获得一系列光学指标。仪器常由鱼眼镜头、数码相机、冠层图像分析软件和数据处理器组成[66]。通过光学仪器测量的方法虽然仪器价格相对较高、野外操作耗时费力、受人为因素和天气状况影响较大、维修不易等缺点,但因其具有快速、破坏性小、节省人力物力,效率高的优点得到广泛应用,且其测量精度和准度将随理论和技术的不断完善逐渐提高。
5 森林光环境的生态作用
5.1 影响植物生长发育
光是植物生长发育的主要因素之一,对植被的生活史有深远的影响,它不仅影响着植物的光合作用,还影响着有些种子萌发、幼苗的定居等。
有些种子萌发会受到光的影响[67-68],需要較适宜的光照环境,且不同演替阶段的植物种子对光照环境要求不同。在较低的光照条件下小粒种子一般不萌发,大粒种子在较低的光照条件下具有存活优势[69]。光环境对森林植被更新的影响不容小觑,适宜的光照条件是种子萌发和幼苗定居的关键。幼苗比成体对环境因子更敏感[70],光环境是幼树生长发育过程中最为重要的动态因子和限制性因子[71-72]。喜阳物种幼苗在光照充足的环境下才能更好的生存,耐荫物种则相反。
对于成体植物来说,光合作用能在弱光环境下促进叶片的光合色素含量和叶绿素b含量增加、叶绿素a/b的值[2,73-74]、单叶面积、叶面积比、叶重比、比叶重减小,同时叶片变薄,以便吸收更多的光合有效辐射[75-77],调节和维持叶绿素的含量,有利于缓解光资源不足对生长造成的影响。此外,强光会引起叶片的光抑制,是引起植物产生光抑制的直接原因,且效果十分显著[78]。一般情况下,随着光强的升高,叶片中叶绿素a、b含量均降低[79-81],降低叶绿素含量能有效避免吸收过量光能而引起强光抑制[82]。但也有些植物单位叶面积叶绿素含量不受光强影响[83]。光质能调节光合作用中不同类型叶绿素蛋白质的形成以及光系统之间电子传递[84],不同光质触发不同光受体,进而影响植物的光合特性、生长发育、抗逆和衰老等[85];此外,延长光周期还能显著地促进苗木的生长[86]。
5.2 影响森林群落演替发展、更新
群落演替是一种不断变化的动态过程,是指群落经过一定历史发展时期,由一种类型转变为另一种类型的顺序过程,也就是在一定区域内群落的发展替代过程,又称为生态演替,森林通常被认为是由不同发育阶段的独立斑块组成的嵌合体,这些斑块镶嵌体处于不断的动态变化之中,这些独立斑块在空间上分布格局被称为静态镶嵌,在时间上则构成动态演替。因此,森林成为一个空间上异质、时间上变动的“流动镶嵌体”[87],而林窗在“流动镶嵌体”中起着重要作用,表现在由于大树的死亡、倒伏形成的林窗提高了群落的存活率,树木生长形成森林,冠层郁闭度增加,林窗逐渐减小,直至消失,之后在林窗周围的成熟林中的大樹死亡,形成新的林窗,再一次进入了循环。在整个森林景观中,就形成了此起彼伏的斑块动态过程[88]。在这个过程中,林窗主要是影响森林光环境、植物的侵入和定居,并对群落结构和景观格局产生影响,进而影响着森林群落的演替。
许多群落的植被动态和演替主要是由资源吸收和种间抗逆性的差异驱动的,其中光往往是关键资源[89]。在演替过程中,光照条件线性变化,导致物种的连续和不断的替换[48]。早期演替是唯一的树冠不主导林地的时期,拥有更极端的温度、更强的光照、更大的风速以及表层土壤中的相对湿度和水分水平降低。这些环境指标的变化有利于某些物种,如喜光性植物,同时为其他耐荫性植物物种创造了次优或无法忍受的条件[90]。随着演替进行, 森林群落郁闭度越来越高、冠层结构愈加复杂、林下光照强度日益降低,喜光性植物种子和幼苗不能有效进行正常生长更替,只有干扰发生时去除上层的遮光才能顺利进入林冠层实现更新[68]。后期演替物种的耐荫程度通常较高,光合饱和所需的辐照度水平较低[91],林下幼苗能很好的利用林窗、光斑完成光合、蒸腾、生长等生理过程,占据更好的生态位,排斥演替早期树种。而中期演替物种在两种环境中都生长良好[51]
森林的自然更新方式主要包括种子更新和萌生更新。对于种子更新来说,有些种子萌发会受到光的影响[92],需要较适宜的光照环境,且不同演替阶段的植物种子对光照环境要求不同,对于萌生更新来说,其常普遍存在不利的环境条件和干扰胁迫下。总而言之,自然更新在某种程度上是群落演替的动力,群落演替往往又可以反作用于自然更新。自然更新与群落演替不是孤立的,而是相互作用的,它们之间的关系可以看成是个连锁反应[72]。
5.3 影响森林生态系统的组成
森林上层的光照环境差异不大,但由于冠层、林窗和林下光斑的存在, 导致林下光环境具有较高异质性,影响各种环境因子,形成不同的小生境。光照既可以直接影响对土壤微生物群落(细菌、真菌与原生生物)结构产生影响,如:多数大型真菌物种与低光照生境呈正相关,少数大型真菌显示出与低光照生境的负相关[93];也可以通过作用于植物群落对其产生额外的间接影响[94],光照可以影响林下土壤温度、养分等参数发生变化,最终导致微生物群落组成、分布的动态变化[95]。除了微生物外,光照也影响着昆虫的繁殖率,间接影响昆虫的分布和多样性[96],如在森林中,甲虫喜欢阳光充足的地方[97]。此外,光照还影响栖息在热带森林林下层的食虫鸟类分布,鸟类会根据光照环境选择合适的栖息地[98],不同种类的鸟对强光和弱光环境的选择不同,光环境的改变,可能影响许多森林鸟类的区域种群数量下降[99]。
光环境形成不同的小生境,不同物种有不同的形态、生理适应对策,这保证了不同树种在森林生态系统中的共存,允许具有不同生态要求的物种共存[100],从而影响生态系统的组成。
6 展望
森林光环境是森林环境的重要组成部分,加强森林光环境演变规律和调控技术的研究是森林生态学的一个重要研究领域。随着森林多重生态系统服务功能需求的日益加强,有关森林光环境变化规律和不同区域典型森林类型光环境特征的研究日益凸显,人们对森林光环境变化规律的了解,可为森林可持续经营与森林抚育管理,以及发展森林康养产业、林下经济(林下种植和林下养殖)等提供光环境参数,为森林资源的有序利用与科学管理提供参考。
森林是一个复杂的生态系统,其中各生态因子间存在着相互影响,因此,有必要加强各生态因子的综合性研究。森林生态系统处于一个动态的、长期的过程,其中光环境在森林种间竞争、苗木更新等方面发挥着重要作用,如从群落演替开始到演替结束的每个阶段,光因子影响作用都不一样,因此,除了重视光的时间变化外,还应该加强光对植被生活史不同阶段的研究。
(责任编辑:段丽丽)
参 考 文 献:
[1]
蒋丽金.生物光化学[J].有机化学,1983(2):83-91.
[2] 徐飞,郭卫华,徐伟红,等.不同光环境对麻栎和刺槐幼苗生长和光合特征的影响[J].生态学报,2010,30(12):3098-3107.
[3] Lochhead K D,Comeaup G.Relationships between forest structure,understorey light and regeneration in complex Douglas-fir dominated stands in south-eastern British Columbia[J].Forest Ecology and Management,2012,284(22):12-22.
[4] 姜闯道,高辉远,邹琦,等.叶角、光呼吸和热耗散协同作用减轻大豆幼叶光抑制[J].生态学报,2005,25(2):319-325.
[5] 王莉,林莎,李远航,等.青海大通典型林分冠层结构与林下植被物种多样性关系研究[J].西北植物学报,2019,39(3):524-533.
[6] Atsushi T,Atsushi K S Y.Methods for estimating understory light conditions using crown projection maps and topographic data[J].Ecological Research,2006,21(4):560-569.
[7] Sisira E,Singhakumara B M P,Mark S A.Variation in canopy structure,light and soil nutrition across elevation of a Sri Lankan tropical rain forest[J].Forest Ecology and Management,2008,256(6):1339-1349.
[8] 黃练忠,杨进良,徐庆华,等.城市森林群落林冠结构与林下光环境的关系[J].中南林业科技大学学报,2019,39(9):53-58.
[9] 杨建宇.古田山常绿阔叶林冠层结构及其光环境特征[D].齐齐哈尔:齐齐哈尔大学,2015.
[10] Way D A,Pearcy R W.Sunflecks in trees and forests:from photosynthetic physiology fo global change biology[J].Tree Physiology,2012,32(9):1066-1081.
[11] Pearcy R W.The light environment and growth of C3 and C4 tree species in the understory of a Hawaiian forest[J].Oecologia,1983,58(1):19-25.
[12] Youngkeun S,Youngryel R.Seasonal changes in vertical canopy structure in a temperate broadleaved forest in Korea[J].Ecological Research,2015,30(5):821-831.
[13] Chazdon R L.Light variation and carbon gain in rain forest understorey palms[J].Journal of Ecology,1986,74(4):995-1012.
[14] Jiquan Chen,Sari C S,Thomas R,et al.Microclimate in forest ecosystem and landscape ecology[J].BioScience,1999,49(4):288-297.
[15] Leakey A D B,Scholes J D,Press M C.Physiological and ecological significance of sunflecks for dipterocarp seedlings[J].Journal of Experimental Botany,2005,56(411):469-482.
[16] Nicholas V L B.Gap-phase regeneration in a tropical forest[J].Ecology,1985,66(3):682-687.
[17] 郭志华,张旭东,黄玲玲,等.落叶阔叶树种蒙古栎(Quercus mongolica)对林缘不同光环境光能和水分的利用[J].生态学报,2006,26(4):1047-1056.
[18] Sari C S.Modeling temperature gradients across edges over time in a managed landscape[J].Forest Ecology and Management,1999,117(1):17-31.
[19] 张一平,王进欣,刘玉洪,等.西双版纳不同季节热带次生林林窗光照时空分布特征[J].南京林业大学学报,2001,25(1):13-17.
[20] 王壮,杨万勤,吴福忠,等.高山森林林窗对苔藓及土壤微量元素含量的影响[J].生态学报,2018,38(6):2111-2118.
[21] 陈荟,梁康,马亨发,等.贵州月亮山自然保护区野生草本植物群落不同光环境物种多样性及结构特征[J].贵州科学,2022,40(1):82-86.
[22] 唐正.濒危植物大叶风吹楠生理特性、构件生物量及叶功能性状研究[D].长沙:中南林业科技大学,2020.
[23] 魏玉龙,李嘉悦,温晶,等.基于林缘效应对兴安落叶松天然更新及分布格局的研究[J].西北林学院学报,2019,34(3):15-20.
[24] Muscolo A,Bagnato S,Sidari M,et al.A review of the roles of forest canopy gaps[J].Journal of Forestry Research,2014,25(4):725-736.
[25] 王卓.长白山阔叶红松林林窗光强异质性对主要树种幼树光合作用的影响[D].北京:北京林业大学,2009.
[26] Chazdon R L,Fetcher N.Photosynthetic light environments in a lowland tropical rainforest in Costa Rica[J].Journal of Ecology,1984,72(2):553-564.
[27] Thomas L P,William J P.Gap light regimes influence canopy tree diversity[J].Ecology,1989,70(3):552-555.
[28] 陈梅,朱教君,闫巧玲,等.辽东山区次生林不同大小林窗光照特征比较[J].应用生态学报,2008,19(12):2555-2560.
[29] Troendle C A.The potential for water yield augmentation from forest management in the rocky mountain region1[J].JAWRA Journal of the American Water Resources Association,1983,19(3):359-373.
[30] 王进欣,张一平,马友鑫,等.西双版纳人工林林窗光照剖线分布特征[J].植物资源与环境学报,2000,9(1):28-31.
[31] 臧润国,徐化成,高文韬.红松阔叶林主要树种对林隙大小及其发育阶段更新反应规律的研究[J].林业科学,1999,35(3):4-11.
[32] Leakey A D B,Press M C,Scholes J D.Patterns of dynamic irradiance affect the photosynthetic capacity and growth of dipterocarp tree seedlings[J].Oecologia,2003,135(2):184-193.
[33] Denslow J S,Ellison A M,Sanford R E.Treefall gap size effects on above-and below-ground processes in a tropical wet forest[J].Journal of Ecology,1998,86(4):597-609.
[34] Montgomery R A,Chazdon R L.Forest structure,canopy architecture,and light transmittance in tropical wet forests[J].Ecology,2001,82(10):2707-2718.
[35] 郭志华,胡启鹏,王荣,等.喜树幼苗的叶悬挂角和叶柄角对不同光环境的响应和适应[J].林业科学研究,2006,19(5):647-652.
[36] 汤爱仪.常绿阔叶林不同演替階段树种幼苗对不同光环境的适应[D].重庆:西南大学,2008.
[37] 冉春燕.缙云山常绿阔叶林几个树种幼苗对不同光环境的响应与适应[D].重庆:西南大学,2006.
[38] 毛培利.海南岛热带山地雨林不同演替阶段植物功能群光生态适应性研究[D].北京:中国林业科学研究院,2007.
[39] 黄慧敏,董蓉,何丹妮,等.冠层结构和光环境的时空变化对紫耳箭竹种群特征的影响[J].应用生态学报,2018,29(7):2129-2138.
[40] 崔佳玉.林隙光照及对常绿阔叶林维管地被植物的影响[D].广州:华南农业大学,2016.
[41] 李俊清,李景文.中国东北小兴安岭阔叶红松林更新及其恢复研究[J].生态学报,2003,23(7):1268-1277.
[42] 吴志祥,杜莲英,谢贵水,等.海南岛橡胶林光合有效辐射的时空分布[J].西北林学院学报,2013,28(3):13-21.
[43] 周光,徐玮泽,万静,等.长白山阔叶红松林不同演替阶段林下红松幼苗能量与养分季节动态[J].应用生态学报,2021,32(5):1663-1672.
[44] 孙思焱.金丝楸人工纯林林分结构及幼苗光合特性分析[D].北京:北京林业大学,2020.
[45] Gill D S,Amthor J S,Bormann F H.Leaf phenology,photosynthesis,and the persistence of saplings and shrubs in a mature northern hardwood forest[J].Tree Physiology,1998,18(5):281-289.
[46] Augspurger C K,Bartlett E A.Differences in leaf phenology between juvenile and adult trees in a temperate deciduous forest[J].Tree Physiology,2003,23(8):517-525.
[47] Matsuo T.Forest structure drives changes in light heterogeneity during tropical secondary forest succession[J].Journal of Ecology,2021,109(8):2871-2884.
[48] 李庆康,马克平.植物群落演替过程中植物生理生态学特性及其主要环境因子的变化[J].植物生态学报,2002,26(z1):9-19.
[49] 李昊民,李宁云,喻庆国,等.天童山常绿阔叶林优势树种光合生理特性在其群落演替中的生态意义研究[J].西部林业科学,2011,40(2):13-20.
[50] Bazzaz F A,Carlson R W.Photosynthetic acclimation to variability in the light environment of early and late successional plants[J].Oecologia,1982,54(3):313-316.
[51] 李元,宋亮,莫雨轩,等.西双版纳热带季节雨林光合有效辐射的垂直分布格局和季节动态[J].生态学杂志,2022,41(9):1683-1690.
[52] Zhang Q,Zhang T J,Chow W S,et al.Photosynthetic characteristics and light energy conversions under different light environments in five tree species occupying dominant status at different stages of subtropical forest succession[J].Functional Plant Biology,2015,42(7):1-9.
[53] 孟陈,徐明策,李俊祥,等.栲树冠层光合生理特性的空间异质性[J].应用生态学报,2007,18(9):1932-1936.
[54] 段文标,王丽霞,陈立新,等.红松阔叶混交林林隙大小及光照对草本植物的影响[J].应用生态学报,2013,24(3):614-620.
[55] Tinya F,Márialigeti S,Király I,et al.The effect of light conditions on herbs,bryophytes and seedlings of temperate mixed forests in O ' ' rség,Western Hungary[J].Plant Ecology,2009,204(1):69-81.
[56] 樓一恺,范忆,戴其林,等.天目山常绿落叶阔叶林群落垂直结构与群落整体物种多样性的关系[J].生态学报,2021,41(21):8568-8577.
[57] 谭一波,何琴飞,郑威,等.珠江流域中上游防护林冠层结构对林下植被的影响[J].生态学杂志,2016,35(12):3148-3156.
[58] 韩有志,王政权.森林更新与空间异质性[J].应用生态学报,2002,13(5):615-619.
[59] 魏红洋,董灵波,刘兆刚.大兴安岭主要森林类型林分空间结构优化模拟[J].应用生态学报,2019,30(11):3824-3832.
[60] Brantley S T,Young D R.Contribution of sunflecks is minimal in expanding shrub thickets compared to temperate forest[J].Ecology,2009,90(4):1021-1029.
[61] 张宏芝,朱清科,赵磊磊,等.陕北黄土坡面微地形土壤化学性质[J].中国水土保持科学,2011,9(5):20-25.
[62] 王冬至,张志东,牟洪香,等.结构方程模型在落叶松林经营中的应用[J].北京林业大学学报,2015,37(3):69-75.
[63] 王家华,李建东.林窗研究进展[J].世界林业研究,2006,19(1):27-30.
[64] 魏胜龙,陈志彪,陈志强,等.微地形上太阳辐射模拟及与地表温度关系研究[J].国土资源遥感,2017,29(1):129-135.
[65] 杨永川,达良俊,由文辉.浙江天童国家森林公园微地形与植被结构的关系[J].生态学报,2005,25(11):38-48.
[66] 祁漫宇,朱维斌.叶面积指数主要测定方法和设备[J].安徽农业科学,2012,40(31):15097-15099
[67] 栗昕羽,朱梅,李晓乐,等.远红光辐照对干旱胁迫下生菜种子萌发及幼苗生长的影响[J].中国农业大学学报,2022,27(5):123-133.
[68] 潘婷,雷云,申玲芝,等.针阔混交林生态系统特征及生态效益分析[J].山地农业生物学报,2021,40(5):40-47.
[69] Swaine M D,Whitmore T C.On the definition of ecological species groups in tropical rain forests[J].Vegetation,2004,75(1):81-86.
[70] 张群,范少辉,沈海龙.红松混交林中红松幼树生长环境的研究进展及展望[J].林业科学研究,2003,16(2):216-224.
[71] 唐丽,李淳,骆凯,等.几种环境因子对光叶紫花苕种子萌发的影响[J].中国草地学报,2021,43(9):71-77.
[72] 程瑞梅,沈雅飞,封晓辉,等.森林自然更新研究进展[J].浙江农林大学学报,2018,35(5):955-967.
[73] 黄俊,郭世荣,吴震,等.弱光对不结球白菜光合特性与叶绿体超微结构的影响[J].应用生态学报,2007,18(2):352-358.
[74] 刘文海,高东升,束怀瑞.不同光强处理对设施桃树光合及荧光特性的影响[J].中国农业科学,2006,39(10):2069-2075.
[75] Scholes J D,Press M C,Zipperlen S W.Differences in light energy utilisation and dissipation between dipterocarp rain forest tree seedlings[J].Oecologia,1996,109(1):41-48.
[76] 张金玲,陈海鹏,程达,等.毛乌素沙地臭柏光抑制响应机制研究意义[J].干旱区研究,2018,35(4):882-890.
[77] 王振磊,陈海江,林敏娟,等.黄金梨和鸭梨叶片光合作用的光抑制及其恢复的比较研究[J].园艺学报,2009,36(9):1261-1268.
[78] 冯强,刘宁,王暾,等.不同森林光环境下4种灌木1年生幼苗形态和光合生理特性[J].中南林业科技大学学报,2016,36(5):93-101.
[79] 林凡,马松亚,潘锋,等.不同光照强度对格木苗木生长和生理生化特性的影响[J].安徽农业科学,2016,44(8):170-173.
[80] 周振翔,李志康,陈颖,等.叶绿素含量降低对水稻叶片光抑制与光合电子传递的影响[J].中国农业科学,2016,49(19):3709-3720.
[81] 张兰,王静,张金峰,等.辽东栎幼苗生长和生理特性对光照强度的响应[J].中南林业科技大学学报, 2021,41(11):73-81.
[82] Jan M A,Park Y I,Chow W S.Photoinactivation and photoprotection of photosystem II in nature[J].Physiologia Plantarum,1997,100(2):214-223.
[83] 张书娜.光照和施肥对林冠下珍贵阔叶树种人工更新的影响[D].哈尔滨:东北林业大学,2015.
[84] 许大全,高伟,阮军.光质对植物生长发育的影响[J].植物生理学报,2015,51(8):1217-1234.
[85] 马光恕,廉华,闫明伟.不同覆盖材料对大棚内番茄生长发育的影响[J].吉林农业科学,2002,27(4):41-43.
[86] 龙作义,刘汉平,吴全德.光周期对红皮云杉苗木的影响[J].牡丹江师范学院学报(自然科学版),1999,(1):12-13.
[87] 梁晓东,叶万辉.林窗研究进展(综述)[J].热带亚热带植物学报,2001,9(4):355-364.
[88] 刘珂艺,王庆贵,邢亚娟.森林动态演替模型的研究进展[J].中国农学通报,2018,34(13):43-49.
[89] Canham C D,Finzi A C,Pacala S W,et al.Causes and consequences of resource heterogeneity in forests:interspecic variation in light transmission by canopy trees[J].Canadian Journal of Forest Research,1994,24(2):337-349.
[90] Mark E S.The forgotten stage of forest succession:early-successional ecosystems on forest sites[J].Frontiers in Ecology and the Environment,2011,9(2):117-125.
[91] 郑芬.南岭小坑常绿阔叶林林下光环境变化及对幼树功能性状的影响[D].北京:中国林业科学研究院,2020.
[92] 杨期和,宋松泉,叶万辉,等.种子感光的机理及影响种子感光性的因素[J].植物学通报,2003,20(2):238-247.
[93] Chen Y.Macrofungal species distributions depend on habitat partitioning of topography,light,and vegetation in a temperate mountain forest[J].Scientific Reports,2018,8(1):13589-13602.
[94] 暴家兵.小尺度下光環境不均一性对微生物群落特征的影响[D].太原:山西大学,2020.
[95] 刘延滨,王庆成,王承义,等.退化落叶松人工林近自然化改造对土壤微生物及养分的影响[J].生态学杂志,2012,31(11):2716-2722.
[96] Gossner,Martin M.Light intensity affects spatial distribution of Heteroptera in deciduous forests[J].European Journal of Entomology,2009,106(2):241-252.
[97] Sverdrup-Thygeson A,Ims R A.The effect of forest clearcutting in Norway on the community of saproxylic beetles on aspen[J].Biological Conservation,2002,108(3):377-378.
[98] Henry S P.Absence of microclimate selectivity in insectivorous birds of the Neotropical forest understory[J].Biological Conservation,2015,188:116-125.
[99] Patten M A,Smith-Patten B D.Testing the microclimate hypothesis:light environment and population trends of Neotropical birds[J].Biological Conservation,2012,155:85-93.
[100] Zavala M A,Angulo O, Parra R B, et al.An analytical model of stand dynamics as a function of tree growth,mortality and recruitment:the shade tolerance-stand structure hypothesis revisited[J].Journal of Theoretical Biology,2007,244(3):440-450.
Characteristics of Forest Light Environment and Its Ecological Effects
Jin Peng, Xu Ming, Ran Fuju, Lei Yun,Meng Chuncan, Zhang Jian*
(College of Life Sciences,Guizhou University,Guiyang,Guizhou 550025,China)
Abstract:
Light is a key energy input to forest ecosystems.The evolution mechanisms of light environment in forest are complex.We summarize the light environment indicators and monitoring methods at the forest edge,forest interior and forest window,and also summarize their daily,seasonal and interannual changes based on analyzing the characteristics of light environment in forest on a temporal scale.At the same time,the research progress of the forest light environment at home and abroad were reviewed from vertical and horizontal gradient.It also explores the ecological role of the forest light environment in terms of individuals,communities and ecosystems,with the aim of providing a reference for understanding the synergistic mechanisms between the spatial and temporal dynamics of forest vegetation and the light environment,and providing a basis for promoting sustainable forest management.
Keywords:
forest light environmental; light environmental characteristics; temporal and spatial variations; ecological function
