海洋超微型蓝细菌聚球藻的生态学研究进展
2023-06-13贺博闻史文卿林大晖
郑 强,贺博闻,史文卿,陈 奇,林大晖,王 煜,2
(1.厦门大学海洋与地球学院,近海海洋环境科学国家重点实验室,福建省海洋碳汇重点实验室,福建 厦门 361102;2.厦门大学环境与生态学院,福建 厦门 361102)
聚球藻(Synechococcus)是地球上数量最多的原核光合自养生物之一(图1),在上层海洋中其丰度为102~106mL-1[1].聚球藻在全球范围内分布广泛,其个体微小(0.6~2 μm)[2],却占据了海洋浮游植物总生物量的12%~15%[3].聚球藻、原绿球藻(Prochlorococcus)及束毛藻(Trichodesmium)构成了海洋中最古老的原核光合自养生物类群——蓝细菌(cyanobacteria)[4].最早的蓝细菌于35亿~33亿年前的地质化石中被发现[5],约在27亿年前开始不断进化光合产氧能力[6].蓝细菌光合系统相关基因的进化分析表明,包括聚球藻在内的蓝细菌主要类群可能均由其近源类群黏杆菌藻属(Gloeobacter)分化而来[7].在后续的进化过程中,聚球藻产生了承载光合作用的重要生物膜结构——类囊体,同时也演变出更为复杂的藻胆体[8-9],使它们具备更强大的光合作用能力.聚球藻进而成为海洋中重要的初级生产者,与原绿球藻共同贡献了约25%的海洋净初级生产力[10].
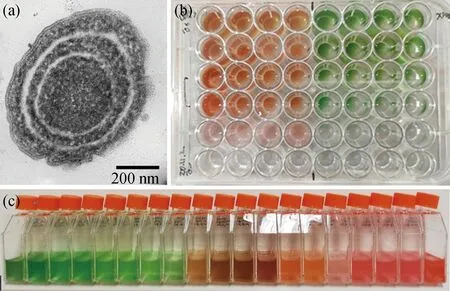
(a)透射电镜成像;(b)48孔板稀释纯化;(c)不同色素聚球藻培养体系,从左至右依次为聚球藻CBW1002、PCC7002、PCC7003、CCMP1333、XM3、XM5、XM24、SYN、CBW1106、CBW1107、CBW1005、XM11、YX04-1、YX04-3、YX04-4、YX02-1、WH7803、WH8012、YX04-2(藻蓝蛋白/藻红蛋白比例由高到低).图1 聚球藻的形态及不同色素聚球藻培养体系Fig.1 The morphology of Synechococcus and their cultures containing different pigments
1979年科学家发现聚球藻在海洋中广泛存在,此后聚球藻逐渐成为科学家研究的热点[2,11].伴随着克隆文库、聚合酶链式反应(PCR)、高通量测序、蛋白质组和单细胞测序等分子生物学手段的发展,聚球藻的分布范围、系统发育分类、生理特性等不断被揭示,其生态价值与贡献也逐渐被人们认可和重视.同时,在当前全球变化的影响下,海洋环境不断发生变化,聚球藻的生理活动及其对生态系统的贡献也将随之发生改变.本文从聚球藻对初级生产力的贡献、遗传多样性,及其与异养细菌、蓝细菌病毒的互作机制4个方面对相关研究进展进行总结,同时在全球变化大背景下对聚球藻在海洋中的生态贡献及生态意义进行展望,以期为未来聚球藻研究提供一定的理论参考.
1 聚球藻的生态意义
1.1 聚球藻对初级生产力的贡献
聚球藻作为海洋中重要的光合自养生物,可通过藻胆素、叶绿素a等多种色素进行光合作用,对海洋碳固定有重要贡献[12-13].在热带和亚热带东北大西洋的研究中发现,聚球藻细胞的固碳速率为(9.5±4.3) fg/h,其碳固定量可达该海域总碳固定量的(21±13)%[14],与近岸海域聚球藻对海洋的碳固定贡献率(>20%)相当[15].即使从净初级生产力角度估算,聚球藻仍可占全球海洋净初级生产力的16.7%左右,这足以表明它们在海洋碳固定中发挥着举足轻重的作用.此外,在全球变化背景下,生态模型结果表明聚球藻可能在全球海洋中占据更广阔的地理分布范围,进而在未来海洋中贡献更高的初级生产力[10].Flombaum等[10]以跨越35年遍及太平洋、南大西洋和南印度洋的庞大数据集为基础,建立了定量生态位模型,预测到21世纪末聚球藻的全球细胞丰度将增加14%.在有关2100年原绿球藻、聚球藻和超微型真核浮游植物分布变化的研究中,除聚球藻丰度的上升外,还预测超微型真核浮游植物丰度将下降8%[16].这表明聚球藻对海洋初级生产力贡献的增加并不单纯来自细胞数量增多,竞争性群落演替也是聚球藻贡献潜能提升的重要因素.聚球藻及其他蓝细菌类群(如束毛藻)同时可进行生物固氮,尤其在寡营养海域的新生氮供给中发挥重要作用[17-18].因此,未来聚球藻对海洋初级生产力及其中的物质元素循环将会有更大贡献.
1.2 聚球藻在微食物环中的作用
浮游植物是微食物环的重要物质基础,聚球藻自身固定的有机碳可被其他营养级生物分解或再矿化,进而推动海洋中碳的周转流动[19](图2).聚球藻固定CO2合成的有机物质部分会释放到周围环境,异养细菌可利用转化这些有机质为自身生物量,供给细胞生长繁殖,同时还可再矿化有机碳为CO2,使其再次被聚球藻等光合自养生物利用或直接以CO2形式返回到大气中[13,21].此外,聚球藻及异养细菌又可被鞭毛虫、纤毛虫等原生动物摄食,继而被中型浮游动物(如桡足类)消费,最后被鱼类等海洋生物捕食,从而进入经典食物链(网)[22].有机碳及能量在此过程中层层传递,被转化为生物量,或通过呼吸代谢消耗,或作为排泄物被分解者利用[23].

DOM.溶解有机质;POM.颗粒有机质;RDOM.惰性DOM.图2 聚球藻在海洋微生物环中的作用(参考文献[20],有修改)Fig.2 The roles of Synechococcus in the marine microbial loop (modified from Ref.[20])
1.3 聚球藻的海洋碳沉降
聚球藻作为海洋生物碳泵的重要参与者,一部分细胞会以直接或间接方式沉降到深海[24-25](图2).直接沉降途径主要指聚球藻等浮游植物产生透明胞外多糖颗粒,促进细胞黏合聚集,进而直接下沉至深海[26-27].尽管聚球藻个体微小,但在大西洋百慕大时间序列研究中发现,它们的聚集体占沉积物捕集器中颗粒有机碳总量的百分比可达2%~13%[28].此外,在间接沉降方面,多项关于深海碳通量的研究均明确表明,聚球藻存在于深海沉积物及深海消费者肠道中,证实了聚球藻经浮游动物摄食向海洋深层的输送[28-30].通过直接和间接沉降,聚球藻一般以与上层海洋净初级生产总量成正比的量级关系向下输出,因此其对海洋碳沉降的贡献不容忽视[24,31].
2 聚球藻的遗传多样性
2.1 聚球藻的全球丰度
聚球藻广泛分布于开阔大洋、近岸、河口等海域[32].聚球藻的全球海洋年平均总丰度约7×1026mL-1[10],在不同海域中常见丰度约103~105mL-1[33-34].在西太平洋沿海,聚球藻年际丰度变化为500~56 000 mL-1,数值在夏季达到最高,而冬季相对较低[35].这种季节丰度变化规律与以往诸多研究一致[36-38],其背后机制主要为聚球藻(相较于真核藻类而言)具有较高的温度适应和低浓度营养摄取能力[39-40].
在一些极度寡营养的地中海海域,聚球藻以约4×105mL-1的平均高丰度成为浮游植物的主导类群[41].在大西洋和印度洋海域聚球藻同样也有着较高的丰度,大西洋海域的聚球藻丰度为103~104mL-1[42-43],而在印度洋海域其最高丰度可达2×105mL-1以上[44].在营养盐含量较高的河口或泻湖区,聚球藻展现出比大洋更高的丰度.例如,Wang等[45]研究发现,切萨皮克湾的聚球藻丰度可达106mL-1,贡献了总浮游植物叶绿素a浓度的20%~40%.此外在地中海近岸地区的高盐泻湖中,Mercado等[46]同样发现最高达6×106mL-1的高丰度聚球藻.
2.2 聚球藻的分类及分布
随着聚球藻逐渐成为科学家的研究热点,其分类不断被完善和细化.以藻胆蛋白种类和组成作为分类标准,海洋聚球藻可分为藻蓝蛋白(1型)、藻红蛋白Ⅰ(2型)和藻红蛋白Ⅱ(3型);同时部分含有藻红蛋白Ⅰ和Ⅱ的株系可进一步基于其发色团上藻尿胆素 (phycourobilin,PUB)与藻红素(phycoerythrobilin,PEB)的比例,由低到高细分为3a~3c和3f亚型,而依据光质改变PUB与PEB比例的特殊株系则归为3d亚型[47-48].
以细胞光合色素组成、运动能力、鸟嘌呤+胞嘧啶(G+C)含量及嗜盐性进行综合分类时,聚球藻又可被分为5.1、5.2和5.3亚群,其中5.1亚群基于系统发育分析进一步分为5.1A和5.1B两大亚类[49].在此基础上,基于分子遗传特征,各亚群(类)还可通过不同分子标记进一步细分为多个分支.最为常用的分子标记为16SrRNA基因及分辨率更高的16SrDNA和23SrDNA基因之间的转录间隔(internally transcribed spacer,ITS)序列[50-51],基于ITS序列分类的具体分支情况见表1.此外,硝酸盐还原酶基因(nitrate reductase,narB)[53]、光合系统Ⅰ基因(photosystem Ⅰ gene,psbA)[54]、RNA聚合酶C1基因(RNA polymerase,rpoC1)[48]、藻红蛋白Ⅰ编码基因(phycoerythrin gene,cpeBA)[55]和细胞色素B6基因(cytochrome B6 subunit,petB)[56]等序列变异性更高的功能基因也常被用于辅助分类,以获得更精细的分类(支)结果.然而,功能基因相较于常用的分子标记(16SrDNA基因或ITS序列)难以覆盖全部聚球藻株系[32,57],这对分类结果完整性的影响不容忽视.

表1 基于ITS序列的聚球藻分类(支)
目前,聚球藻的分类工作仍在不断更新.2020年Salazar等[58]依据聚球藻基因组特征创建了一种新的分类系统,以科属等级分类替代上述的亚群分支.尽管该系统目前还没有被广泛使用,但聚球藻未来的分类方式仍值得期待.
聚球藻的生长范围纵贯赤道至两极,遍及近岸到大洋的真光层.5.1亚群在聚球藻中占据优势地位,其主要分支Ⅰ~Ⅳ在全球范围的分布特征已得到普遍认可:分支Ⅰ和Ⅳ常共同出现于高纬度近海或温带中营养水域,纬度约在30° N以北和30° S以南;分支Ⅱ则以较高的丰度广泛分布于更为温暖且寡营养的亚热带或热带水域,纬度在30° N~30° S之间;分支Ⅲ更偏向于生长在寡营养水域,但无明显的纬度偏好[32,44,52,59].而5.1亚群其余分支的分布情况也在一些文献中有所总结,例如:分支Ⅴ、Ⅵ、Ⅶ广泛分布在热带和亚热带水域[43-44];分支Ⅷ更适应于高盐水域[49,60];分支Ⅸ和Ⅹ则较为少见且丰度较低[43-44].相较于广泛分布的5.1亚群,5.2亚群主要集中分布在河口地区,如切萨皮克湾[61].5.3亚群则零散分布于地中海、东中国海及马尾藻海等海域[49,62-63].
需要强调的是,目前尚未有研究表明聚球藻的遗传多样性与其生理特性间存在明确对应关系.因此,聚球藻各类群的地理分布不能完全依靠其从属分类(支)判定,具体的环境条件和生态因素也可能塑造聚球藻的生理特性,进而限制其分布范围[38,64].如何厘清聚球藻生理特性及遗传特性间错综复杂的联系,将是未来亟待解决的问题之一.
2.3 聚球藻的基因组特性
海洋聚球藻的基因组平均约为2.62 Mbp,一般由一个环状染色体组成,与其他海洋细菌基因组相比较小[32].2003年首个海洋聚球藻基因组被成功测序(分离自热带大西洋海域的聚球藻菌株Synechococcussp. WH8102),其基因组大小为2.43 Mbp,预测到2 778个蛋白编码基因[65-66].截至2022年2月,NCBI数据库(http:∥www.ncbi.nlm.nih.gov)中已测序的聚球藻目(Synechococcales)下基因组共有605个,主要集中在原绿球藻科(Prochlorococcaceae)(238个基因组)与聚球藻科(Synechococcaceae)(286个基因组).截至2021年11月,Integrated Microbial Genomes &Microbiomes(IMG/M)数据库(http:∥img.jpi.doe.gov/m/)中已有1 068个聚球藻目相关的基因组数据[67].随着测序技术的不断发展,越来越多的海洋聚球藻基因组被测序,如图3所示为聚球藻XM-24全基因组图谱(本课题组未发表数据),海洋聚球藻对其所处环境条件的适应机制也被不断探究和挖掘[68].

图3 聚球藻XM-24全基因组图谱Fig.3 whole genome map of Synechococcus sp. XM-24
在基因组基本特征方面,基于较为完整的基因组数据分析,海洋蓝细菌类群的核心基因约为911个,泛基因数量可达27 376个以上[55].其中,聚球藻基因数量通常在2 840个左右,其平均G+C含量(58.25%)高于近缘种类原绿球藻(41.46%)[55].在基因组结构和组成上,海洋聚球藻独特于其他不同环境分离的蓝细菌类群.2003年Palenik等[66]对海洋聚球藻菌株WH8102、两株原绿球藻和一株淡水聚球藻进行了最早的海洋聚球藻比较基因组分析,发现海洋聚球藻基因组特征与其余株系有明显区别,这些差别主要来源于其对所在海洋环境的适应.类似地,2016年对24株来自不同水域聚球藻菌株的基因组研究中,Coutinho等[69]基于聚球藻菌株共有的同源基因进行系统发育重建,发现聚球藻属内部菌株基因多样性十分丰富,并强调淡水株系和海洋株系应基于基因组成进一步划分.2021年Chen等[70]比较了包括聚球藻、原绿球藻以及其他蓝藻在内的650株蓝细菌基因组,正式依据陆生、淡水和海洋3种生境对其进行划分,发现蓝藻可以通过其泛基因组中数百个光照、营养条件、盐度等特定功能基因表达以及水平基因转移,获得新的功能基因的方式以适应不同的环境.这种生境适应途径在多项研究中也得到证实[68,71].
不同生态型的海洋聚球藻类群具有各异的环境适应策略.2006年Palenik等[72]将分离自沿岸海域的聚球藻Synechococcussp. CC9311,与开放大洋海域的聚球藻WH8102进行比较基因组研究,发现二者通过较大差异的基因组成来适应各自所处环境,尤其体现在基因调控、转运和代谢、细胞运动能力以及光适应方面.例如在信号传导方面,开放大洋海域的聚球藻菌株WH8102的双组分信号转导系统十分有限,仅包括5个组氨酸激酶传感器和9个反应调节器,对应于其相对稳定的生态系统;相比之下,近岸海域的海洋聚球藻CC9311有更多的信号转导系统相关基因(11个组氨酸激酶和17个反应调节因子),以适应沿海更为多变的环境条件;不同进化支的海洋聚球藻同样可基于基因多样性适应不同的生态位,出现在开放大洋的聚球藻属进化支Ⅱ、Ⅲ、Ⅳ只有较少数量的调节系统,而另外一些进化支Ⅰ、Ⅴ、Ⅵ、Ⅷ、Ⅸ和5.2亚群的聚球藻则具有更多基因以应对多变的环境条件.综上可见,水平基因转移可以使聚球藻获得特殊功能基因,而环境压力的选择作用使之保留下来,从而利于不同菌株适应其独特生境.
3 聚球藻与其共栖异养细菌相互作用
光合自养与异养微生物的相互作用关系在海洋微食物环以及生物地球化学循环中起着重要作用[20,73].实验室分离的聚球藻培养体系中往往含有大量的异养细菌,利用流式细胞细菌分选聚球藻再培养,或加入抗生素抑制异养细菌的生长,均只能在短时间内将异养细菌控制在一个低丰度水平;随着聚球藻的生长和抗生素作用的去除,异养细菌会再次快速生长.这些异养细菌是随聚球藻分离和生长过程而自然选择下来与聚球藻共栖的异养微生物,也是研究光合自养与异养微生物群落互作关系的良好实验对象.聚球藻与异养细菌之间存在着密切的协同作用关系,几种具有不同过氧化氢酶活性的异养细菌均能够支持低初始密度聚球藻在固体培养基上的生长[74].与无菌(低浓度异养细菌)培养相比,聚球藻与异养细菌共培养能够使其生长得更好,存活时间更久[75].
不同生态型的聚球藻与多种关系密切的异养细菌间有着复杂的互作关系.前期本课题组研究发现,富营养生态型Synechococcussp. XM-24和寡营养生态型Synechococcussp. YX04-3与共栖异养细菌培养显示出不同的动态关系[21].在寡营养聚球藻培养体系中,可能是由于聚球藻释放了更多溶解有机物,大部分优势异养细菌类群更倾向于游离的生活方式;并且在聚球藻指数生长期,异养细菌丰度可高于聚球藻丰度[76].
通过分析富营养和寡营养生态型的聚球藻培养体系,发现聚球藻共栖的优势异养细菌类群是相似的,主要来源于CFB(Cytophaga-Flavobaeteria-Baeteriodes)菌群、α-变形杆菌(α-proteobacteria)、γ-变形杆菌(γ-proteobacteria)以及放线菌(Actinobacteria)[21].聚球藻和异养细菌形成的关系网络可能因生境的不同而存在差异,共栖异养细菌群落在实验室条件下高度稳定[77].黄杆菌属(Flavobacteria)通常是聚球藻共培养体系中最优势的类群,一般倾向于附着生活,善于降解复杂高分子有机物或生物聚合物[78-80].对存在于聚球藻培养体系的黄杆菌进行基因组分析,发现其中包含多种聚合物降解酶基因及与滑行运动和黏附聚集相关的基因,表征了其附着能力和多糖降解及复杂蛋白质水解利用的潜能[76,80-81].玫瑰杆菌属(Roseobacter)也是共栖体系中的优势类群,它们有着游离和附着两种生活方式,既能利用高分子量有机物,实现高分子量向低分子量有机物转化,又能够利用聚球藻和黄杆菌产生的低分子量代谢副产物,有着较强的环境适应能力[75,79-80].
聚球藻与共栖异养细菌培养中,二者在营养物质循环与利用、应对氧化压力以及维生素B12(vitamin B12,VB12)合成等方面形成协同作用关系[76,80].缺失清除活性氧功能或VB12合成能力的类群依赖于具有这些功能的微生物类群,共同维持稳定的共栖体系[76,80,82].共栖体系中也存在着拮抗的相互作用,例如玫瑰杆菌Marivitasp. XM-24在聚球藻生长后期可能与其竞争磷酸盐,拟杆菌Roseivirgasp. XM-24会释放一些具有杀藻活性的蛋白,可能加速聚球藻死亡[80].而当将环境微生物群落接种于呈指数增长的聚球藻培养体系进行长期培养时,聚球藻和异养细菌之间还会经历从拮抗到互利共生的关系转变[83].
聚球藻释放至环境中的有机物质控制着培养体系中异养细菌的生活策略和群落演替,同时异养细菌代谢利用有机质存在一定的顺序[20-21,79].聚球藻将一部分光合产物释放至环境中,黄杆菌能够将其中高分子量有机物代谢转化为低分子量有机物,α-变形杆菌进一步将低分子有机物代谢转化为无机营养盐供聚球藻再利用,从而在共培养体系中形成一种协同作用的闭合回路(图4),维持聚球藻培养体系的长期稳定[84].

PSⅠ.光系统Ⅰ;PSⅡ.光系统Ⅱ;NADPH.还原型辅酶Ⅱ;ATP.三磷酸腺苷;ADP.二磷酸腺苷;SOD.超氧化物歧化酶;SusCD.参与生物聚合物结合和摄取的蛋白复合体;SwmAB.细胞运动表面蛋白;PUL.普鲁兰多糖;PHA.聚羟基脂肪酸酯.图4 基于蛋白质数据的聚球藻(a)与共栖异养细菌黄杆菌(b)及α-变形杆菌(c)相互作用(参考文献[76],有修改)Fig.4 Interactions between Synechococcus (a),and associated heterotrophic bacteria Flavobacteria (b) and α-proteobacteria (c) based on the metaproteomic data (modified from Ref.[76])
4 聚球藻病毒
侵染聚球藻和原绿球藻等蓝细菌的病毒被称作噬藻体或蓝细菌病毒(cyanophage),是海洋噬菌体的重要组成类群之一,在微生物群落结构调节及海洋生物地球化学循环中发挥重要作用[85-86].病毒侵染往往被认为是海洋蓝细菌死亡的重要途径,但侵染率和死亡率的研究估算范围较大.早期研究表明每天10%~30%的海洋蓝细菌被病毒裂解,而最近通过“iPolony”方法估计原绿球藻的病毒感染率普遍较低,仅造成原绿球藻每天0.35%~4.8%的死亡率[85,87].尽管死亡率较低,但由于蓝细菌在海洋中数量巨大,蓝细菌病毒裂解所释放的有机质及无机营养盐仍对海洋C、N、P等化学元素循环具有重要影响.
4.1 聚球藻病毒特征及分类
与所有噬菌体相同,蓝细菌病毒由蛋白质外壳包裹核酸,营严格胞内寄生生活,通过感染宿主细胞进行繁殖,依据其形态和生活史等特征可分为多种类型(图5).根据其生活史可分为裂解性噬菌体和溶原性噬菌体,不同生活策略反映噬菌体对于环境的适应性,通常在环境中有较高宿主丰度时噬菌体倾向于裂解性生活,而较低宿主丰度时则倾向于溶原性噬菌体[85,88].根据噬菌体可侵染的宿主范围和裂解量可分为低裂解量宽泛宿主范围和高裂解量狭窄宿主范围两类[89].目前所发现的蓝细菌病毒均为有尾双链DNA类型,从海洋聚球藻中分离的病毒多为宿主范围较广的肌尾噬藻体,从河口区域的聚球藻中分离的短尾和长尾噬藻体则有较高的宿主专一性[90].此外,较为“温和”的前噬藻体目前仅在淡水蓝细菌株系中被诱导出[91],而侵染海洋聚球藻的病毒主要是具有较强侵染性和高裂解量的裂解性病毒[90,92].

图5 聚球藻病毒的形态Fig.5 Morphology of Synechococcus virus
4.2 聚球藻病毒和宿主的相互作用
海洋中蓝细菌的丰度、生理状况以及昼夜变化等因素会影响蓝细菌病毒的丰度[93-94].在自然环境中,聚球藻等蓝细菌在基因表达和代谢等方面表现出昼夜节律以适应光暗变化周期[95-96].可能由于对宿主细胞的吸附以及对宿主光合作用能量的依赖等,蓝细菌病毒侵染宿主过程也表现出昼夜节律[95,97-98].蓝细菌病毒中存在周期性表达的基因,并在海洋表层侵染宿主具有同步性,可导致溶解有机物的同步释放[88,99].
病毒裂解宿主细胞将大量营养元素(C、N、P、S、Fe等)释放进入微食物环(图2),继而通过异养细菌的代谢转化将其中一部分向更高营养级传递,即病毒回路,该过程中同时会产生难以被生物利用的RDOM,进而贡献于海洋RDOM库[100-101].随着光谱、质谱和波谱等技术的应用,研究人员发现病毒裂解聚球藻产物中包含大量荧光类DOM(fluorescent DOM,FDOM),一部分相对活性的DOM能够被异养细菌进一步利用,另一部分则相对惰性,且与深海FDOM的荧光信号相似,可能是深海RDOM的前体物质[76,102].此外,蓝细菌病毒在裂解聚球藻时会释放大量含N和S的小分子物质,能够被特定的微生物类群优先利用[103].在聚球藻与异养细菌共培养体系中加入蓝细菌病毒,发现聚球藻裂解期间,共栖异养细菌可快速代谢聚球藻裂解释放的有机物,使培养体系内铵盐累积,而磷酸盐的再生累积较为滞后,说明含N和含P有机物参与不同的微生物代谢过程[75,104].
病毒与其宿主之间的相互作用还包括种群多样性的调控以及病毒辅助宿主代谢等.在环境中观察到聚球藻病毒与宿主群落变化具有一致性,病毒通过“Kill the Winner”机制维持宿主群落多样性[105-106],同时通过裂解蓝细菌释放出的产物影响其他微生物的群落结构与多样性,进而维持整个群落的生态平衡[80,104].此外,在病毒的基因组中还存在一些辅助代谢基因,能够调节宿主光合作用、碳代谢以及核苷酸合成等代谢途径并促进新病毒的产生[107-108].
5 总结与展望
本文归纳了聚球藻这一重要浮游植物类群的生态学意义、遗传多样性及其与共栖异养细菌和病毒的互作关系,阐释了聚球藻在全球海洋尺度碳循环中扮演的源头角色和后续与其他物种间的紧密联系.值得关注的是,近年来人类活动对全球变化产生重大影响[109],全球海洋环境正发生不容忽视的转变,聚球藻在未来海洋中的生态地位也将随之受到影响.
首先,CO2排放的增加扰乱了海洋碳酸盐缓冲体系,降低了海水pH,造成海洋酸化现象发生[110].此外,过量温室气体导致的温室效应将升高海洋表层温度,升温协同增加的河流和降水输入量加大海洋表层及深层密度差,加剧层化现象,阻碍氧气传递及营养物质的垂直向上输送,使得海洋部分区域出现缺氧现象,同时寡营养海域面积增加[111].环境条件的变化与生物生存息息相关[112],海洋生境动荡将引起聚球藻的多种生理响应,例如:适度的CO2浓度增加可加快细胞光合速率、下调碳浓缩机制,使更多能量用于聚球藻生长[113-114];而同时产生的海洋酸化胁迫却会带来细胞呼吸作用增强、藻类耐高光胁迫能力降低等负面影响[115];升温在聚球藻耐受限度内外分别发挥正负双向作用[116-117];而缺氧及营养盐降低也将不同程度地限制细胞的正常生长[118-119].但当受到多种环境因子的叠加影响时,聚球藻的生理响应会更为复杂且难以预料,因此目前尚无法对其未来具体生长、分布、类群演替情况及生态地位下定论[114,120].然而,多个时间序列观测和模型预测结果表明,聚球藻极可能通过丰度及生态位的改变在未来海洋中发挥更大的生态作用[10].尽管聚球藻的细胞体积小,但营养吸收快且代谢消耗低,比体积大的浮游生物具有更强的生存竞争优势[121],不仅可在全球变化下向高纬度地区扩张地理分布[10,122],甚至还可在寡营养海域取代硅藻等更大体积的浮游植物成为主要的海洋初级生产者,贡献更多海洋初级生产力[123].与此同时,海洋经典食物链(硅藻/甲藻→桡足类→鱼)或将向微型食物链(聚球藻→鞭毛虫/纤毛虫→桡足类→鱼)偏移[124].当营养价值相对较低的聚球藻成为主要的被摄食对象,后续摄食者的生长及群落结构会受到怎样的影响?海洋生物地球化学循环及能量流动又将因此发生怎样的变化?诸多相关的科学问题还等待进一步探索.可以预见,多环境因子叠加效应的生理、生态实验以及结合历史航次数据构建高可信度的生态模型将在此类难题破解中发挥重要作用.
