红曲霉ABC转运蛋白基因家族的鉴定与生物信息学分析
2023-06-07李燕萍
韩 尧,李燕萍*
(南昌大学a.食品科学与技术国家重点实验室;b.中德联合研究院;c.食品学院,江西 南昌 330047)
ABC转运蛋白(ATP-binding cassette transporters,ATP结合盒转运蛋白)是在生物界广泛存在的、能够催化ATP水解并利用水解所产生的能量促使底物实现跨膜转运的一类膜整合蛋白[1]。其底物种类包括糖、氨基酸、蛋白质和金属离子等多种生物分子,参与多种生物过程,如真菌的多药物外排、植物激素运输、信号转导、细胞解毒、病毒防御以及抗原传递[2]。ABC转运蛋白家族具有保守结构,典型结构是由2个核苷酸结合结构域(nucleotidebinding domain,NBD)和2个跨膜结构域(transmembrane domain,TMD)组成,其中NBD结构域相对保守,行使ATP水解酶功能,为跨膜运输提供能量,而TMD是跨膜通道,其结构及序列具有底物的特异性[3]。底物从细胞膜外向细胞内转运的过程如图1所示,具体如下:内向性ABC转运蛋白,先形成外周蛋白和底物的复合体(Substrate binding protein,SBP),而后与ABC转运蛋白相互作用,把底物传递给它的TMD部分。
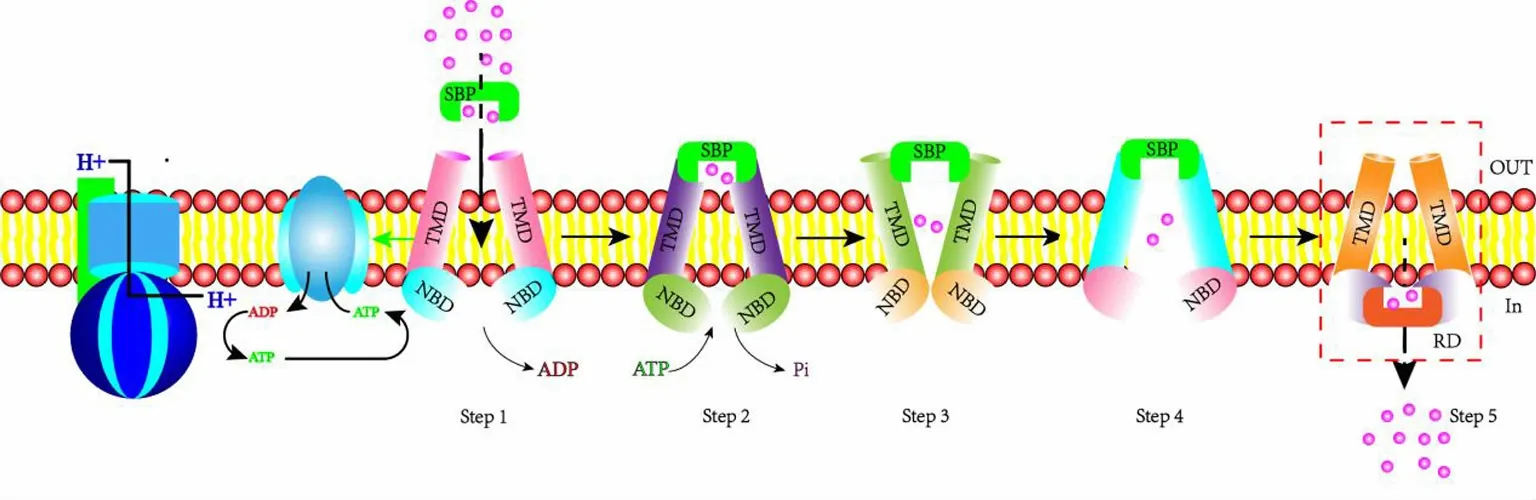
图1 内向 ABC 转运蛋白转运机理模型图[5] SBP (Substrate binding protein):底物结合蛋白;RD (Regulatory domain):调节区域;粉色空心小球代表底物Fig.1 Model of Substrate Binding Protein transport mechanism[5] SBP (Substrate Binding Protein); RD (Regulatory domain):Regulatory area.The pink hollow spheres represent the substrate
ABC转运蛋白的2个TMD间存在一个亲水性通道孔隙,在转运底物前,靠近膜外的通道口处于开放状态,靠近膜内的通道口处于关闭状态;在进行底物跨膜转运时,膜外结合蛋白结合底物后并将其传送给TMD,同时把信号传递到NBD,NBD在水解ATP后,其构象发生变化并将信号传递到TMD,TMD的构象亦发生变化;底物被送到通道孔隙中后,朝向外周质的通道口随即关闭,朝向胞质的通道口随即开放,底物进入细胞内。而底物从细胞内转运到细胞外时,除ABC转运蛋白上的内外通道口开放顺序相反外,其他转运过程都基本一致[4]。
ABC转运蛋白最初作为营养物质内运蛋白在细菌中被发现,随后因参与癌细胞的多药耐药性(Multidrug resistance,MDR)引起人们关注。真菌中ABC转运蛋白种类较多,功能各异。白色假丝酵母耐药性蛋白CDR1(Candida drug resistance)是第一个被报道的致病中的多向耐药性ABC转运蛋白[6]。CDR1的底物包括了结构上无关的唑类、脂类和类固醇,敲除白色假丝酵母中的CDR1基因导致其对所有检测的唑类和其他代谢抑制剂高度敏感性,而HEX 1启动子通过对CDR1基因增强性的过表达导致白色假丝酵母耐药性的增加[7]。产黄青霉(Penicilliumchrysogenum)中ABC转运蛋白编码基因ABC40主要参与苯乙酸的外排,提高菌株苯乙酸及其他弱酸的耐受力[8];指状青霉(Penicilliumdigitatum)中的ABC转运蛋白编码基因主要参与细胞中药物外排[9];BcatrB编码的灰霉菌(Botrytiscinerea)ABC转运蛋白主要负责亲脂类化合物的转运[10];在酵母体系表达烟曲霉PDR 5类ABC膜转运蛋白的研究表明,ABC膜转运蛋白的缺失导致烟曲霉[11]底物特异性和能量依赖性外排方面产生巨大的差异;红曲霉中囊泡转运蛋白Ypt7基因[12]缺失引起了ABC转运蛋白相关基因表达的变化。这说明不同的转运途径相互联系,相互影响。对于同一底物的运输,可能有不同运输途径可供选择。
红曲霉在我国生产使用已经有1000多年的历史,且红曲霉是目前世界上唯一被批准可用于生产食用色素的微生物[13]。因此,在食品发酵生产领域红曲霉具有重要的研究价值。红曲霉能够产生醇、酸、酯等多种芳香物质(初级代谢产物)和多种水解酶类如淀粉分解酶、蛋白质分解酶、半乳糖分解酶、核糖核酸分解酶等,使食品在发酵过程中产生优质香气和甘甜味道;而它的次级代谢产物如色素、抗菌素、胆固醇抑制剂、中药成分等更是近年来人们研究的热点[14]。越来越多的研究表明,ABC转运蛋白作为最大的初级主动转运蛋白,主要参与真菌氨基酸、糖类、维生素等物质的运输,但是对于红曲霉内ABC转运蛋白至今少有研究和明确的分类。本研究提取橙色红曲霉AS3.4384基因组DNA,经质量验证合格后进行二代建库测序(北京诺禾致源科技股份有限公司),并对测序得到的序列进行组装及注释。鉴定和分析了红曲霉AS3.4384中ABC转运蛋白基因家族成员数目;在本研究中,我们鉴定出33条红曲霉ABCs基因序列,分为7大亚类。从外显子-内含子组织、基序组成、序列片段分布、系统发育和同源性分析等方面进行了综合分析。构建了红曲霉ABC转运蛋白系统进化树,探讨了ABC转运蛋白基因家族成员间的进化关系。该研究结果为红曲霉ABC转运蛋白基因功能的研究提供了基础资料。
1 实验方法
1.1 红曲霉ABC转运蛋白家族成员的鉴定
利用黑曲霉、烟曲霉和酵母3种相似真菌ABC转运蛋白基因家族成员蛋白序列作为查询序列,通过本地BLAST比对程序搜索,以e值≤1e-5为搜索条件,从红曲霉蛋白序列集中鉴定ABC转运蛋白序列。将鉴定获得ABC蛋白序列提交CDD数库,(https//www.ncbi.nlm.nih.gov/Structure/bw-rpsb/bwrpsb.cgi),Pfam)[15]。进一步验证ABC转运蛋白的NBD和TMD保守结构域,不存在ABC蛋白保守结构域或者分子量不在50~300 kDa范围之内的蛋白序列被去除,最后所有非冗余高置信度序列均命名为红曲霉ABC转运蛋白(MsABC)。
1.2 红曲霉ABC转运蛋白序列分析和结构表征
所有高置信度的MsABC蛋白序列提交到Ex-PASy(http://web.expasy.org/protpara-m/)以计算氨基酸数量、分子量和理论等电点(PI)。MEME程序(版本4.11.2,http://alte-rnate.meme-suite.org/tools/meme)用于识别MsABCs序列中的保守基序,参数如下:任意重复次数,最多10个错配,最佳基序宽度为6~100个氨基酸残基。在基因结构显示服务器(GSDS,http://gsds.cbi.pku.edu.cn/)上对MsABCs基因的外显子-内含子结构进行了鉴定[16]。
1.3 红曲霉ABC转运蛋白的系统发育分析和分类
利用来自黑曲霉、烟曲霉和酵母的ABC转运蛋白的全长序列(附加文件1)和新鉴定的MsABCs进行了系统发育分析。所有获得的序列首先由ClustatX[17](版本1.83)软件用默认参数进行比对,然后使用MEGA 6[18]软件构建了一个无根邻接系统发育树,并进行了1 000次的自举检验。根据系统发育树的拓扑结构和上述3个物种的ABC转运蛋白的分类,将红曲霉ABC转运蛋白分为不同的亚类。
1.4 红曲霉ABC基因启动子顺式作用元件分析
红曲霉的生长发育受到光照、营养物质、温度、PH和水分等胁迫作用的严重影响,导致红曲色素等代谢产物降低,品质下降。红曲霉受到逆境胁迫时会产生相应的应答反应,以降低或消除逆境胁迫给生长带来的危害。红曲霉的这种应答反应是一个涉及多基因、多信号途径及多基因产物的复杂过程。首先是在转录水平上调相应基因的表达,转录水平上的调控是由启动子和与之相互作用的转录因子共同完成的。因此将MsABCs编码序列的上游启动子序列(1.5 kb)提交到PlantCare[19](https://bioinformatics.psb.ugent.be/WebTools/plantcare/html/)以鉴定参与脱落酸响应元件(ABA-Responsive Element,ABA)、参与抗氧化的响应元件(Antioxidant Response Element,ARE)、参与脱水、低温和盐胁迫的脱水响应元件(Dehydration Responsive Element,DRE)、参与生长素的响应元件(IAA-Responsive Element,IAA)、参与低温响应元件(Low temperature Response Element,LTRE)、参与茉莉酸甲酯响应元件(Methyl Jasmonate,MeJA)等6个响应元件。
1.5 红曲霉ABC基因的序列分布及同源性分析
将MsABCs编码基因所在的基因全片段序列导入Mauve软件[20],添加需要比对的基因序列,选择全基因组序列比对,设置默认参数,比对完成后获得基因组位点间的线性关系。将分析获得相关性基因序列使用TBtools软件[21]绘制基因内共线性图谱。
2 结果
2.1 红曲霉ABC转运蛋白的鉴定与分析
通过本地BLASTP比对获得36条序列,通过关键词搜索在NCBI数据库获得5条序列。(红曲霉ABC转运蛋白研究较少,大多数未进行标注。)去除重复序列后,保留39条序列,提交给CDD(C-onserved Domain Database)数据库(https://www.ncbi.nlm.nih.gov/cdd)来确认ACD(Actin CrossLinking Domain)结构域。排除没有典型ACD和分子量在50~300 kDa范围之外的序列。最终确定33条红曲霉ABC转运蛋白序列,并根据其亚类进行命名,表1列出了MsABCs蛋白的序列名称、序列ID、序列位置、开放阅读框长度、外显子数量、氨基酸数量、分子量和等电点。MsABCs蛋白的长度从617个氨基酸(MsABC-31)到2600个氨基酸(MsABC-32)不等。MsABCs蛋白的分子量介于69.2 kDa(MsABC-31)~291.6 kDa(MsABC-32)之间。MsABCs基因分布在22条红曲霉序列片段上。MsABCs蛋白的预测等电点为5.30(MsABC-11)~9.23(MsABC-12)。
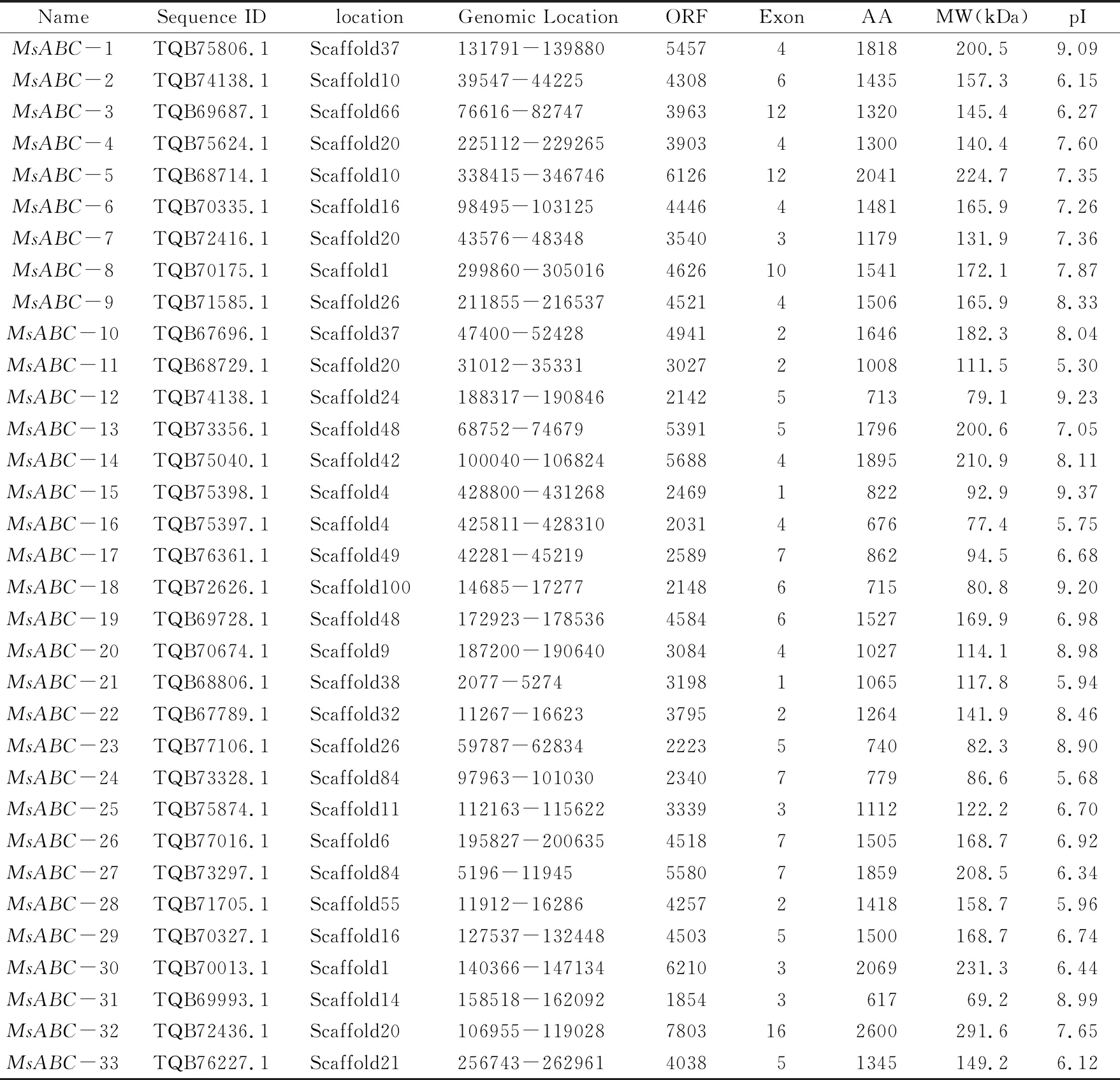
表1 红曲霉 ABCs 转运蛋白结构信息Tab.1 Structure of Monascus ABCs transporter
2.2 红曲霉ABC基因结构
通过基因组DNA与MsABCs全长cDNA的比对,确定内含子/外显子的结构和相位。图2显示在MsABCs基因中,绝大多数MsABCs基因都含有内含子,只有MsABC-21不含内含子,大部分MsABCs基因含有2~7个内含子,只有4个基因含有10个以上内含子,即MsABC-3含有12个内含子、MsABC-5含有12个内含子、MsABC-8含有10个内含子、MsABC-32含有16个内含子。推断不含内含子为串联复制的基因,含有较多内含子的为片段复制的基因,红曲霉ABC转运蛋白多为分段复制基因。不过有趣的是,多条片段复制基因之间存在高度的保守区域,例如虽然MsABC-32序列比MsABC-3序列长,但是保守区具有相同的内含子相位,这一结果提示这两个片段重复基因之间存在着特殊的系统发育关系。
通过motif序列分析工具(Multiple EM for Motif Elicitation,MEME)对MsABCs的保守基序进行分析,发现了5个保守的motifs基序。这些保守基序的长度从21到50个氨基酸不等。表2列出了5个motifs序列的详细信息。根据CDD数据库比对分析,motif 3完全对应于保守的ACD区域。Motif 1、motif 2和motif 4的全序列一起形成了高度保守的完整ACD结构域。大多数MsABCs序列(60.6%)含有motif 1、motif 3和motif 4组合,推测它们功能与跨膜区域相关。MsABC-15、MsABC-16、和MsABC-31缺乏相应的基序组合,未能与其余类型的MsABCs序列共聚类。ACD结构域的不同组成表明转运蛋白功能多样性。系统发育树中的同一组MsABCs具有共同的基序,表明它们具有高度保守性。聚在同一亚家族的motif保守基序组成相似,说明成员之间进化关系较近。不同的亚家族之间motif保守基序存在差异,motif出现频率越高说明序列保守程度越高。
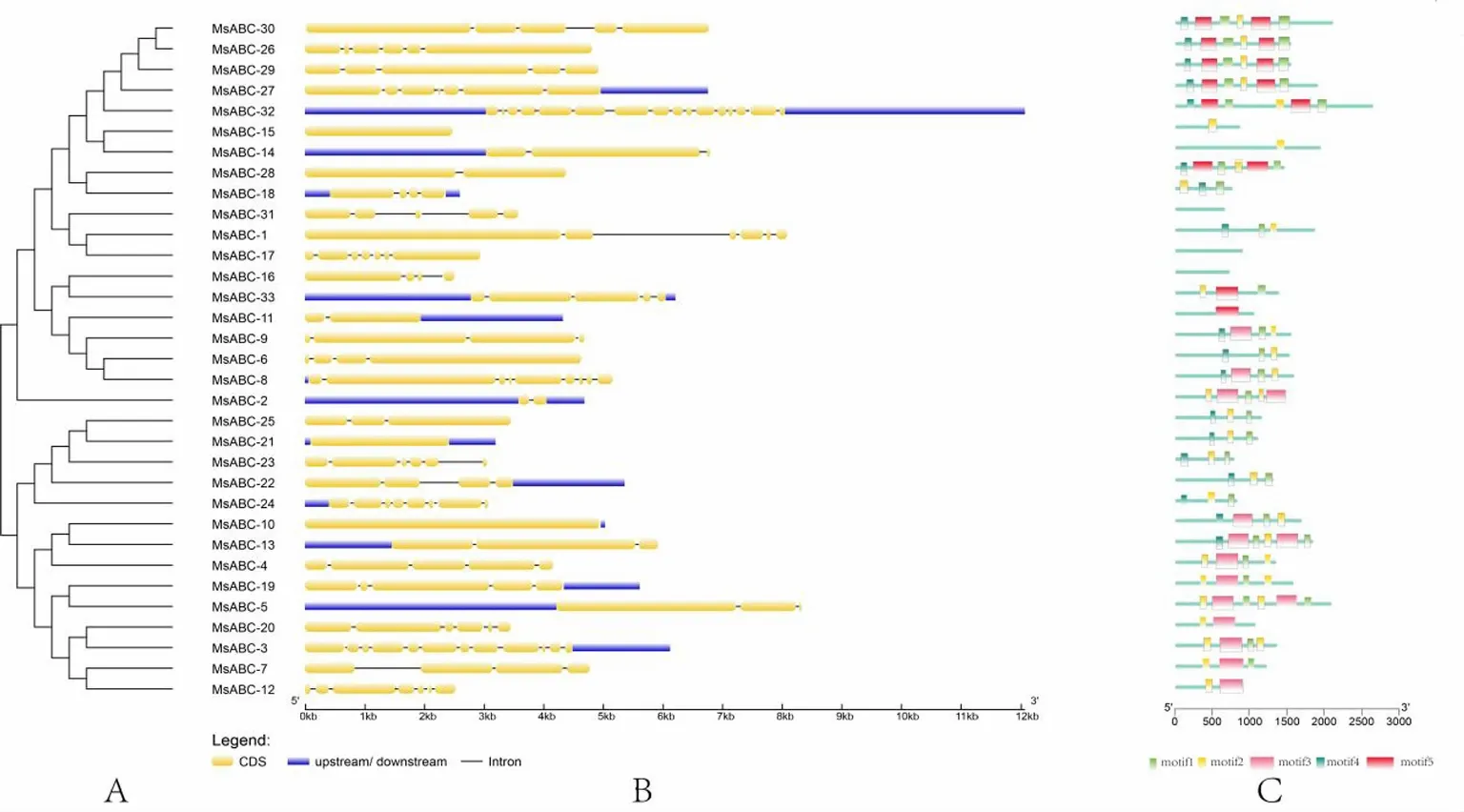
图2 MsABCs基因的系统发育关系(A)、基因结构(B)和保守基序分析(C)。MsABCs基因的上游/下游区域用蓝色方框表示。外显子的长度可以通过底部的刻度来推断;MsABCs基因保守基序的分布,不同颜色的方框中显示了5个不同的motifs区域。有关图案的详细信息,请参阅表2。

表2 红曲霉 ABCs 转运蛋白保守基序片段Tab.2 Conserved motif fragments of Monascus ABCs transporter
2.3 红曲霉ABC转运蛋白的系统发育分析
为了分析黑曲霉、烟曲霉、酵母和红曲霉中ABC转运蛋白的进化关系,利用黑曲霉、烟曲霉、酵母和红曲霉ABC转运蛋白序列,采用邻位连接法(Neighbor joing,NJ)构建无根环状系统发育树(图3)。在系统发育树中,共有来自黑曲霉的20条序列、烟曲霉的25条序列、酵母菌的43条序列和红曲霉的33条序列进行了系统发育树分析。121条ABC转运蛋白按照转运机制和功能的不同分为ABCA、ABCB、ABCC、ABCD、ABCE、ABCF和ABCG 7种亚类。根据邻位连接法(Neighbor joing,NJ)构建的系统发育树得出,33条红曲霉ABC转运蛋白序列很好地聚类到7种ABC转运蛋白亚类中。值得注意的是,MsABC亚家族除了与相同物种间成员聚类外,存在MsABC成员与来自不同物种的同一亚科的成员的亲缘关系比与来自同一物种的其他ABC成员的亲缘关系更密切,这意味着同一ABC亚家族除了在相同物种间具有较高的同源性外在不同物种之间同样存在较高的同源性,显示出ABC蛋白序列在进化过程中具有较高的保守性。
2.4 红曲霉ABC基因启动子序列中与胁迫相关的顺式调控元件
为了进一步研究MsABCs基因在非生物胁迫反应中的潜在调控机制,将MsABCs基因翻译起始位点上游1.5 kb序列提交到PlantCare以检测顺式作用元件。图4显示了6个非生物胁迫响应元件:脱落酸(ABA)响应元件、抗氧化(ARE)的响应元件、脱水(DRE)响应元件、应激生长(IAA)的响应元件、低温(LTRE)响应元件和茉莉酸甲酯(MeJA)相关的6个响应元件。每条MsABC基因启动子序列至少含有5个胁迫应答顺式元件,说明MsABCs基因的转录与这些非生物胁迫有关。33条MsABCs基因启动子序列有28条含有LTRE响应元件,这表明红曲菌在高温条件下可能存在热应激反应。另外几乎所有的MsABCs基因启动子序列含有ABA、ARE、LTRE、MeJA相关响应元件。总之顺式元件分析表明MsABCs基因可以响应多种环境的非生物胁迫。
2.5 红曲霉ABC基因序列分布及同源性分析
为进一步研究MsABCs基因在片段上分布和基因之间的相互联系,我们使用Mauve软件进行物种内基因共线性分析,该软件能够快速有效分析基因组之间有无大片段序列重排现象,是否存在共线性相关,是否存在LCBs(Locally Collinear Bloc-ks)局部共线区?如相关性较强,则说明基因组进化上可能经历着相似途径,若差异较大则说明在长期的选择压力下,通过在LCBs区域扩充新的基因来获得更好的生存空间,分期较早。Mauve软件包括MauveAligner和ProgressveMauve两个算法:
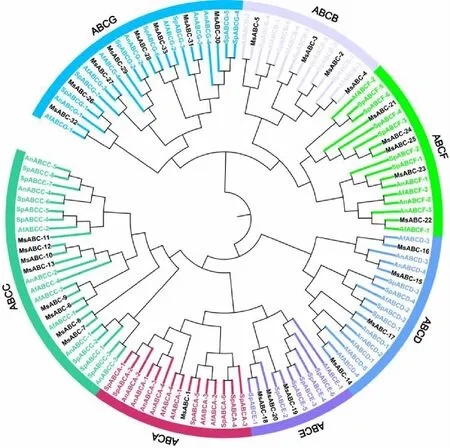
图3 黑曲霉、烟曲霉、酵母和红曲霉 ABC 转运蛋白的环状系统发育树(MsABC标注红曲霉、AnABC标注黑曲霉、AfABC标注烟曲霉、SpABC标注酵母)
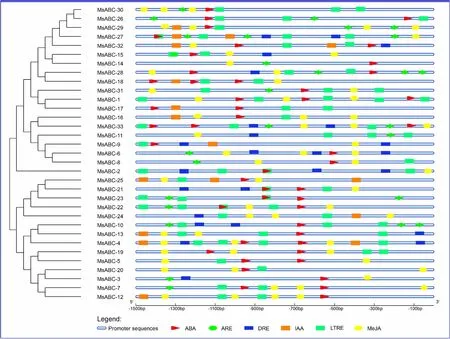
图4 预测了MsABC启动子中的顺式元件。使用PlantCare分析了33条MsABCs的启动子序列(-1500 bp)。可以根据底部的比例尺推断到翻译起始点的上游长度。
MaueAligner可以有效地识别多重基因组中的保守基因组区域,重新排列区域,保守区域中的插入序列和精确的断点等。此外还可以识别核苷酸替换,小片段的插入和缺失。但是MauveAligner算法只能识别在所有比对基因组中都存在的保守序列,存在于部分物种中的保守序列不能被识别,所以适合比对近源物种。ProgressiveMauve算法可以检测到只在部分基因组中存在的同源序列,可以比对分化距离较远的序列,多适用于物种间共线性分析。本课题研究红曲霉内同源性且ABC转运蛋白基因具有较高的保守性,因此采用MauveAligner算法进行物种内共线性分析。
图5显示,MsABCs转运蛋白基因分布在22条红曲霉序列Scaffolds片段上,Scaffold 20含有Ms-ABCs基因片段最多(4条),大部分Scaffolds含有1~2条MsABCs序列,Scaffolds序列长度与所含有MsABCs基因数目无正相关。根据Holub[22]的描述,包含两个或多个基因的200 kb以内的染色体区域被定义为串联重复基因,比对分析发现红曲霉连锁群含有12组串联重复基因。这些结果表明,部分MsABCs基因可能是由基因串联复制产生的。
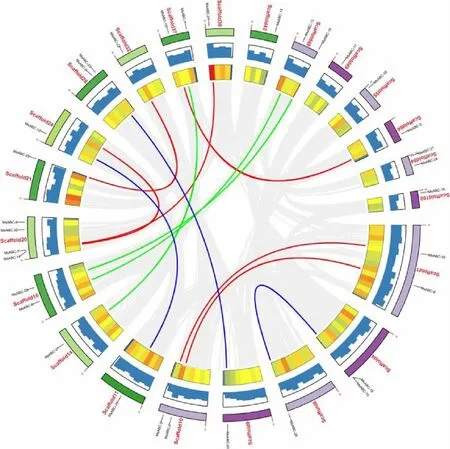
图5 MsABCs基因的分布和序列片段间关系示意图。灰色线条表示红曲霉基因组中所有的同步性区块,有色线条表示相关的MsABCs基因对
3 讨论
ABC转运蛋白超家族存在于从微生物到人类近3000多种生物体内,是生物界广泛存在的一类利用ATP水解提供能量促使底物跨膜转运的膜整合蛋白,包括内运蛋白(Importers)和外排蛋白(Exporters)。最近的临床医学研究表明,随着致病真菌深度感染的增加和抗真菌药物的大量使用,越来越多的抗真菌药物治疗效果明显下降,根本原因在于真菌产生了多药耐药性,而编码ABC转运蛋白基因的过度表达是致病真菌获得多药耐药性的主要原因之一[23]。越来越多致病真菌的ABC基因家族陆续被鉴定出来,如烟曲霉和白色念珠菌(Candidaalbicans)。但是,人们对红曲霉ABC基因家族却知之甚少。
本研究共鉴定出33条MsABCs转运蛋白基因,并对其结构、序列定位、系统发育、基因重复、胁迫相关顺式元件以及基因组内共线性进行了分析。这项研究对MsABCs基因家族的研究提供了全面的信息,将有助于理解红曲霉中MsABCs基因的功能差异。
基因的结构在多基因家族的进化中起着至关重要的作用[24]。在本研究中,含有1~3内含子MsABCs基因的比例(30.30%)与烟曲霉(35.71%)和黑曲霉(30.95%)相似。这种内含子数量和基序排列之间的相关性进一步证实了MsABCs基因的分类。在一些研究中,内含子很少或没有内含子的基因在物种中的表达水平被认为是增强的。为了及时应对各种环境压力,基因必须被迅速激活,内含子较少的紧凑基因有助于快速应对,这也是基因长期进化的一种内在表现。
本研究系统分析了红曲霉33条ABC转运蛋白基因对非生物胁迫的响应,结果表明,内含子较少的MsABCs基因含有较多的非生物响应元件。基于系统发育树分析将33个蛋白聚为7个亚家族,通过分析红曲霉和其他菌属ABC转运蛋白家族的进化关系,发现与黑曲霉、烟曲霉等ABC转运蛋白的聚类结果基本一致,不同菌属的ABC转运蛋白的相同亚类聚集在一个分支。说明红曲霉与其他菌属的ABC转运蛋白具有较高的同源性,这为红曲霉ABC转运蛋白功能分析提供了参考依据。
先前的研究在烟曲霉[25]、黑曲霉[26]、白色假丝酵母[27]、酿酒酵母菌[28](Saccharomycescerevisiae)和新型隐球酵母[29]中分别鉴定出49,34,27,30和54个ABC转运蛋白基因。新型隐球酵母ABC基因数量较多与其染色体数目较多、基因组较大有关。在红曲霉中发现了33条MsABCs基因,与同曲霉属的黑曲霉中的ABC基因数量相近。在早期的研究中,ABC转运蛋白被分为8个亚家族,按照科学命名从ABCA-ABCH,行使的功能各不相同。ABCA在细胞体内主要介导脂质的运输[30];ABCB主要介导生长代谢产物以及外源物的运输有关[31];ABCC与细胞膜离子通道的调节过程相关[32];ABCD与脂肪酸的过氧化转入相关[33];ABCE和ABCF两种转运蛋白在大多数的生物内都具有较高的同源性,说明它们可能具有执行基础的运输功能;ABCG在真菌的多种抗药性基因起耐受作用[11];ABCH与有关抗药性的原核转运蛋白具有较高的同源性[34]。我们的研究中,系统发育树显示MsABCs基因被分成7种不同的亚家族。红曲霉ABC基因族中不存在ABCH亚家族,这可能是进化过程中基因丢失的结果。
在这些亚家族中,ABCC和ABCG是最大的亚家族,8个MsABCs基因属于这两类亚家族,在黑曲霉和烟曲霉中这两个亚家族也都有较高的表达。另根据进化树可以看出,不同物种的同一亚科的ABC序列比同一物种的ABC序列更相似,但属于不同的亚科。结果表明黑曲霉、烟曲霉、酿酒酵母和红曲霉中的ABC蛋白可能存在同源性,MsABCs成员与来自不同物种的同一亚科的成员的亲缘关系比与来自同一物种的其他ABC成员的亲缘关系更密切,这意味着同一ABC亚家族在不同物种之间具有较高的同源性,在不同物种之间ABC亚家族具有较高的保守性。
4 结论
本研究对红曲霉ABC家族进行了全基因组分析,确定了33条MsABCs序列。随后基于生物信息学方法,对MsABC序列在基因结构、系统发育、序列片段定位、基因复制、逆境相关顺式元件、基因组内共线性关系以及非生物逆境等方面进行了分析。多数MsABCs基因对环境胁迫作用敏感,说明MsABCs基因在红曲霉应对环境压力时起重要作用。本研究提供了有关红曲霉MsABCs基因家族的全面信息,并将有助于确定MsABCs基因的功能。
