烟酰胺单核苷酸腺苷转移酶1对视网膜发育过程中中央碳代谢稳定性的作用
2023-06-05金潇秦应祥罗凯
金潇,秦应祥,罗凯
(中国科学院大学重庆仁济医院1.眼科,2.药剂科,重庆 400000)
烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD)在氧化还原代谢中发挥核心作用,烟酰胺单核苷酸腺苷转移酶1(nicotinamide nucleotide adenylyltransferase 1,NMNAT1)还可作为许多酶的底物,包括SIRTs、PARPs、CD38、CD157 和 SARM1等,与衰老、细胞增殖、免疫、神经变性、分化和发育等多种生物学过程有关[1-5]。在哺乳动物组织中,视网膜似乎特别依赖于适量的NMNAT1来维持细胞生存和发挥生理功能[6-8]。NMNAT1是一种高度保守蛋白,可催化烟酰胺单核苷酸(NMN)或烟酸单核苷酸(NaMN)的腺苷酰化作用以形成NMNAT1,这是所有哺乳动物NMNAT1生物合成途径的关键步骤[9-10]。目前NMNAT1的多种超出其在氧化还原代谢中典型作用的功能受到了广泛的关注,本实验旨在探讨NMNAT1对小鼠视网膜中心碳代谢过程的影响。
1 材料与方法
1.1 动物模型
NMNAT1fl/fl小鼠(维通利华)与野生型 129/SV-E小鼠(维通利华)充分回交。将NMNAT1fl/fl小鼠与Six3启动子下表达Cre重组酶的转基因小鼠(Six3-Cre)杂交,获得条件敲除小鼠[11]。杂交产生杂合的Nmnatfl/wt和Six3-Cre Nmnatfl/wt后代,它们与NMNAT1fl/fl小鼠进一步杂交以产生接近孟德尔比率的条件敲除(Six3-Cre Nmnatfl/fl)和同窝对照(NMNAT1fl/fl)小鼠。实验小鼠定期与野生型129/SV-E小鼠回交以保持遗传完整性。将实验小鼠维持在标准的12 h光照/黑暗循环下,提供充足的食物和水。
使用来自耳部穿刺活检的基因组DNA,通过PCR对小鼠进行基因分型。Six3-Cre引物:5′-GCCCTGGCAGTGTCTTAG-3′;5′-CAGTGCGTGTCGTGGAGT-3′。NMNAT 1引物:5′-CCTTGTACCTGACCATGTCAACA-3′;5′-CCTGGGAGGCATAGACCATGTA-3′。引物以0.75 μmol·L-1的终浓度添加到PCR反应中。热循环条件为95 ℃ 2 min,随后完成35个热循环(95 ℃ 30 s,58 ℃ 30 s,72 ℃ 45 s),最后延伸步骤72 ℃ 5 min。
1.2 液相色谱串联质谱(liquid chromatography tandem mass spectrometry, LC-MS/MS)靶向代谢组学
在出生后第4天,对NMNAT1敲除组和空白对照组小鼠视网膜组织中的涉及基本生化途径的112种细胞代谢物的含量水平进行量化。将小鼠用断颈法处死后摘除眼球,将视网膜解剖后置于汉克平衡盐溶液(HBSS)中,并在液氮上快速冷冻。根据既往报道的方案[12]提取代谢物。代谢物提取物通过Shimadzu LC Nexera X2 UHPLC与QTRAP 5500 LC-MS/MS(AB Sciex)联用进行分析。ACQUITY UPLC BEH Amide分析柱(2.1×50 mm,1.7 μCorp.,Milford,MA)用于色谱分离。流动相是含有10 mmol·L-1乙酸铵(pH 8.9) 的水(A)和含有10 mmol·L-1乙酸铵(pH 8.2)的乙腈/水(体积比为95/5)(B)(所有溶剂均为Fisher Scientific的LC-MS Optima级)。总运行时间为11 min。流速为 0.5 ml·min-1。进样体积为5 μl,梯度洗脱为6 min时体积分数95%~61%(B)、8 min时体积分数61%~44%(B)、8.2 min时体积分数61%~27%(B)和9 min时体积分数27%~95%(B)。在每次运行结束时用体积分数95%(B)平衡色谱柱。碰撞气体为N2。正负模式下的离子源条件为:气帘(CUR)=25 psi,碰撞气(CAD)=高,离子喷雾电压(IS)=3 800/-3 800 V,温度(TEM)=500 ℃,离子源气体1(GS1)=50 psi,离子源气体2(GS2)=40 psi。每种代谢物都使用标准品进行了调整,以获得最佳转换。 D4-烟酰胺(Cambridge Isotope Laboratories, Tewksbury, MA)用作内标。使用 MultiQuant 3.0.3 软件(AB Sciex, Concord, ON, CA)对提取的 MRM 峰进行积分。
1.3 视网膜RNA提取、测序和分析
将小鼠用断颈法处死后摘除眼球,将视网膜解剖后置于含有少量 Trizol®试剂(美国Thermo Fisher Scientific公司)的无菌微量离心管中,并在干冰上快速冷冻。用手持匀浆器匀浆解冻样品并孵育5 min,提取总RNA。向每个样品中加入 20 μl氯仿,将样品短暂涡旋,在室温下孵育4 min,然后以12 000 r·min-1离心10 min。将上清液移置含有50 μl异丙醇的离心管中,并在室温下孵育10 min。最后,样品再次以12 000 r·min-1离心10 min。去除上清液,用体积分数75%乙醇洗涤沉淀3次,干燥后重悬于DEPC处理过的水中。 全转录组测序在NovaSeq 6000平台上使用 Illumina TruSeq Stranded mRNA Library Prep Kit进行,每个样本的总读数深度为100 M。使用FastQC验证读取质量,并使用BBTools 包的Bbduk实用程序修剪引物。使用HISAT2 2.1.0进行读取比对,使用 StringTie 1.3.6组装和量化转录本。使用DESeq2 R包鉴定差异表达的基因。
1.4 统计学处理
数据采用SPSS 25.0统计软件进行分析。每组实验结果均重复3次。采用独立样本t检验对数据进行假设检验,检验标准:α=0.05。
2 结 果
2.1 NMNAT1敲除小鼠视网膜的整体代谢改变
LC-MS/MS分析显示,NMNAT1敲除小鼠视网膜中共有39种代谢物的含量发生了显著改变(P<0.05)。代谢物集富集分析发现多种不同生化途径的均受到了不同程度的破坏,主要包括氨基酸代谢、糖酵解/糖异生、烟酸和烟酰胺代谢以及嘌呤代谢(图1)。
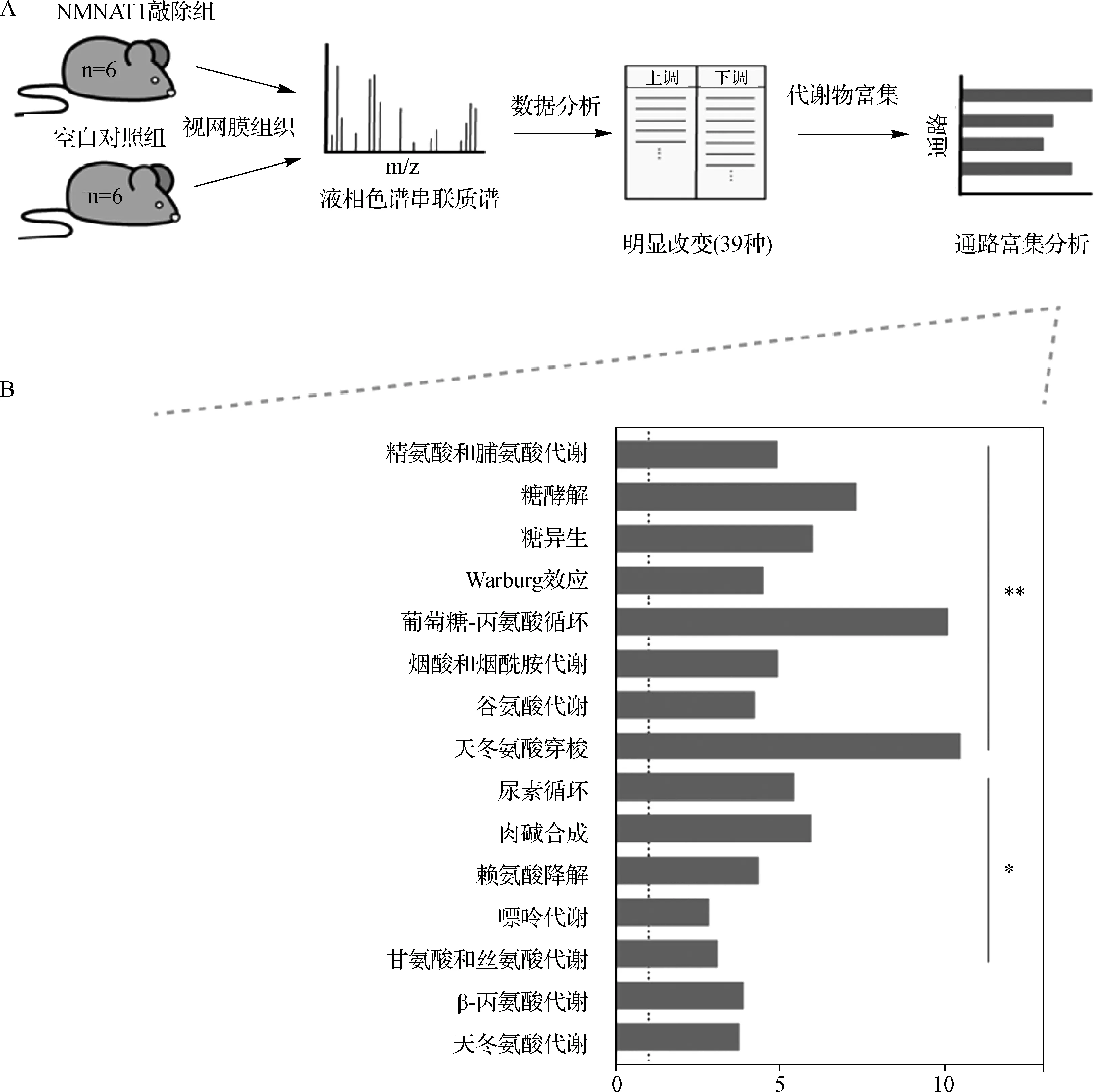
A.代谢组学实验方法的示意图;B.NMNAT1敲除小鼠视网膜中显著变化的代谢物的集富集分析图1 代谢组学富集分析A.Schematic diagram of the experimental approach to metabolomics;B.Set enrichment analysis of significantly altered metabolites in the retina of NMNAT1 knockout miceFig 1 Metabolomics enrichment analysis
2.2 视网膜NMNAT1缺乏对NMNAT1通路的影响
NMNAT1敲除组小鼠视网膜总NMNAT1水平和烟酸腺嘌呤二核苷酸(NaAD) 水平降低约 40%(图2A)。NMNAT1和NaAD均是 NMNAT1 的催化产物(图2B)。下游代谢物NADP和烟酰胺(NAM)的水平分别降低了约25%和55%,而NADH 的水平略有降低(P>0.05)(图2A)。 NAD前体烟酰胺核苷(NR)和烟酰胺单核苷酸(NMN)在NMNAT1敲除组小鼠视网膜中的显著积累(图2A);色氨酸水平没有显著变化,色氨酸是这个时期从头合成NAD的起点(图2A)。
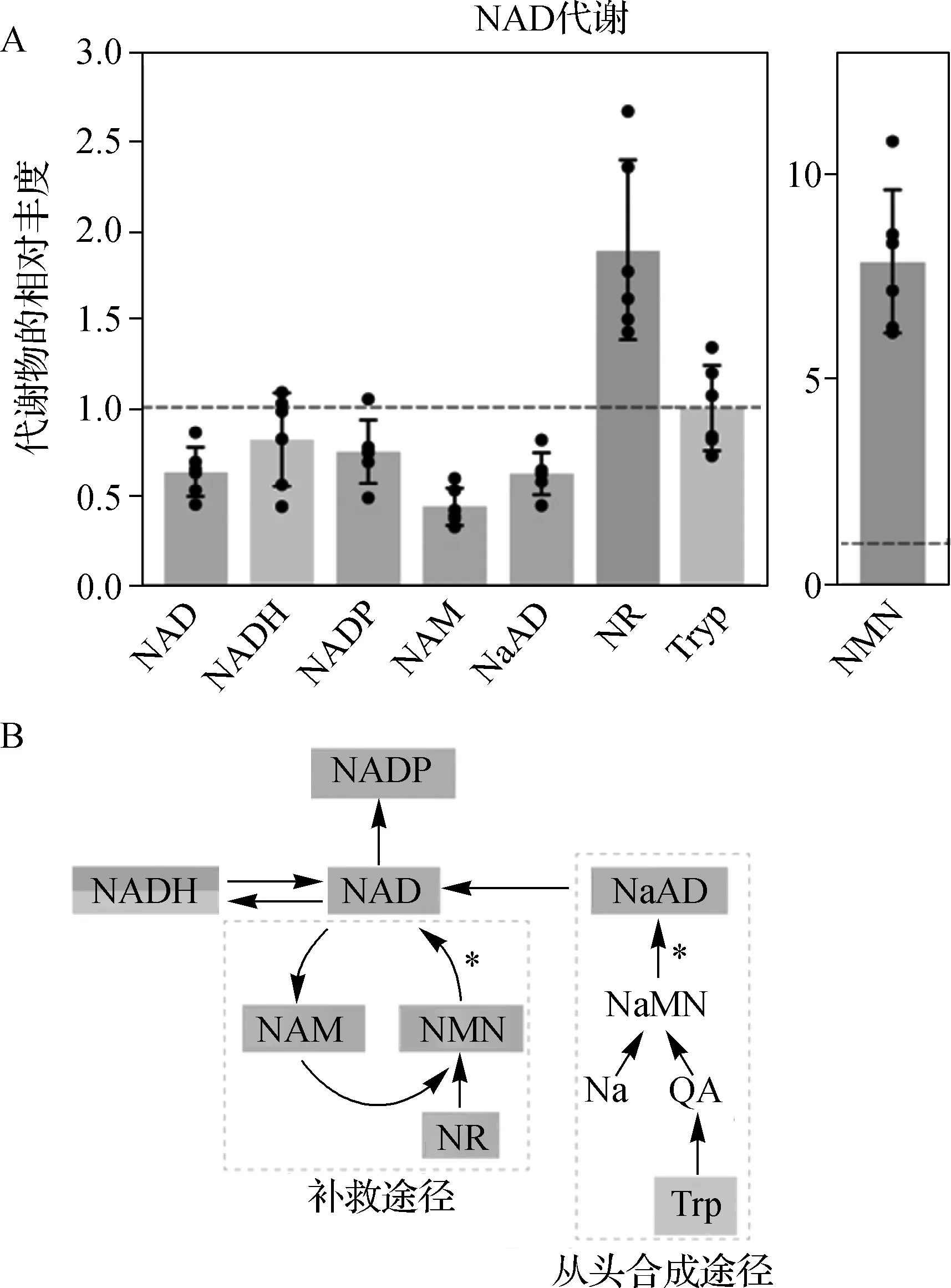
A.质谱法评估小鼠视网膜中 NMNAT1通路代谢物的相对丰度;B.哺乳动物NMNAT1合成途径(NMNAT1催化的步骤以星号表示)图2 NMNAT1敲除小鼠视网膜组织NMNAT1通路代谢物含量的变化A.The relative abundance of NMNAT1 pathway metabolites in the retinas of mice assessed by mass spectrometry;B.Mammalian NMNAT1 synthesis pathway(steps catalyzed by NMNAT1 were indicated with asterisk)Fig 2 Changes of the content of NMNAT1 pathway metabolites in the retinal tissues of NMNAT1 knockout mice
2.3 NMNAT1敲除组小鼠视网膜组织中的糖酵解、三羧酸循环和肌酐代谢的变化
NMNAT1敲除组小鼠视网膜中显著改变的代谢物中富含与糖酵解、糖异生和瓦博格效应相关的代谢物(图1B)。 对这些途径的进一步检查显示上游糖酵解代谢物葡萄糖、葡萄糖6-磷酸(G6P) 和果糖1,6-二磷酸(F16BP) 的水平大幅升高,同时磷酸二羟丙酮(DHAP)和葡萄糖 3-磷酸(G3P) 的水平大幅降低(图3)。这表明葡萄糖利用的中断,与这种效应一致的是,在NMNAT1敲除组小鼠视网膜中,三羧酸循环中间体 α-酮戊二酸(a-KG) 和琥珀酸的水平降低了约30%(图3)。NMNAT1敲除组小鼠视网膜中ATP循环代谢物磷酸肌酸和肌酸的水平降低,但差异无统计学意义(P>0.05)(图3B)。
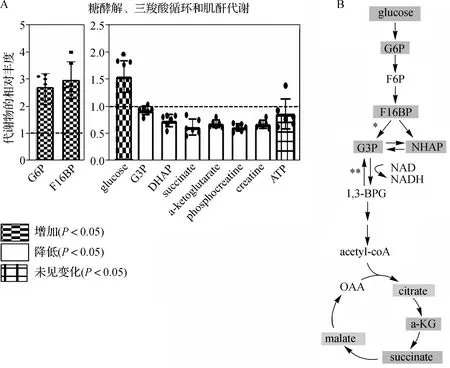
A.质谱法评估NMNAT1敲除组小鼠视网膜中糖酵解、三羧酸循环和肌酐代谢物的相对丰度;B.根据(A) 中代谢物变化着色的糖酵解/三羧酸循环途径(二磷酸果糖酶催化的步骤用单星号表示, GAPDH催化的步骤用双星号标记)图3 NMNAT1敲除组小鼠视网膜组织糖酵解、三羧酸循环和肌酐代谢的变化A.Mass spectrometry was used to assess the relative abundance of glycolysis, TCA cycle and creatinine metabolites in the retinas of NMNAT1 knockout group mice;B.Glycolysis/tricarboxylic acid cycle pathway coloured according to metabolite changes in(A)(steps catalysed by fructokinase diphosphate were indicated with a single asterisk, whereas those catalysed by GAPDH were marked with a double asterisk)Fig 3 Changes of glycolysis, tricarboxylic acid cycle and creatinine metabolism in retinal tissues of NMNAT1 knockout group mice
2.4 NMNAT1敲除组小鼠视网膜糖酵解和三羧酸循环相关酶表达水平的变化
高通量mRNA测序分析显示, NMNAT1敲除组小鼠视网膜中16种糖酵解和4种三羧酸循环酶的表达水平发生改变(图4)。

图4 NMNAT1敲除组小鼠(-/-)和对照组(+/+)糖酵解和三羧酸循环相关酶表达水平的热图Fig 4 Heat map of glycolysis and tricarboxylic acid cycle-related enzyme expression levels in NMNAT1 knockout mice(-/-) and controls(+/+)
2.5 NMNAT1敲除组小鼠视网膜组织磷酸戊糖途径代谢物相对丰度的特异性改变
LC-MS/MS结果表明,NMNAT1敲除组小鼠视网膜组织中的赤藓糖醇(Erythritol)水平显著降低(P<0.05),提示戊糖磷酸途径(PPP) 可能受到破坏(图5)。这与NMNAT1敲除组小鼠视网膜组织中糖酵解通路的破坏和NMNAT1敲除组视网膜中 NADP水平的降低一致。
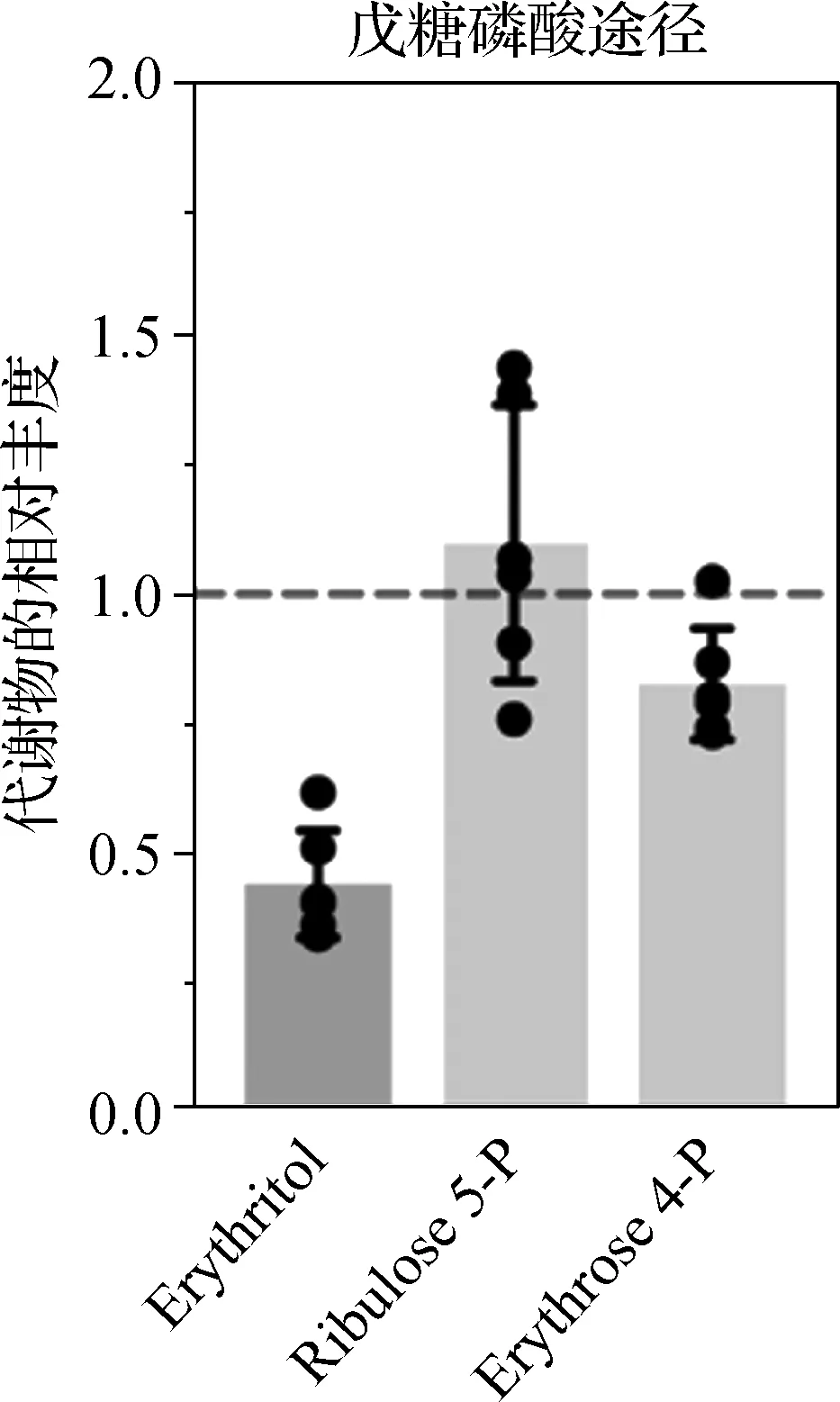
图5 NMNAT1敲除组小鼠视网膜组织磷酸戊糖途径代谢物相对丰度的特异性改变Fig 5 Specifically alteration of the relative abundance of pentose phosphate pathway metabolites in the retinal tissues of NMNAT1 knockout mice
3 讨 论
NMNAT1在不同组织细胞中广泛表达,并且其中许多突变会降低NMNAT1的催化活性或压力相关稳定性[13-15], 目前已有超过30种NMNAT1基因突变被证明与Leber 先天性黑蒙和视锥视杆细胞营养不良有关[9-10,13-14,16-17]。但NMNAT1的突变很少会导致眼外表型[18],此外另外两种 NMNAT 亚型(高尔基体相关 NMNAT2 和线粒体相关NMNAT3)均可在视网膜中检测到但与失明无关[19],因此NMNAT1在视网膜中的作用备受关注。
此前有研究通过对光感受器中缺乏NMNAT1途径酶NAMPT小鼠的研究,描述了视网膜中NMNAT1具有关键作用[6]。此外NMNAT1是迄今为止唯一与失明有关的NMNAT1途径酶,因此NMNAT1异常导致的视网膜发育异常的机制有待进一步研究。最近已有几项报告指出,视网膜NMNAT1部分或完全抑制时,小鼠在出生后第1周内开始出现视网膜变性,并在1个月大时基本完成视网膜变性[20],而小鼠中NMNAT1的整体缺失将导致小鼠在胚胎死亡[21]。目前的研究结果表明NMNAT1在视网膜中具有多种潜在的不同作用,体外研究表明 NMNAT1促进去乙酰化酶功能以促进视网膜祖细胞的存活[19],而成熟小鼠中NMNAT1的缺乏导致由NADase SARM1介导的光感受器快速死亡[22]。虽然这些研究表明视网膜 NMNAT1的多种功能超出其在氧化还原代谢中的典型作用,但这些功能重叠的程度以及动物模型和患者中NMNAT1相关视网膜营养不良严重程度的机制尚未被全面探索。
根据代谢组学结果可以推测出NMNAT1合成了视网膜总NMNAT1池中的约40%,这与之前的报道的模型[22]大致一致。糖酵解通量受损似乎是组织中NMNAT1耗竭的一个普遍特征,因为既往研究报告了投射神经元和骨骼肌肌管NAMPT抑制或缺失时,将导致GAPDH上游糖酵解代谢物的积累[23]。值得注意的是,Lundt等[23]提出了在NAMPT抑制的肌管中糖酵解通量逆转的证据,因此我们认为这一效应可能解释了我们模型中G3P和DHAP水平的下降。
除了糖酵解障碍,我们还检测到NMNAT1基因敲除小鼠视网膜中嘌呤核苷酸和氨基酸代谢途径的特异性缺陷。作为一种特别增生的组织,视网膜被认为高度依赖于足够的核苷酸和氨基酸来支持细胞的转录和翻译[24]。本实验中的一些代谢变化,例如嘌呤前体黄嘌呤和氨基酸天冬氨酸的积累,似乎与NMNAT1不足或视网膜变性有更广泛的联系。另一方面,我们还确定了这些途径中的一系列代谢变化,这些变化在NMNAT1基因敲除视网膜中尚未被报道,因此在未来的研究中有待进一步探索。
在这项研究中,我们通过生成和表征视网膜特异性NMNAT1敲除小鼠模型来研究 NMNAT1介导的NMNAT1代谢在视网膜中的作用。利用代谢组学和转录组学方法,我们证明了NMNAT1缺失会导致视网膜中严重的特异性代谢缺陷,并导致中央碳、嘌呤核苷酸和氨基酸代谢均受到损害,并且可能是导致严重视网膜变性的原因。总体而言,我们的结果揭示了 NMNAT1相关视网膜变性中以前未被重视的复杂性,为NMNAT1缺陷的视网膜特异性表现提供了可能的解释,并为进一步研究视网膜中NMNAT1代谢奠定了基础。
