高产IAA哈茨木霉突变株M95的筛选及其对黄瓜的促生作用
2023-05-29高雅新王甲果张翔凯李驰马磊刘东阳沈其荣
高雅新,王甲果,张翔凯,李驰,马磊,刘东阳,沈其荣
(南京农业大学江苏省固体有机废弃物资源化高技术研究重点实验室/江苏省有机固体废弃物资源化协同创新中心,江苏 南京 210095)
实现乡村生态振兴是我国当前农业发展的重要目标,而化肥的过量施用导致土壤质量下降、土地富营养化、重金属污染、生态平衡失调,严重破坏乡村生态环境[1]。生物有机肥中的微生物可以通过分解难溶的植酸钙等营养物质和分泌植物激素促进植物生长;同时可以通过诱导植物抗病基因的表达和分泌抗生素等抑制病原菌的生长。生物有机肥的合理施用有助于维持良好生态环境和土壤生产力,是解决乡村生态问题的有效途径[2]。目前,生物有机肥中存在功能微生物种类单一,产品效应不稳定等问题。因此,筛选高效、稳定的微生物功能菌对生物有机肥行业发展具有重要意义。木霉菌(Trichodermasp.)属于半知菌类真菌[3],是土壤及根际中广泛存在的一种有益微生物。它不仅对多种病原菌具有显著拮抗作用,且对黄瓜、大豆和茶树等植物具有明显的促生作用[4-6]。国内外已有超过50种木霉商品化制剂成功应用于农业生产过程中[7]。哈茨木霉(Trichodermaharzianum)NJAU 4742是本实验室筛选获得的植物有益真菌(PGPF),其分泌的Swollenin蛋白(TgSWO)可作用于黄瓜植株根冠和侧根发生部位,通过YoaJ结构域介导细胞壁的扩张并改变黄瓜根系结构,促进侧根的发生及木霉的定殖,进而促进黄瓜的生长[8]。同时,NJAU 4742能够通过分泌类植物激素Harzianolide促进植株根系伸长和根尖分化,进而促进植株苗期的生长;并且Harzianolide还能增加植株体内抵御病原微生物相关酶的活性,从而提高番茄对核盘菌的系统抗性[9]。Li等[10]研究发现,NJAU 4742能够通过活化土壤中的磷以及微量元素等矿质元素间接促进番茄植株的生长。另外,NJAU 4742可以合成和分泌生长素(IAA)进入宿主根系,这也是其促进植株生长的重要机制之一[11]。木霉合成IAA主要通过色氨酸依赖途径,其IAA生物合成中的关键基因包括吲哚乙酰胺水解酶基因(iaaH2)、腈水解酶基因(nit1和nit2)和吲哚乙醛脱氢酶基因(ald1和ald2)[11-12]。
目前,对于NJAU 4742的促生机制研究已有较多报道,但通过突变提高其促生能力及其机制的研究还较少。本研究以NJAU 4742作为原始菌株,通过辐射突变获得1株高产IAA突变株M95,其生长素合成关键基因的转录水平显著提高,通过水培试验和盆栽试验评估M95对黄瓜苗期的促生作用,为制备新型木霉有机肥提供新的材料和理论基础,并为后续进一步探究NJAU 4742的促生机制提供良好的试验材料。
1 材料与方法
1.1 供试菌株与材料
哈茨木霉NJAU 4742由本实验室保存,该菌株ITS基因组序列在NCBI的登录号为GQ 337429。为防止菌种的退化,每次使用的菌种都从-80 ℃甘油保存的孢子悬液中活化。菌株M95由NJAU 4742通过辐照突变而来,并保存于-80 ℃甘油中。
黄瓜(CucumissativusL.)品种为‘露丰’,购于江苏省农业科学院;发酵用玉米秸秆和米糠购于当地农贸市场,育苗基质和植物有机肥(pH7.69,全氮含量17.08 g·kg-1,全磷含量7.13 g·kg-1,全钾含量14.41 g·kg-1,有机质含量453.00 g·kg-1)由江苏禾喜生物科技有限公司提供,供试土壤采自江苏省宜兴市水稻土(31°20′41″N,119°45′60″E),水稻土pH7.00,有机质含量5.27 g·kg-1,总氮含量0.25 g·kg-1,全磷含量0.35 g·kg-1,有效磷含量6.13 mg·kg-1,速效钾含量68.00 mg·kg-1。
Salkowski指示剂:将10 mL 0.5 mol·L-1FeCl3溶液与500 mL 35%(体积分数)高氯酸混合,避光保存。PDA培养基:39.00 g PDA(购自BD Difco)加去离子水定容至1 L。马铃薯葡萄糖液体培养基(PDB):200.00 g马铃薯(去皮),20.00 g葡萄糖,定容至1 L。MS培养基:4.74 g MS培养基,加去离子水定容至1 L。1/4 MS营养液:1.19 g MS培养基(海博生物,HB8469-5),加去离子水定容至1 L。上述培养基配制后需115 ℃灭菌30 min备用。
1.2 木霉突变株的制备与筛选
1.2.1 木霉的活化以及孢子悬液的制备将-80 ℃甘油保存的NJAU 4742孢子悬液取出,滴加在PDA平板上,置于28 ℃培养箱中培养至菌丝长满,再置于28 ℃光照培养箱中培养2~3 d,待长满绿色孢子后,每个平板加入3 mL无菌去离子水,用涂布棒将菌丝轻轻刮下,然后通过4层纱布过滤,所得滤液即为木霉孢子悬液,在显微镜下利用血球计数板测定孢子悬液中的孢子含量。
1.2.2 木霉突变株的制备将准备好的NJAU 4742孢子悬液用无菌去离子水稀释至1×107mL-1,与醋酸-醋酸钠缓冲液1∶1(体积比)混合后加入96孔板中,并贴上封板膜,置于生物X射线辐照器(RS 2000 Pro,RADSOURCE,美国)中进行辐射,辐射条件如下:分别按照84、84和82 Gy的剂量进行3次照射,剂量数共计250 Gy。辐射后获得木霉突变株,保存在-80 ℃甘油中待用。
1.2.3 木霉突变株的初筛将木霉突变株菌悬液涂布到PDA平板上,待孢子萌发后,挑选长势良好的突变株,将其培养至产孢并制成孢子悬液,然后将孢子悬液滴加至PDA平板上,培养2~3 d后,取直径为 6 mm 的菌饼移接至新制备的PDA平板上,28 ℃培养36 h后与NJAU 4742比较其生长半径,挑选半径较大且形态正常的木霉突变株进行下一步试验。
1.2.4 木霉突变株的复筛木霉发酵液中IAA的提取:将不同木霉突变株的孢子悬液(1×107mL-1)接种于含250 μmol·L-1色氨酸和10 g·L-1葡萄糖的MS培养基中,28 ℃、170 r·min-1避光培养7 d。将培养7 d的发酵液4 ℃、8 000 r·min-1离心10 min,取上清液,经0.22 μm无菌滤膜过滤后获得无菌发酵液。将上述发酵液用2 mol·L-1HCl调节pH值至2.00,再用等体积乙酸乙酯萃取3次,萃取液在37 ℃下进行旋转蒸发,将浓缩物质溶解于1 mL甲醇中待测。
将木霉发酵液提取物与Salkowski指示剂[13]按照1∶2(体积比)比例混合后进行显色反应,在避光条件下静置30 min,观察颜色变化(变为粉红色表示有IAA产生),将颜色显著深于野生型的突变株进行更深入的研究。利用IAA标准品(Sigma)配成浓度为1、5、10、15、20和25 mg·L-1的溶液备用,将上述IAA标准溶液和木霉发酵液提取物分别与Salkowski指示剂按1∶2(体积比)的比例混合,避光反应30 min,用酶标仪测定D535值,每个处理进行3次生物学重复。以含色氨酸和葡萄糖的MS培养基提取物调零并作为空白对照,根据标准曲线计算不同处理中IAA含量,选取产量较高的5个处理进行下一步试验。
HPLC法测定IAA产量:利用高效液相色谱(HPLC,Waters 96)进一步测定IAA产量。将IAA标准品溶于甲醇后,配成浓度为50、100、200、300、400和500 μg·L-1的溶液备用。将不同浓度的IAA标准品和木霉发酵液提取物进行HPLC分析,采用Waters C18柱,流动相为A(甲醇)、B(0.1%乙酸)的体积比为 60∶40,流速为0.4 mL·min-1,柱温为25 ℃,进样量为10 μL,检测波长为220 nm。
1.3 WT和M95的碳源利用特征分析
用Biolog FF微平板(Biolog,美国)测定野生株NJAU 4742(WT)和M95的碳源利用情况,比较其碳源利用特性。用棉签蘸取少量木霉孢子置于FF接种液中,混合均匀后用浊度仪(Turb 550,WTW,德国)调节浊度至75%,保持每个样品的浊度一致,向FF板的每个微孔中加入100 μL悬浮液,置于28 ℃培养箱中培养,前4 d每隔12 h测定1次D值,后3 d每隔24 h测定1次,测定波长为490和750 nm。Biolog FF微平板中具有95种碳源,可以反映菌株在生长过程中对不同碳源的利用情况[14]。
1.4 WT和M95中IAA合成关键基因的定量分析
将WT和M95的孢子悬液(1×107mL-1)接种于含250 μmol·L-1色氨酸和10 g·L-1葡萄糖的MS培养基中,28 ℃、170 r·min-1避光培养7 d,用无菌纱布过滤,无菌水清洗3次,收集菌体并沥干水分,将菌体用液氮冷冻后用RNA提取试剂盒(RNeasy Plant Mini Kit,凯杰生物,德国)提取总RNA,将RNA浓度稀释至基本一致后,用反转录第1链cDNA合成试剂盒(RevertAidTMFirst Strand cDNA Synthesis Kit,赛默飞,美国)合成第1链cDNA并进行实时荧光定量分析(QTOWER实时荧光定量PCR仪,德国)。使用tef基因作为内参基因,基因相对转录水平用2-ΔΔCT计算[15]。特异性引物(表1)均用Premier 6软件设计。
1.5 黄瓜水培试验
1.5.1 萌发孢子液的制备取1 mL 1×108mL-1木霉孢子悬液于100 mL PDB培养基中,28 ℃、170 r·min-1培养24 h,经无菌滤纸过滤后,用无菌水冲洗3次,并稀释至1×107mL-1获得萌发孢子液,置于4 ℃冰箱待用。

表1 哈茨木霉中生长素相关基因的定量引物Table 1 Primers used for quantification the transcriptional changes in auxinbiosynthesis-related genes of Trichoderma harzianum
1.5.2 黄瓜种子的消毒及催芽将黄瓜种子在50 ℃无菌去离子水中浸泡30 min,随后立即转入30 ℃无菌水浸泡1 h;将水倒掉后,先用70%的乙醇浸泡3~4 min,浸泡过程要持续晃动,再用无菌水冲洗3次,然后用2%的次氯酸钠浸泡3~4 min,最后用无菌水多次冲洗,直至将次氯酸钠冲洗干净。挑选籽粒饱满的种子置于30 ℃培养箱中进行催芽,48 h后转入26 ℃的温室培养[16]。
1.5.3 水培试验待黄瓜幼苗的2片子叶展开后,移入装有50 mL 1/4 MS营养液的50 mL锥形瓶中,继续在26 ℃的温室中培养至2叶1心期,挑选长势一致的幼苗移入装有不同处理营养液的100 mL锥形瓶中振荡培养,观察黄瓜幼苗长势。共设置3个处理:1)CK,仅用100 mL 1/4MS营养液培养;2)WT,99 mL 1/4 MS营养液中加入1 mL NJAU 4742萌发孢子液;3)M95,99 mL 1/4 MS营养液中加入1 mL M95萌发孢子液。每个处理10个重复。温室条件下每隔1~2 d添加1次营养液,保持营养液为100 mL。
1.6 黄瓜盆栽试验
1.6.1 木霉固体发酵取1 mL 1×107mL-1的木霉孢子悬液,加入100 mL PDB培养基中,28 ℃、180 r·min-1培养至液体变为绿色。将玉米秸秆自然风干后,用铡刀将其铡成1~2 cm的片段,与过1 mm筛的米糠和水按1.0∶0.3∶2.0(质量/质量/体积)的比例进行混合,同时加入6.67%的氨基酸水解液,调节pH值至2.50,获得的固体基质在115 ℃灭菌60 min。冷却后,按照100 g·L-1的比例接种木霉发酵液,装入发酵盘中进行浅层发酵,28 ℃条件下避光培养3~4 d,获得木霉高密度固体菌种(孢子浓度为1×108g-1)。
1.6.2 黄瓜种子的催芽及育苗将消过毒的黄瓜种子均匀摆放在铺有无菌滤纸的90 mm培养皿中,滤纸用2 mL无菌去离子水润湿,置于30 ℃暗培养2 d,待胚芽长至约1 cm时,转移至含有育苗基质的育苗穴盘中,每穴1粒种子,每日浇水1次。
1.6.3 盆栽试验待黄瓜幼苗生长至2叶1心时,挑选长势一致的幼苗移入装有3 kg不同处理土壤的盆钵中,并观察其长势,待具有显著性生长差异时拍照记录,并测量各项生物指标。共设置3个处理:1)CK,添加有机肥40.00 g;2)WT,添加含5%(质量分数)NJAU 4742高密度固体菌种的有机肥40.00 g;3)M95,添加含5%(质量分数)M95高密度固体菌种的有机肥40.00 g。每个处理10个重复。常规田间管理,每日浇水2次。
1.7 测定方法
1.7.1 土壤及有机肥养分测定利用pH计测定土壤和有机肥pH值(水土质量比和水肥质量比均为2.5∶1);利用元素分析仪(Vario MAX,Elementar,德国)测定土壤和有机肥总碳、总氮含量;分别采用氢氧化钠-钼锑抗分光光度法、碳酸氢钠-钼锑抗比色法和乙酸铵-火焰光度法测定土壤全磷、速效磷和速效钾含量[17]。有机肥全磷和全钾的测定参照《有机肥料:NY/T 525—2021》。采用硫酸-过氧化氢消解,分光光度法测定有机肥全磷含量;采用硫酸-过氧化氢消解,火焰光度法测定有机肥全钾含量。
1.7.2 黄瓜生物性状待每个处理的生物性状出现显著性差异时,分别测定不同处理的株高、茎粗、根长、叶绿素、叶面积以及地上部和地下部干/鲜重。
1.7.3 根际土壤DNA提取将黄瓜根系轻轻抖动去除附着在根系表面的土壤后,用无菌剪刀将采集的各样品根系剪成1 cm长的小段,放入含有50 mL无菌水和5颗玻璃珠(直径5 mm)的锥形瓶中,28 ℃、170 r·min-1振荡30 min,然后用超声波清洗机超声处理20 min后将根系去除,获得土壤悬浮液;将土壤悬浮液在4 000 r·min-1、4 ℃条件下离心10 min,弃去上清液,沉淀即为根际土壤,放置在-80 ℃超低温冰箱中保存待用。将根际土壤样品取出后称取0.50 g,采用土壤DNA提取试剂盒(QIAGEN GmbH,德国)严格按照其操作说明进行根际土壤总DNA的提取。每个处理设置3个重复,其中每个重复由提取的3份DNA混合而成。采用核酸定量仪NanoDrop 2000(Thermo Scientific,Waltham,MA,美国)将提取的DNA进行定性定量测定,并统一稀释至25.00 ng·μL-1,保存于-20 ℃冰箱待用。
1.7.4 根际土壤总细菌、总真菌和总木霉数量的测定利用实时PCR荧光定量仪(Applied Biosystems,美国)测定根际土壤中总细菌、总真菌和总木霉的数量。每个处理设置3个生物学重复,用无菌去离子水作为阴性对照。所有引物具有相同的扩增条件,PCR体系:引物F(10 μmol·L-1)1 μL,引物R(10 μmol·L-1)1 μL,2×IQTMSYBR Green Supermix 10 μL,模板DNA(25.00 ng·μL-1)1 μL,加无菌去离子水至20 μL。qPCR反应条件:95 ℃ 6 min,95 ℃ 15 s,60 ℃ 1 min,进行35个循环,产物熔解曲线条件为:55~95 ℃,每6 s升高温度0.5 ℃。特异性引物(表2)均用Premier 6软件设计。基于Morrison等[18]的方法,根据样品阈值(CT)计算出每克干土中总细菌、总真菌和总木霉的拷贝数,结果以对数表示[lg(copies·g-1)]。

表2 总细菌、总真菌和总木霉的定量引物Table 2 Primers for quantifying the total bacteria,fungi and Trichoderma sp.
1.8 数据分析
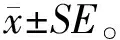
2 结果与分析
2.1 木霉突变株的初筛
从突变体库中挑选出99株突变株接种到PDA平板上,并观察其生长形态及菌落半径(图1)。其中,有10株突变株无论是生长速度还是菌落形态与野生株NJAU 4742(WT)相比都发生明显的变化,推测是菌株核心代谢通路的关键基因发生了突变,这样的突变株不适合于后期的深入研究,因此直接舍弃。突变株的初筛结果表明,在未舍弃的突变株中,有9株突变株与WT生长完全一致,培养48 h后,菌落半径为2.70 cm。有38株突变株菌落半径显著大于野生株,生长速度最快的为M36,菌落半径达3.25 cm,增加了20.37%,其次是M10、M12、M14和M95,其菌落半径均为3.20 cm,约为WT的1.19倍。挑选菌落形态正常且半径显著大于WT的38株突变株,进行下一步试验。
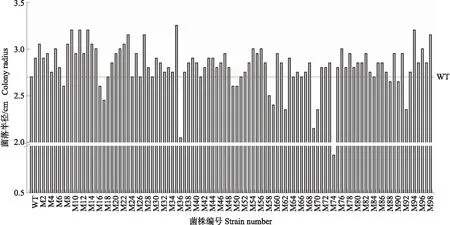
图1 不同NJAU 4742突变株的菌落半径Fig.1 The colony radius of different mutant strains of NJAU 4742WT:野生株NJAU 4742 Wild strain NJAU 4742. 下同 The same as follows.
2.2 木霉突变株的复筛
利用Salkowski比色法进一步比较通过初筛获得的突变株产IAA的能力。结果表明,与WT相比,大多数突变株都能产生一定量的IAA。通过粉色深浅的定性试验,初步舍弃4株几乎未产生IAA的突变株,挑选出26株颜色相对较深的突变株并对其产生的IAA进行定量分析。定量结果表明,WT发酵液中IAA质量浓度为0.39 μg·mL-1,而有17株突变株IAA的产量高于WT,其中M95的发酵液IAA质量浓度最高,达0.46 μg·mL-1,比WT提高了20.51%,其次是M8、M14、M71和M92,其发酵液的IAA质量浓度分别为0.46、0.46、0.44和0.43 μg·mL-1(图2)。挑选出M8、M14、M71、M92和M95这5株突变株,用HPLC法进一步测量其发酵液的IAA质量浓度,各处理的产量分别为456.81、454.92、436.33、427.40和463.42 μg·L-1,该结果与Salkowski比色法测定结果一致。因此,确定突变株M95为IAA高产的突变株,保存后用于后续试验。
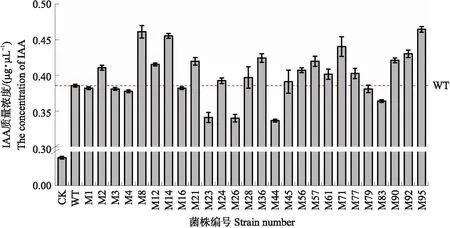
图2 Salkowski比色法测定NJAU 4742及其突变株发酵液中生长素(IAA)含量Fig.2 Detection of IAA production in fermentation broth of NJAU 4742 and its mutants by Salkowski reagent
2.3 突变株碳源利用特征
采用Biolog FF微孔板方法比较M95与NJAU 4742(WT)碳源利用情况,检测结果如图3所示。结果表明,M95与WT对大多数碳源的利用主要在前24 h,其生长速率大致为先增加后降低的趋势。影响两者生长的碳源主要是一些糖类及其衍生物质,而这些物质对两者影响的差异主要出现在生长中后期,尤其是72 h以后。当以D-阿拉伯糖醇、龙胆二糖和α-D-葡萄糖作为唯一碳源时,M95与WT的生物量从48 h开始差异显著,且以D-阿拉伯糖醇为唯一碳源时在培养120 h后二者生物量差异最为显著,M95的生物量比WT增加了72.65%。在以龙胆二糖和α-D-葡萄糖为唯一碳源的处理中培养96 h,M95的生物量比WT分别增加了53.59%和102.00%。当以半乳糖苷果糖作为唯一碳源培养120 h,M95的生物量比WT增加了84.66%。当以D-山梨糖醇作为唯一碳源时,M95的生物量在144 h时与WT差异最为显著,M95的生物量为WT的1.51倍。在i-赤藓醇、D-果糖、D-半乳糖、丙三醇、D-甘露糖和D-海藻糖作为唯一碳源的处理中,培养168 h的处理间差异最为显著,M95的生物量分别为WT的3.25、3.93、3.15和2.36倍。而在L-岩藻糖为唯一碳源处理中,M95与WT的生物量从5 d起开始显现出差异,7 d 时达到最大,M95的生物量比WT增加了99.18%。上述结果表明,i-赤藓醇、D-果糖、D-山梨醇和D-海藻糖更能促进M95的生长,且i-赤藓醇和D-果糖对M95和WT的生长影响差异显著,因此推测这4种碳源可能有利于M95的代谢,是否能够促进其产IAA还有待进一步深入研究。
图3 NJAU 4742和M95对不同碳源的利用率及其在不同碳源中的生长情况Fig.3 Utilization intensities of different carbon sources by NJAU 4742 and M95 and the growth rate ofNJAU 4742 and M95 at different carbon sources
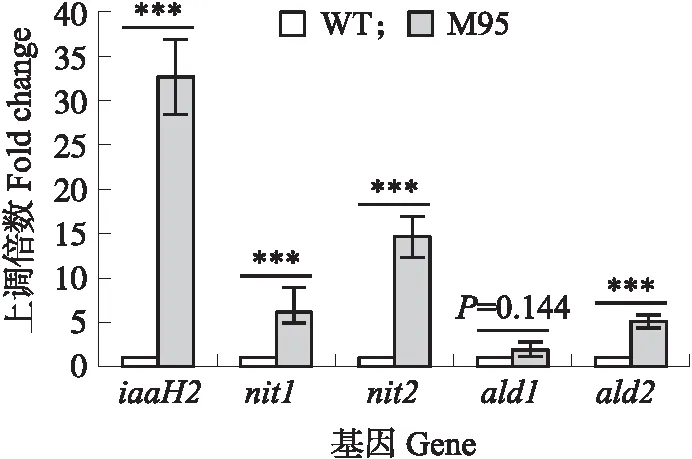
图4 M95 IAA合成相关基因的表达Fig.4 Expression of IAA biosynthesis gene in M95以WT为基准,WT的各基因表达水平上调倍数视为1。Based on WT,fold change of each gene expression level in WT is considered as 1. ***P<0.001.
2.4 M95和NJAU 4742中IAA合成关键基因的定量分析
根据Liu等[11]的研究结果,NJAU 4742(WT)主要通过色氨酸依赖途径合成IAA,而其关键基因为吲哚乙酰胺水解酶基因(iaaH2)、腈水解酶基因(nit1和nit2)和吲哚乙醛脱氢酶基因(ald1和ald2)。为了探究M95产IAA能力提高的分子机制,用荧光定量PCR方法对比了M95与WT中吲哚乙酰胺水解酶基因、腈水解酶基因和吲哚乙醛脱氢酶基因的转录水平。其结果如图4所示,与WT相比,M95与吲哚乙酰胺途径相关的iaaH2、与吲哚乙腈途径相关的nit1和nit2以及与吲哚丙酮酸途径相关的ald1和ald2的转录水平均有不同程度的上调。其中iaaH2的表达水平上调最大,为野生型的32.68倍,nit1和nit2分别上调了6.09和14.58倍,而ald1和ald2仅上调1.90和5.08倍。以上结果表明,M95合成生长素的能力显著高于WT,且主要变化集中在吲哚乙酰胺途径和吲哚乙腈途径,尤其是iaaH2和nit2基因表达水平的上调倍数最大,因此推测M95产IAA能力提升主要由于吲哚乙酰胺水解酶和腈水解酶的增加。
2.5 黄瓜水培试验结果
分别将WT和M95的萌发孢子接种到含有100 mL 1/4 MS营养液的黄瓜幼苗中,共培养7 d后观察其地上部和地下部的生长状况,结果如图5所示。结果表明,M95和WT均能在黄瓜根系上定殖,并促进黄瓜幼苗地上和地下部的生长,且在水培条件下,M95对黄瓜幼苗的促生效果比WT更好(图5-A)。对地上部而言,M95对黄瓜幼苗株高和叶面积的作用最为显著,与CK相比分别增加了57.93%和28.21%,与WT相比分别增加了14.79%和16.84%(图5-B、D)。同时,M95也显著提高黄瓜幼苗地上部的鲜重和干重,其中地上部鲜重比WT提高了30.79%,地上部干重提高了47.61%。因此,M95促进黄瓜幼苗茎的伸长和叶片的伸展,提高其地上部干物质的积累。
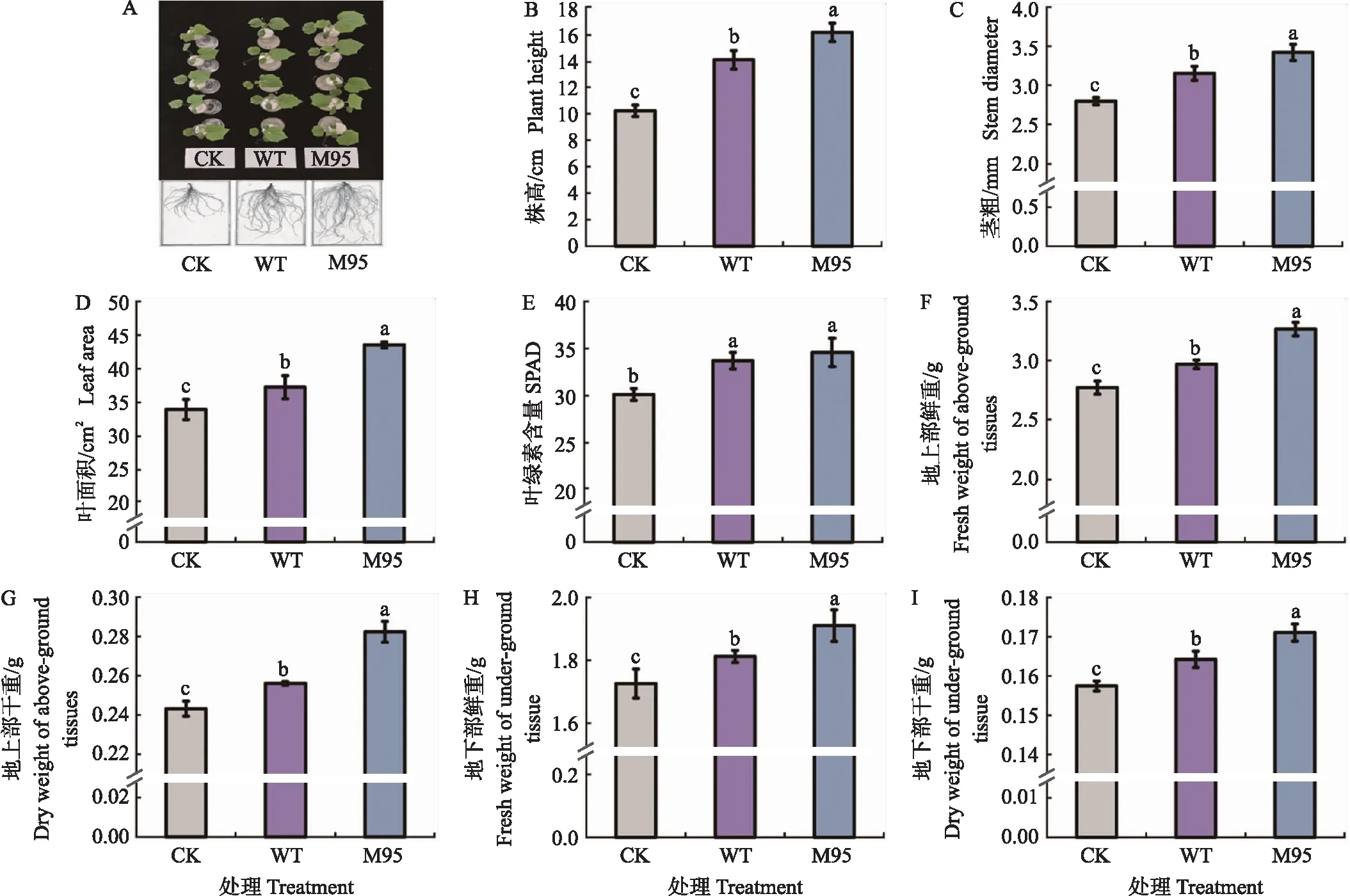
图5 M95和WT在水培条件下对黄瓜幼苗生长的影响Fig.5 The effect of M95 and WT on seedling growth of cucumber in hydroponic experiment1)A. M95和WT在水培条件下对黄瓜幼苗地上部和根系的影响 The effect of M95 and WT on the shoots and the roots of cucumber in hydroponic experiment;B-I. M95和WT在水培条件下对黄瓜幼苗生物指标的影响 The effect of M95 and WT on the biological indicators of cucumber in hydroponic experiment. 2)CK:空白对照 Blank control;WT:添加野生株NJAU 4742的处理 The treatment with wide strain NJAU 4742;M95:添加突变株M95的处理 The treatment with mutant M95. 3)不同小写字母表示在0.05水平差异显著。下同。Different lowercase letters indicate significant differences at 0.05 level. The same as follows.
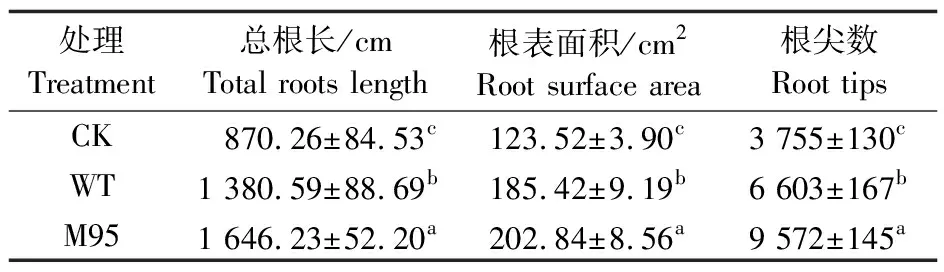
表3 M95和WT在水培条件下对黄瓜根系的影响Table 3 The effect of M95 and WT on the cucumberroot in hydroponic experiment
M95和WT对黄瓜幼苗地下部的影响均大于地上部。M95处理中黄瓜地下部干重为CK处理的2.83倍,比WT处理增加了0.48倍(图5-I)。同时,比较了M95和WT菌株在水培条件下对黄瓜幼苗根系的影响。结果(表3)表明,二者均可显著促进黄瓜根系的生长和侧根的发生,其中M95处理显著增加黄瓜幼苗根尖数,与CK相比提高了1.55倍,与WT处理相比提高了0.45倍。M95处理对黄瓜总根长和根表面积的影响也十分显著,与CK相比分别提高了89.17%和64.23%,比WT处理分别提高了19.24%和9.40%。上述结果表明,在水培条件下M95对黄瓜幼苗的促生作用显著优于WT,主要体现在提高了植株在自然状态下的含水量和干物质的积累。同时,与WT相比M95显著促进了黄瓜根的伸长和侧根的发生,增加了植株的根表面积,有利于其对营养物质的吸收和运输,从而促进地上部的生长。
2.6 黄瓜盆栽试验结果
从表4可知:M95处理和WT处理黄瓜地上部及地下部的干/鲜重均显著高于CK,且M95处理显著高于WT处理。与CK相比,M95处理黄瓜地上部鲜重和干重分别提高了46.70%和72.27%,地下部鲜重和干重分别提高了69.29%和95.26%;M95处理黄瓜地上部鲜重和干重是WT处理的1.13和1.22倍,而地下部鲜重和干重分别为1.18和1.34倍。为了验证M95的促生效果,在相同条件下完成了第2季的盆栽试验,第2季盆栽试验结果与第1季结果呈现出较好的一致性。上述的研究结果进一步证明,在土壤环境中M95对黄瓜的促生效果依然显著高于WT。
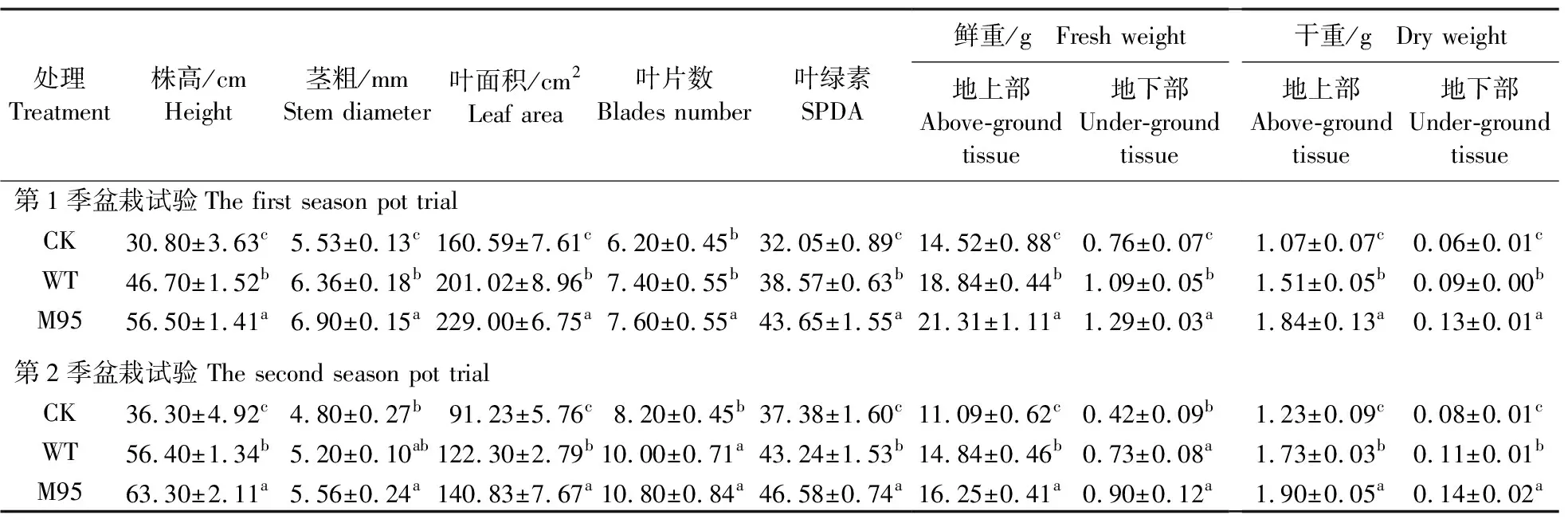
表4 2季盆栽试验中不同处理黄瓜植株各生物指标Table 4 The biological indexes of cucumber plants with different treatments in two season pot trials
2.7 不同施肥处理对黄瓜根际微生物数量的影响
从图6可知:M95处理和WT处理均能显著增加根际土壤中总真菌和总木霉的数量,但只有施用M95生物有机肥能显著增加根际土壤中总细菌的数量,其中细菌数量显著高于真菌数量。M95处理中根际的真菌数量最高,是CK的7.73倍,比WT处理增加了189.94%;同时,M95处理中木霉属的数量占总真菌数量的0.016%,比对照组增加了11.96倍,但与WT处理相比并无显著差异。与CK相比,WT处理的根际细菌数量并无显著差异,但M95处理的根际中细菌数量却显著增加。上述结果进一步说明M95和WT均能在植物根际生存并大量繁殖,且能提高根际土壤微生物多样性;同时,M95处理对土壤微生物数量的影响显著大于WT处理,这有利于更多的根际有益微生物的生长进而协同促进黄瓜生长。
3 结论与讨论
IAA可以通过加速顶端优势的形成、侧根和不定根的形成、调节根形态、促进细胞分裂和细胞伸长等来介导根结构的变化,从而起到良好的促生作用[19-20]。除植物本身外,包括细菌和真菌在内的多种微生物也可以合成并分泌IAA。已有研究表明,外源合成的IAA在一定的浓度下同样可以起到促进植株生长的作用[21]。史娟等[22]探究哥斯达黎加链霉菌(Streptomycescostaricanus)菌株A-m1发现,其IAA产量达1.04 μg·mL-1,对番茄幼苗具有显著的促生作用;宋欢等[23]发现从雷公藤中分离出来的内生真菌NS33具有较高的IAA分泌能力,能够显著促进雷公藤组培苗生长;陆妍吉等[24]从白菜连作土壤中分离获得的阿耶波多氏芽胞杆菌(Bacillusaryabhattai) 和特基拉芽胞杆菌(Bacillustequilensis)均能分泌IAA,从而对白菜幼苗起到不同程度的促生作用。本研究通过X射线照射突变的方法获得了IAA高产突变株M95,其IAA产量比WT提高了20.51%,这可能是M95在水培和盆栽试验中对黄瓜幼苗的促生效果比WT均有显著提高的原因之一。
真菌中IAA生物合成的途径已有较为广泛的研究。已知生长素的生物合成途径分为色氨酸依赖途径(Trp)和非色氨酸依赖途径两大类,而包括木霉在内的真菌主要通过色氨酸依赖途径合成IAA。色氨酸依赖途径包括吲哚丙酮酸途径(IPA)、吲哚乙酰胺途径(IAM)、吲哚乙腈途径(IAN)和色胺途径(TAM)[25]。已有研究发现,稻瘟病菌(Magnaportheoryzae)和油菜黑胫病菌(Leptosphaeriamaculans)均可通过IPA途径合成IAA。刘秋梅[26]研究发现,NJAU 4742可以通过IPA、IAM、IAN和TAM 4种途径合成IAA。通过蛋白组学鉴定和荧光定量PCR试验筛选,发现木霉生长素生物合成中的关键基因为吲哚乙酰胺水解酶基因(iaaH2)、腈水解酶基因(nit1和nit2)和吲哚乙醛脱氢酶基因(ald1和ald2)。本研究发现,与野生株WT相比,M95的iaaH2基因和nit2基因的表达水平均大幅提高,分别为WT的32.68和14.58倍。该结果进一步说明腈水解酶和吲哚乙酰胺水解酶基因转录水平的提高,是M95产IAA能力增强的重要原因,但是nit2和iaaH2转录水平的提高是由哪个基因突变导致的目前还尚未可知,有待后续更深入的研究。
植株的干/鲜重体现了植株干物质的积累及其含水量,是反映植株生长情况的重要指标之一。本研究发现,无论是水培还是盆栽试验条件下,M95都能显著提高黄瓜幼苗的各项生物指标(株高、茎粗、叶面积和干/鲜重),对地下部干重的影响也较显著。这说明M95对植株的作用首先体现在根系上,具体表现为促进根的伸长以及侧根的发生,从而促进植株根系对水分以及养分的吸收,进而促进地上部的生长。本实验室前期研究发现,NJAU 4742可以通过调节与寄主根系相互作用过程中的不同代谢产物和信号物质来加速侧根形成并调节根系形态,从而增加植株生物量[11],这与本研究结果一致,且本研究中M95对黄瓜根系的促生作用显著强于WT。另外,有研究表明,根际微生物功能和土壤微生物群落介导的有机物转化和养分循环过程能够提高植物对养分的吸收利用效率,也会反过来影响微生物群落结构和活动[27]。方伟等[28]研究发现施用生物有机肥能显著改变土壤中细菌和真菌的丰度,加速土壤碳氮循环,提高土壤养分的有效性,从而促进植物生长。姚有华等[29]分别用芽胞杆菌(Bacillussp.)TSXJ4和青霉菌(Penicilliumsp.)GSXJA对青稞种子进行浸种处理后再栽培,发现根际土壤中的细菌及真菌群落结构发生了显著变化,主要体现在其稳定性提高,土壤肥力显著增加,促进了青稞的生长。本研究发现,添加NJAU 4742和M95生物有机肥均对土壤微生物群落产生了一定的影响,且M95能够显著增加土壤细菌和真菌的数量。这可能是因为M95影响了黄瓜根系化感物质的分泌,这些物质在土壤中积累,使得土壤环境发生了改变,更有利于细菌和真菌的生长,导致土壤微生物数量增加。
