高温下不同施肥量对水稻产量品质形成的影响及其与能量代谢的关系分析
2023-05-18王文婷马佳颖李光彦符卫蒙李沪波林洁陈婷婷奉保华陶龙兴符冠富秦叶波
王文婷 马佳颖 李光彦 符卫蒙 李沪波 林洁 陈婷婷 奉保华 陶龙兴 符冠富 秦叶波
高温下不同施肥量对水稻产量品质形成的影响及其与能量代谢的关系分析
王文婷1, #马佳颖1, #李光彦1, 3符卫蒙1李沪波1林洁1陈婷婷1奉保华1陶龙兴1符冠富1秦叶波2, *
(1中国水稻研究所, 杭州 311401;2浙江省农业技术推广中心, 杭州 310020;3扬州大学, 扬州 225009;#共同第一作者;*通信联系人, email: qyb.leaf@163.com)
【目的】近年来极端高温天气频发,严重抑制水稻产量品质形成,优化植株营养状态,改善能量代谢可减缓高温热害,但高温下不同施肥量对籽粒能量代谢的影响及其与产量、品质及耐热性形成的影响仍未见报道,其作用机制的阐明可为水稻耐热性抗风险栽培技术研发提供重要理论依据。【方法】以浙江省推广大面积较大的单季杂交籼稻品种中浙优8号为材料,采用人工气候室盆栽的试验方法开展研究。设置不施肥(0-NPK)、1/2肥料(1/2-NPK)和正常施肥(1-NPK)三个肥料用量处理,开花当天移至人工气候室高温处理15d(高温处理设置昼/夜分别为36/28℃,常温对照设置昼/夜分别为28/25℃)。【结果】无论常温或高温下,在供试肥料水平下随着施肥量增加,中浙优8号产量、结实率及千粒重均呈逐渐增加的趋势;高温下,结实率、千粒重的下降幅度随施肥量增加而下降。然而,整精米率随施肥量增加而下降,垩白度则呈增加的趋势。高温处理后籽粒可溶性糖、MDA、H2O2、腺苷三磷酸酶(ATPase)及PARP [poly(ADP-ribose) polymerase]含量高于常温对照处理,淀粉、非结构性碳水化合物、抗氧化酶活性及ATP含量则低于常温对照。无论常温或高温下非结构性碳水化合物、抗氧化酶活性、ATP及ATPase均随着施肥量的增加而提高,而MDA、H2O2及PARP含量则呈下降趋势。常温下,能量感受器基因及相对表达量随施肥量增加而下降,而则呈增长趋势。与常温对照相比,高温处理后上调表达而相对表达量随施肥量增加而上调【结论】增施肥料能有效缓解花期高温对水稻结实率及千粒重的影响,但外观品质与加工品质存在变劣的趋势。常温下,增施肥料品质变劣可能与籽粒灌浆速率加快有关,而高温导致品质下降主要在于能量不足,难以满足产量及品质形成的需求。
水稻;高温热害;施肥量;产量品质;能量代谢
近几十年来,随着全球工业化进程,人类活动对环境的影响日益扩大,温室气体肆意排放引发的温室效应加剧了全球气候变暖,极端高温天气发生的频率、强度和持续时间均呈增加趋势[1]。根据国家气候中心监测评估,综合考虑高温热浪事件的平均强度、影响范围和持续时间,2022年夏季的区域性高温事件综合强度已达到1961年有完整气象观测记录以来最强。水稻(L.)作为世界上最重要的粮食作物之一,对我国乃至全球粮食安全生产意义重大[2-4]。然而,水稻生殖生长阶段对温度比较敏感,此期发生极端高温天气将导致水稻大幅度减产,因而水稻高温热害多年来一直是研究的热门课题。
水稻的高温伤害程度因时期而异。高温对水稻营养生长的影响较小,除非长时间发生45℃以上的高温天气[5];抽穗开花期最高温度38 ℃以上,持续3~5 d,将导致小穗育性大幅度下降[6];稻米品质形成的最佳温度籼稻为21℃~25℃,粳稻为21℃~24℃,温度过高或过低,均会影响到稻米品质的形成[7]。一般情况下,水稻花期高温可抑制花药开裂、花粉萌发及花粉管在雌蕊组织的伸长,致使小穗败育;籽粒灌浆期高温可干扰籽粒植物激素代谢,降低籽粒灌浆关键酶活性,导致籽粒充实受阻、垩白度增加、整精米率及食味品质下降。Zhang等认为开花期高温不仅导致小穗败育,还会抑制同化物转运,进而影响到籽粒充实及最终粒重的形成,糖信号传递受阻是花期高温导致“源-库-流”关系失衡的主要因素[8]。因而,外源蔗糖可一定程度减缓高温热害对水稻籽粒充实的影响[9]。另外,外源脱落酸、水杨酸均能有效提高水稻穗期耐热性[10-12]。虽然水稻高温热害的研究较多,但在生产中所能采用的有效措施仍然比较有限,因而水稻对高温热害的响应机理及其应对途径还需要进一步研究,尤其在能量代谢方面。
能量是一切生命活动的基础,而能量代谢是指生物体通过物质代谢,从外界摄取糖类、脂类及蛋白等营养物质,经分解吸收将其化学能释放转化为生物体可以利用的能量,该过程包括能量的释放、转移、贮存和利用[13]。水稻产量、品质及耐热性的形成均需要消耗大量能量,因而高温干扰能量平衡及代谢[9,14],致使能量生产及利用效率下降,难以同时满足产量、品质及耐热性对能量的需求,最终导致产量下降、品质变劣[15]。有鉴于此,能促进能量产生效率,减少能量消耗,提高水稻产量、改善稻米品质的高温热害减缓技术研究亟待进行。氮、磷、钾是水稻生长发育过程中不可缺少的营养元素,其丰缺度直接影响着水稻生理特性、生化代谢、养分协同吸收、最终产量品质的形成[16,17]。优化水稻植株营养,不仅提高肥料利用效率,还能有效提高水稻抗逆性。据报道,高温下生物炭配合磷施用能提高水稻产量及稻米品质[18]。另外,氮素在稻米品质应对高温热害中也发挥了重要作用,但结果不尽相同[19-21]。研究表明,氮、磷、钾元素主要通过影响光合作用、呼吸作用、电荷平衡和磷酸化调节能量代谢;其中以氮素的影响最大,氮素缺乏不仅抑制水稻植株能量的产生,还加剧能量消耗;其次分别为磷和钾[22];植物磷缺乏会改变光合作用和碳代谢,ATP合成受阻,最终限制生物量积累和产量形成[23,24];钾元素虽然不与共价有机分子结合,但在渗透调节、电荷平衡、稳态酶激活、膜电位和膜蛋白转运过程中起着至关重要的作用[25,26]。虽然有关氮、磷、钾元素影响水稻生长发育的研究众多,但其含量在籽粒中的变化特征与能量代谢及耐热性关系的研究还比较缺乏;另外,目前的研究主要以N或P或K元素为主,高温下不同施肥量对籽粒能量代谢的影响及其产量品质形成的研究较少。鉴此,本研究以中浙优8号为试验材料,设置不同施肥量处理,研究高温下施肥量对水稻产量、品质、千粒重、结实率、干物质积累及分配、抗氧化物酶活性、碳水化合物代谢及能量代谢的影响,明确不同施肥量在水稻产量、品质及耐热性形成中的作用及其与能量代谢的关系。
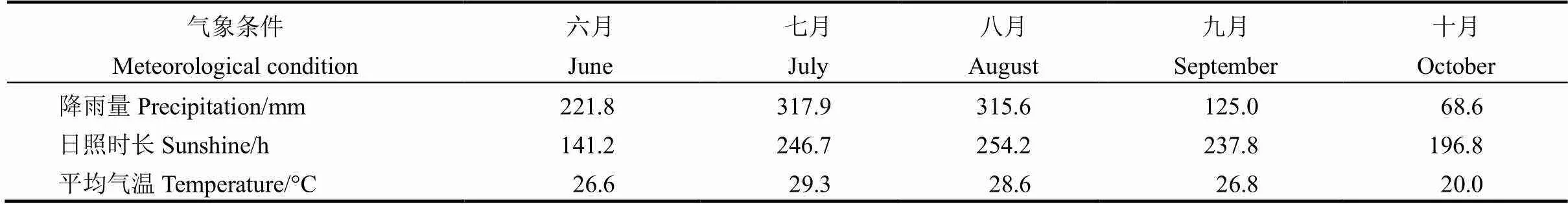
表1 水稻生长期降雨量、日照时长以及平均气温的变化
六月降雨量、日照时长以及平均气温从6月10日开始计算。
Precipitation, sunshine hours and average temperature in June were recorded from June 10 to 30.
1 材料与方法
1.1 试验材料
本研究在浙江省杭州市中国水稻研究所富阳试验基地进行,水稻材料为籼型杂交稻中浙优8号。种子经浸种催芽后播于水稻秧盘中,在自然状态下生长至3叶1心时移栽至盆栽桶内(高20 cm×直径19 cm),采用随机区组设计,每桶4株均匀分布,每处理设置5次重复。盆栽土为稻田黏壤土,pH值为6.81,有机质含量为41.98 g/kg,碱解氮135.89 mg/kg,有效磷34.36 mg/kg,速效钾105.13 mg/kg。风干后粉碎,每盆约5 kg干土,混匀装盆加水浸泡24 h后移栽。实验设置了3个肥料用量处理,分别为不施肥(0-NPK),1/2正常肥料用量(1/2-NPK)和正常施肥(1-NPK)。正常施肥量为氮肥(尿素0.4907 g/桶,含氮量46%),按底肥∶分蘖肥∶穗肥=2∶2∶1的比例分期施用。磷肥(2.35 g过磷酸钙/桶,P2O5≥12%)以于底肥一次性施用,钾肥(氯化钾0.38 g/桶,K2O≥60%)按底肥∶分蘖肥=1∶1的比例分期施用。其余梯度按比例缩减。水稻开花当天移至人工气候室进行高温处理,高温处理昼/夜温分别为36/28℃,常温对照昼/夜温分别设置为28/25℃,相对湿度昼夜分别为80%和70%,自然光照条件(平均光照强度为1000 µmol·m−2·s−1),高温处理15 d。高温结束后移至自然条件下生长至成熟。
1.2 测定项目与方法
1.2.1 产量、结实率、千粒重
水稻成熟收获后,将籽粒烘干至恒重。通过考查实粒数、秕粒数及空粒数,计算结实率及千粒重和产量。
1.2.2 整精米率和垩白度
每个样品设定3个重复,每个重复稻谷重不低于130g,利用砻谷机(大竹otake-FC2R, 日本)脱壳后利用精米机(yamamoto山本VP-32,日本)研磨,测定整精米率;使用大米外观品质检测分析仪(万深SC-E,杭州万深检测科技有限公司)测定垩白度。
1.2.3 碳水化合物
可溶性糖及淀粉的含量采用蒽酮比色法[27]测定。非结构性碳水化合物(NSC)为可溶性糖和淀粉含量之和。
1.2.4 抗氧化酶活性和H2O2、MDA含量
超氧化物歧化酶(SOD)活性采用Giannopolitis等[28]的方法测定。过氧化物酶(POD)活性采用Maehly 等[29]的方法。过氧化氢酶(CAT)活性采用Zhang 等[11]方法测定。抗坏血酸过氧化物酶(APX)活性测定采用Bonnecarrere 等[30]的方法。H2O2含量的测定采用Brennan等[31]的方法,稍作修改。将约0.2 g冷冻叶片于研钵中加入4 mL 10 mmol/L 3-氨基-1,2,4-三唑磨成匀浆,6000 r/min离心15 min,取上清液。2 mL上清液中加入1 mL 0.1% TiCl4(溶于20% H2SO4),混匀后静置5 min,6000 r/min下离心10 min弃去沉淀,在410 nm处测定上清液的吸光度,并制作标准曲线,根据标准曲线求得样品中H2O2含量;根据Dhindsa等[32]的方法测定MDA含量,MDA浓度计算公式为:C(μmol/L) =6.45×(OD532−OD600)−0.56×OD450。根据上述公式求得样品MDA浓度后,计算样品中MDA的含量。
1.2.5 ATP、ATPase和PARP含量
齐穗后10 d和齐穗后20 d取水稻籽粒用于能量代谢相关指标测定。ATP、ATPase和PARP均根据试剂盒说明(Dogesce)采用酶联免疫吸附法(ELISA)测定。取0.1 g鲜叶加入1 mL 0.1 mol/L的磷酸缓冲液(PBS, pH7.4),冰浴充分研磨成浆,3000×下离心20 min。收集上清液于450 nm波长下测定。
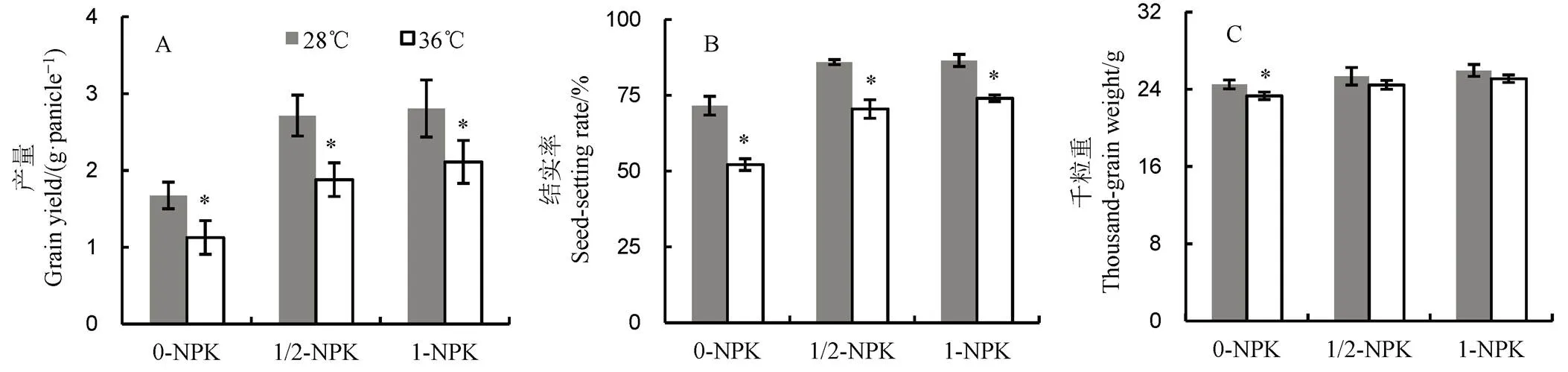
0-NPK、1/2-NPK和1-NPK分别表示NPK施肥量分别为0、1/2正常用量和正常用量。*表示t检验比较分析显著差异。下同。
Fig. 1. Effects of different fertilizer application on grain yield, seed setting rate and thousand-grain weight of rice under heat stress.
1.2.6 实时荧光定量分析
用TRIpure试剂(Aidlab biotenologies, 北京)从100 mg叶片中提取总RNA。利用TOYOBO反转录试剂盒(ReverTra-Ace-qPCR-RT-Master Mix)将RNA转化为第一链cDNA。以SYBR-Green-I (TOYOBO)为荧光标记,将得到的cDNA作为定量PCR扩增的模板,采用两步法在ABI StepOneplus荧光定量仪上运行。
1.2.7 氮、磷、钾含量测定
将水稻样品置于烘箱中105℃杀青30min,60℃干燥24 h。用粉碎机研磨后,将约0.2 g的样品放入消化管中,加入CuSO4-Na2SO4(1∶10)催化剂和浓硫酸。氮和磷含量采用全自动间歇化学分析仪(SmartChem450,AMS,Westco,意大利)测定,总钾含量采用H2SO4-H2O2消化火焰光度计法测定。
1.3 数据分析
采用SPSS软件(11.5,IBM Corp.,Armonk,NY,USA)进行方差分析,用Excel 软件进行数据处理与作图。
2 结果与分析
2.1 高温下不同施肥量对产量、结实率及千粒重的影响
由图1可见:1)增施肥料能提高水稻单穗产量。常温下,与0-NPK处理相比,1/2-NPK和1-NPK处理增产幅度分别为62.49%和68.04%,差异达显著水平;高温下,1/2-NPK和1-NPK处理的产量分别比0-NPK增加66.24%和86.67%;高温处理后水稻产量的降幅随施肥量增加而下降,降幅分别为32.68%,30.77%和24.83%(图1-A);2)高温胁迫显著降低水稻结实率,降幅随施肥量增加呈下降趋势,与常温对照处理相比,0-NPK、1/2-NPK及1-NPK处理的降幅分别为27.15%、17.97%和14.40%(图1-B);3)施肥量增加有助于提高千粒重。常温下,0-NPK,1/2-NPK和1-NPK处理的千粒重分别为24.50 g、25.34 g和25.95 g,高温处理后则分别为23.31 g、24.45 g和25.09 g;与常温对照相比,0-NPK、1/2-NPK和1-NPK处理降幅分别为4.86%、3.51%和3.31%(图1-C)。
2.2 高温下不同施肥量对籽粒垩白度和整精米率的影响
由图2可见:1)增施肥料增加水稻籽粒垩白度。常温下,与0-NPK处理相比,1/2-NPK和1-NPK处理的籽粒垩白度分别增加1.82%和11.56%,而高温处理后则分别上升了0.46%和6.01%。与常温对照相比,高温下0-NPK、1/2-NPK和1-NPK处理垩白度依次增加了170.26%、110.55%和22.25%;2)整精米率随肥料施用量增加而下降。常温下,与0-NPK处理相比,1/2-NPK和1-NPK处理的籽粒整精米率分别降低了7.61%和7.76%,而高温处理后则分别下降了14.14%和14.50%。与常温对照相比,0-NPK、1/2-NPK和1-NPK的整精米率的降幅依次为16.90%、19.99%和20.13%。
2.3 高温下不同施肥量对籽粒碳水化合物含量的影响
由图3可见:1)无论常温或高温处理,籽粒中可溶性糖、淀粉及非结构性碳水化合物含量均随肥料使用量的提高而增加;2)与常温对照相比,高温下0-NPK、1/2-NPK和1-NPK处理的可溶性糖含量分别提高28.39%、10.76%和8.89%,而淀粉含量则分别降低了11.52%、9.07%和9.37%;3)高温下,非结构碳水化合物含量显著低于常温处理,其中0-NPK处理下降幅度最大,比28℃处理降低了7.35%,而1/2-NPK和1-NPK处理分别降低6.76%和7.26%。
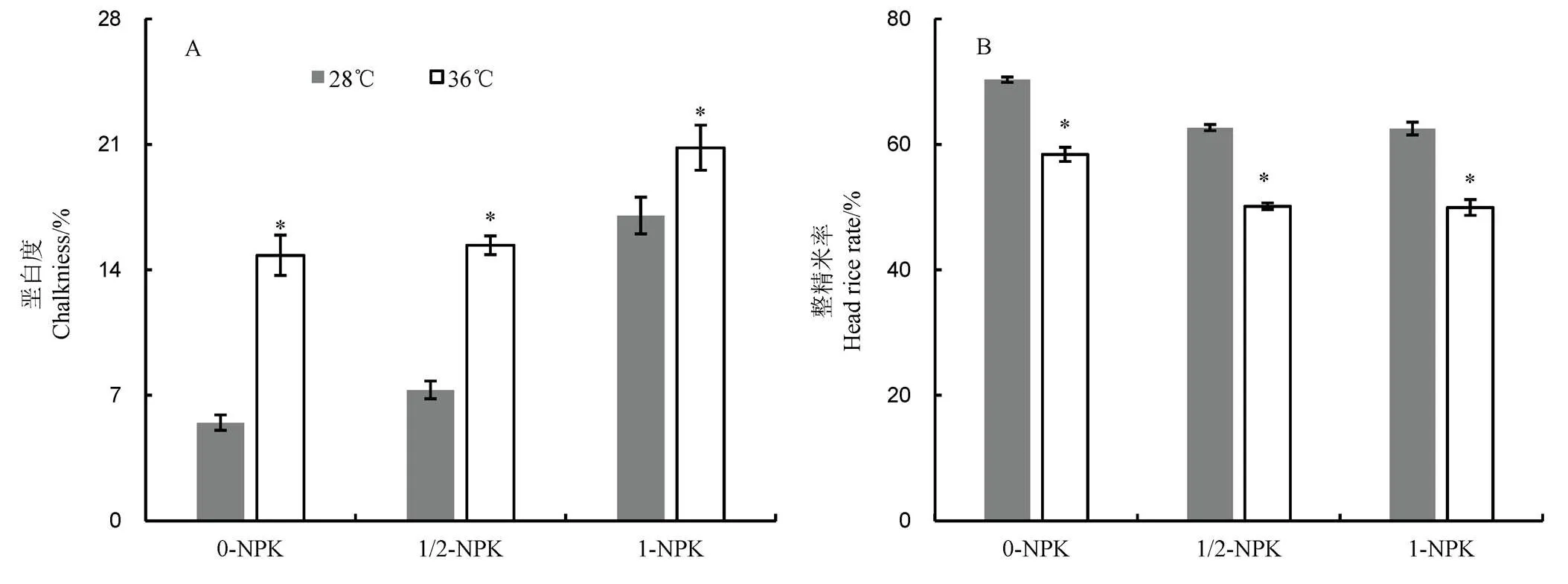
图2 高温下不同施肥量对水稻垩白度和整精米率的影响
Fig. 2. Effects of different fertilizer application levels on chalkiness and head rate of rice under heat stress.
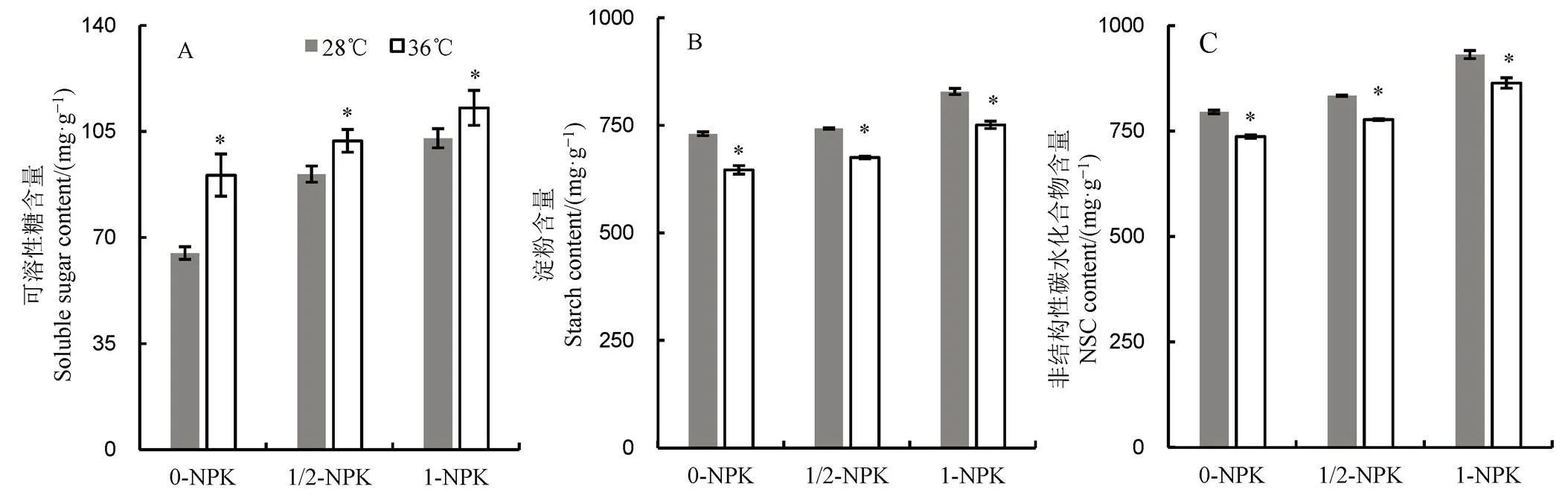
图3 高温下不同施肥量对水稻碳水化合物的影响
Fig. 3. Effect of different fertilizer application levels on carbohydrate contents in rice under heat stress.
2.4 高温下不同施肥量对籽粒中H2O2及MDA含量的影响
由图4可见:1)无论常温或高温处理,籽粒中的H2O2和MDA含量均随着施肥量增加而下降;2)高温下,不同处理的籽粒H2O2含量均显著增加,增加幅度分别为14.95%、29.11%和9.87%;3)和常温对照相比,高温下0-NPK、1/2-NPK和1-NPK处理的MDA含量均有所增加,其中以0-NPK处理的增幅最大,达19.29%,差异达显著水平,而1/2-NPK和1-NPK处理差异不明显。
2.5 高温下不同施肥量对籽粒中抗氧化酶活性的影响
由图5可见:1)无论常温或高温处理,籽粒SOD、POD、CAT及APX活性均随施肥量增加而提高。和常温相比,高温处理后籽粒抗氧化酶活性均呈下降的趋势,其中CAT和APX活性下降幅度最大,在三种肥料处理下差异均达到显著水平;2)与常温对照相比,高温下1-NPK处理的SOD活性降幅最小,差异未达显著水平,而0-NPK和1/2-NPK处理的降幅依次为 35.07%和24.77%,差异均达显著水平;3)POD活性以0-NPK处理的降幅最大,差异达显著水平,而1/2-NPK和1-NPK处理下,常温和高温处理间差异不显著;4)与常温对照相比,高温下0-NPK、1/2NPK和1-NPK处理的POD活性分别下降了23.56%、20.87%和29.63%,而APX的降幅依次为38.36%、34.25%和20.47%。
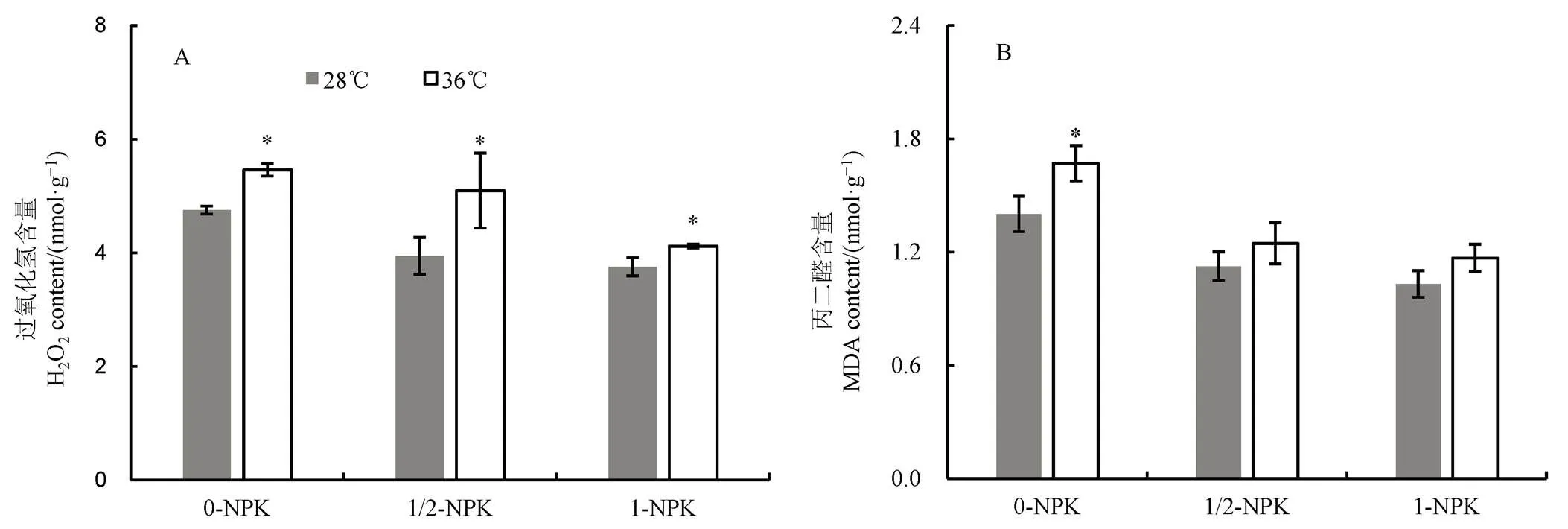
图4 高温下不同施肥量对籽粒H2O2和MDA含量的影响
Fig. 4. Effects of different fertilizer application levels on contents of MDA and H2O2in grains of rice under heat stress.
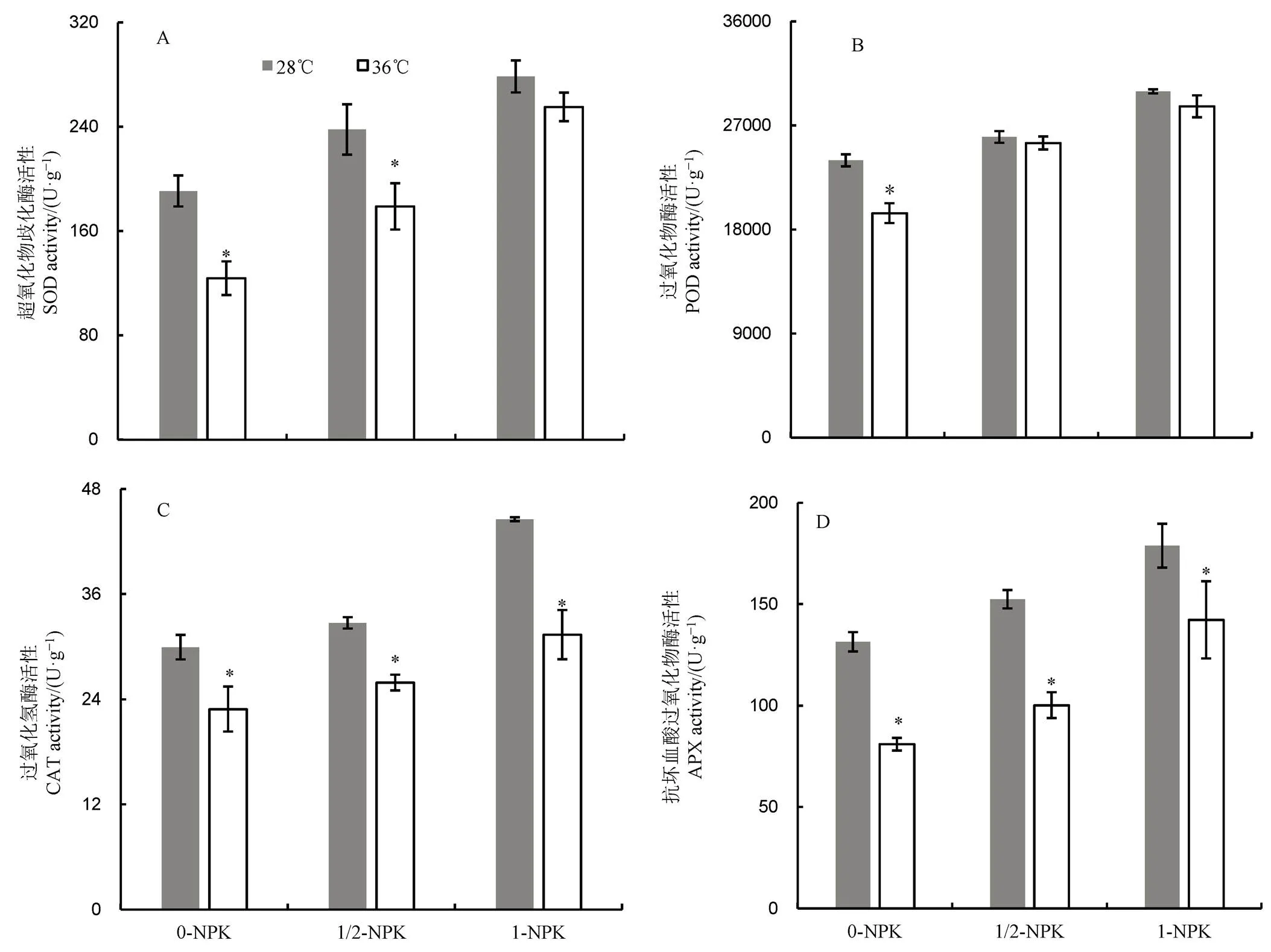
图5 高温下不同施肥量对水稻籽粒抗氧化酶活性的影响
Fig. 5. Effect of different fertilizer application levels on antioxidants enzyme activities of grains under heat stress.
2.6 高温下不同施肥量对籽粒中能量状态的影响
由图6可见:1)无论是常温或是高温下,籽粒中的ATP和ATPase含量均随肥料使用量增加呈增长的趋势。常温下,与0-NPK处理相比,1/2-NPK和1-NPK处理的ATP含量分别增加了8.88%和13.22%,而在36℃高温处理条件下则分别上升了2.72%和9.74%;2)与对照相比,高温下不同施肥处理的ATP和ATPase含量均显著下降;0-NPK,1/2-NPK和1-NPK处理的ATP含量分别降低9.68%、15.41%和13.17%,而ATPase含量则依次为7.72%、7.96%和11.00%;2)两种温度处理下,PARP含量随着施肥量的增加而下降。常温下,与0-NPK处理相比,1/2-NPK和1-NPK处理的PARP含量分别降低了16.50%和17.94%,而高温下的降幅分别为5.80%和6.28%。与常温相比,高温处理后0-NPK、1/2-NPK和1-NPK的PARP含量分别增加了2.48%、15.38%和16.87%。
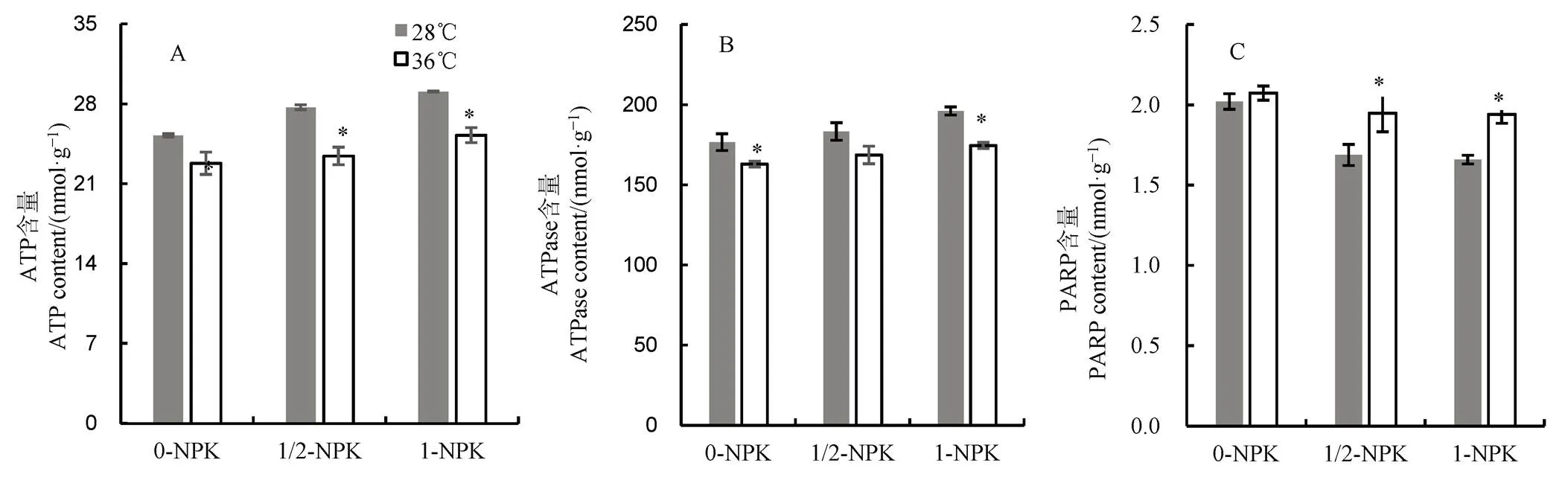
图6 高温下不同施肥量对水稻籽粒能量状态的影响
Fig. 6. Effect of different fertilizer application levels on energy states in grains under heat stress.
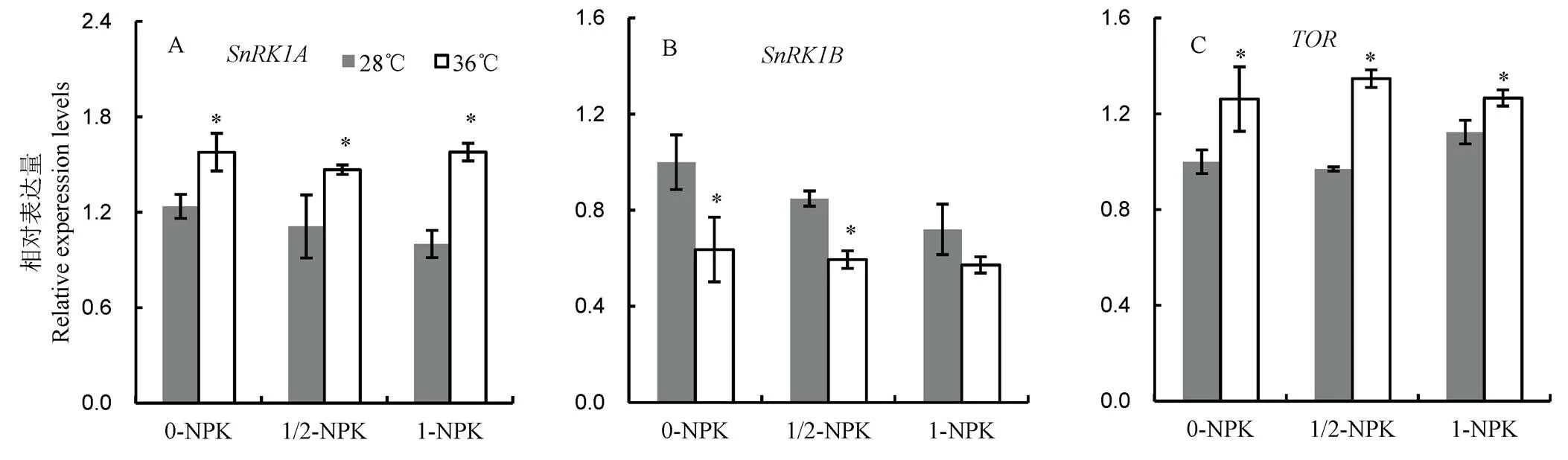
图7 高温下不同施肥量对水稻籽粒能量分配的影响
Fig. 7. Effects of different fertilizer application levels on energy allocation in rice grains under heat stress.
2.7 高温下不同施肥量处理对籽粒中能量分配的影响
由图7可见:1)常温下,和的相对表达量均随着施肥量增加而下降。然而,对高温的响应不一致。高温处理后,不同处理的的表达均显著增加,0-NPK、1/2-NPK和1-NPK处理的增幅分别为27.6%、32.1%和57.9%;的表达量则几乎显著低于常温对照处理,降幅分别为36.4%、29.9%和20.5%。2)基因同样受高温诱导,高温处理后,0-NPK,1/2-NPK和1-NPK处理的表达量均显著高于相应的常温处理,增幅分别为26.2%、39.0%和12.7%。
2.8 高温下不同施肥量对水稻氮、磷、钾含量的影响
由图8可见:1)常温条件下,水稻茎、叶和穗中的全氮含量均随肥料使用量增加呈增长的趋势。与0-NPK处理相比,1/2-NPK和1-NPK处理茎鞘全氮含量分别增加9.63%和24.99%,叶片全氮含量分别增加13.31%和36.98%,而在穗中则分别为5.13%和14.99%;2)与28℃处理相比,高温处理后茎鞘全氮含量均有所下降,叶中的全氮含量上升,尤其是1/2-NPK处理,在不同温度间差异显著。3)常温条件下,水稻茎中全磷含量随肥料使用量增加呈下降的趋势,而在高温条件下却呈现相反的趋势。与0-NPK处理相比,常温条件下1/2-NPK处理全磷含量降低了10.53%,而高温条件下则上升了23.31%;4)无论是常温或是高温下,叶片中全磷含量在1/2-NPK处理中显著高于0-NPK和1-NPK处理;5)高温条件下,穗中全磷含量在不同施肥量下没有差异,但在常温条件下,穗中含磷量随施肥量增加而增加,在1/2-NPK和1-NPK处理中分别提高了40.52%和77.88%。6)无论是常温或是高温下,茎中的全钾含量均随肥料使用量增加而有所上升,但差异不显著。在0-NPK处理下,高温处理全钾含量显著降低,降幅为9.38%;7)叶片中全钾含量在两种温度处理条件下变化不大,但是1-NPK处理显著高于0-NPK和1/2-NPK处理,28℃下增幅分别为35.43%和26.93%,而在36℃下则分别上升了33.64%和30.03%;8)高温条件下,不同肥料处理间,穗中全钾含量差异不显著,而在常温条件下,随肥料使用量增加,穗中全钾含量显著增加,与0-NPK处理相比,1/2-NPK和1-NPK处理分别增加了40.45%和80.01%。
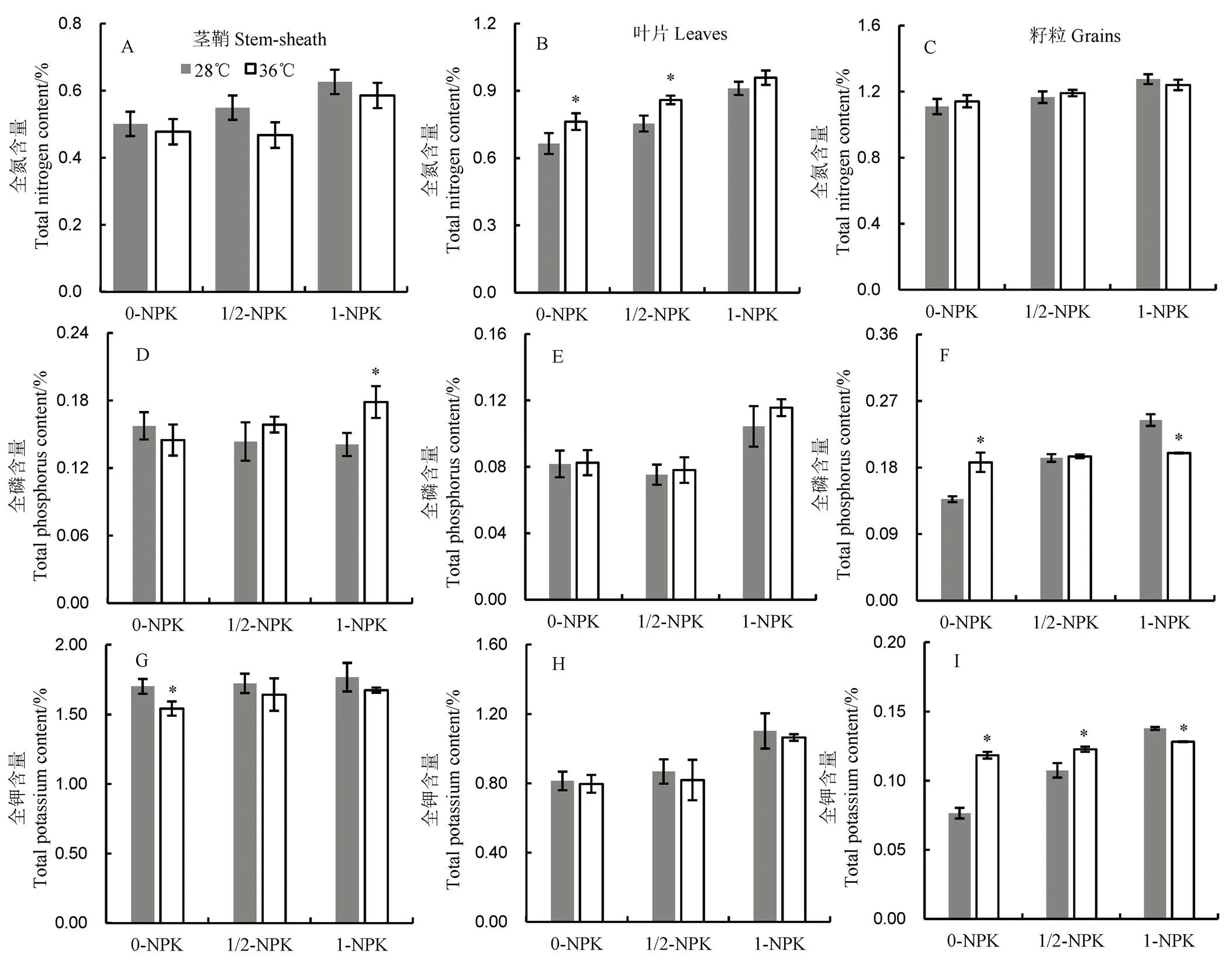
图8 高温下不同施肥量对水稻籽粒氮、磷和钾含量的影响
Fig. 8. Effects of different fertilizer application levels on rice N, P and K contents in rice grains under heat stress.
3 讨论
3.1 高温下不同施肥量对水稻产量形成的影响
已有的研究表明,水稻开花期高温胁迫可抑制颖花及花药开裂,柱头花粉萌发及花粉管伸长受阻,致使小穗败育,进而降低水稻产量[8,9]。本研究结论与其一致,即高温下产量及结实率显著下降,籽粒充实受抑制(图1)。本研究发现随着施肥量的增加,花期高温胁迫对结实率及千粒重的伤害呈逐渐降低的趋势(图1)。赵庆雷等也观察到了类似的现象,即扬花期高温下,水稻结实率随施肥量的减少而降低,其中以无肥处理结实率的降幅最大[33]。另外,前人有关施肥量与水稻耐热性关系的研究表明,随着氮素使用量的增加,高温伤害有所降低[34]。然而,氮素与水稻耐热性之间的关系争议较大,赵决建的研究表明高温下高氮处理结实率明显低于低氮及中氮处理[35]。据报道,中氮处理减少水稻高温热害的效果要明显高于高氮处理[34,36]。高温下氮素对水稻抗逆性影响的结果差异可能与所采用的水稻品种及氮肥使用量不同有关。关于施肥量或氮肥增加提高水稻耐热性作用途径,本研究表明施肥量增加可促进籽粒非结构性碳水化合物积累,增强活性氧清除能力(图3和图5),提高植株抗逆性[37]。对此,缪乃耀等认为施用氮素粒肥显著降低了水稻的叶温和穗温,促进水稻叶片的光合作用,提高抗氧化酶活性,减少MDA积累,增强水稻植株氮代谢,增加硝酸还原酶、谷氨酰胺合成酶活性及氨基酸的含量[38]。另外,高温下增施氮肥可增加叶面积,提高叶片蒸腾速率,降低植株温度[39]。
3.2 高温下不同施肥量对籽粒整精米率和垩白度的影响
稻米品质一般包括碾米品质、外观品质、蒸煮食用品质、营养品质以及特异品质五个方面,主要受环境和遗传因素的影响。研究表明,高温下灌浆期高温使糙米率、精米率、整精米率显著下降,使垩白粒率和垩白度显著升高,导致加工品质和外观品质变差,其中耐热性强的水稻品种的整精米率、垩白度、直链淀粉含量、碱消值和胶稠度等主要米质指标的变幅小于高温相对敏感的品种[40]。另外,通过氮肥处理可有效减小高温胁迫对稻米品质的影响[41]。在本研究中,作者也观察到高温处理后整精米率大幅度下降,而垩百度显著增加,然而施肥量增加则导致品质变劣,表现为垩白度增加,整精米率下降(图2)。虽然前人大多认可增氮能改善稻米的加工品质,但由于品种类型、环境条件试验设计等原因,研究结果并不一致[35]。郭银燕等研究浙江省1995年早稻区试品种碾米品质,认为糙米率、精米率以品种效应为主,整精米率性状以环境效应最大[42]。周培南等认为随施氮量的增加精米率和整精米率降低[43]。在施肥量方面,有研究指出在一定范围内稻米产量随着施肥量的增加而增加,而整精米率随着施肥量的增加而下降[44]。在本研究中,氮磷钾元素同时增加,籽粒氮磷钾元素均随着施肥量的增加而增加,尤其在常温条件下(图8)。然而,和常温对照处理相比,高温处理后籽粒N素含量变化不大,但1-NPK处理中的P和K含量均显著低于对照处理,高温下籽粒中氮磷钾含量的不平衡可能是稻米品质变劣的重要因素,但已有的数据不足以支撑这一假设,因而还需要更进一步研究。
3.3 能量代谢在产量及品质形成中的作用
水稻产量品质形成、逆境响应及同化物的长距离运输均为耗能过程[45,46]。高温等逆境胁迫下,能量不足将抑制花粉萌发和花粉管伸长,同化物转运及代谢受阻,致使籽粒灌浆不充分,限制产量和品质的形成[9]。本研究表明无论常温或高温下,增施肥料能均提高籽粒能量产生效率,表现为ATP和ATPase活性均有增加,而PARP显著下降(图6),与马佳颖等的研究结果比较一致[22]。高温下,籽粒中的ATP和ATPase含量显著低于常温对照处理,而PARP则显著增加,表明和常温对照相比,高温下1/2-NPK和1-NPK处理的能量产生效率明显降低,但能量消耗明显增加(图6)。然而,令人意外的是无论常温或高温,整精米率及垩白度均随着施肥量增加而下降(图2),表明能量代谢在稻米品质形成过程的作用比较复杂,除了ATP含量有关外,也可能与ATP的利用及分配有关[47,48]。据报道,和在能量分配方面发挥重要的作用[48]。与不施肥处理相比,常温下1/2-NPK和1-NPK处理的籽粒和基因相对表达量均有所下降,而则上调表达(图7),表明常温下能量比较充足,籽粒代谢活动比较活跃[47],灌浆速率增加,李志刚等[49]也认为施肥量增加,尤其是粒肥增加能延缓功能叶的衰老,使其在水稻生长后期仍维持较高的光合速率、灌浆速率。相反,高温处理后籽粒中的基因相对表达量增幅随着施肥量提高而增加,的降幅呈下降趋势,而基因的增幅在1-NPK处理中最低(图7),表明高温下施肥量增加有利于优化能量的分配,减少无谓消耗[14]。然而,能量分配的优化不足以弥补高温下能量的不足,可能是导致产量与品质不协同的重要原因[45],类似的研究还未见报道。对此,作者认为常温下籽粒品质变劣,可能与籽粒灌浆速率加快有关,而高温下则可能与能量消耗过大致使能量不足有关。鉴于目前有限的数据不能完全证实这个假设,还需要进一步采用蛋白组学及同化物标记的方法进行验证。
4 结论
本研究表明,增施肥料一方面可以增强抗氧化酶活性增强,从生理上提高水稻抗逆性,缓解高温热害:另一方面可以有效缓解花期高温对水稻结实率及千粒重的影响,但外观品质与加工品质存在变劣的趋势。常温下,增施肥料品质变劣可能与籽粒灌浆速率加快有关,而高温导致品质下降主要在于能量不足,难以满足产量及品质形成的需求。
[1] IPCC. Summary for policymakers//Masson-Delmotte V, Zhai P, Pirani A. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, England: Cambridge University Press, 2021: 1-41
[2] Muthayya S, Sugimoto J D, Montgomery S, Maberly G F. An overview of global rice production, supply, trade, and consumption[J]., 2014, 1324: 7-14.
[3] Bhagirath S, Khawar J, Gulshan M. Rice Production Worldwide[M]. Cham, Switzerland: Springer International Publishing AG, 2017.
[4] Rahaman M M, Shehab M K, Islam A. Total production and water consumption of major crops in south asia during 1988-2013[c]// Conference on Water Security and Climate Change: Challenges and Opportunities in Asia, 28 November - 01 December, 2016. 2016.
[5] Perkins-Kirkpatrick S E, Lewis S C. Increasing trends in regional heatwaves[J]., 2020, 11(1): 3357.
[6] 黄福灯, 李春寿, 刘鑫, 程方民. 高温胁迫对水稻花粉活力的影响[J].浙江农业科学, 2010, (6): 1272-1274.
Huang F D, Li C S, Liu X, Cheng F M. Effects of high temperature stress on pollen viability of rice[J]., 2010, (6): 1272-1274. (in Chinese with English abstract)
[7] 程方民, 刘正辉, 张嵩午. 稻米品质形成的气候生态条件评价及我国地域分布规律[J]. 生态学报, 2002(5): 636-642.
Cheng F M, Liu Z H, Zhang S W. Evaluation on climatic ecological conditions of rice quality formation and regional distribution of our country[J]., 2002(5): 636-642. (in Chinese with English abstract)
[8] Zhang C X, Li G Y, Chen T T, Feng B H, Fu W M, Yan J X, Islam M R, Jin Q Y, Tao L X, Fu G F. Heat stress induces spikelet sterility in rice at anthesis through inhibition of pollen tube elongation interfering with auxin homeostasis in pollinated pistils[J]., 2018, 11(1): 1-12.
[9] Jiang N, Yu P H, Fu W M, Li G Y, Feng B H, Chen T T, Li H B, Tao L X, Fu G F. Acid invertase confers heat tolerance in rice plants by maintaining energy homoeostasis of spikelets[J]., 2020, 43(5): 1273-1287.
[10] 符冠富, 张彩霞, 杨雪芹, 杨永杰, 陈婷婷, 赵霞, 符卫蒙, 奉保华, 章秀福, 陶龙兴, 金千瑜. 水杨酸减轻高温抑制水稻颖花分化的作用机理研究[J]. 中国水稻科学, 2015, 29(6): 637-647.
Fu G F, Zhang C X, Yang X Q, Yang Y J, Chen T T, Zhao X, Fu W M, Feng B H, Zhang X F, Tao L X, Jin Q Y.Effect of salicylic acid on inhibition of spikelet differentiation at high temperature in rice[J]., 2015, 29(6): 637-647. (in Chinese with English abstract)
[11] Zhang C X, Feng B H, Chen T T, Zhang X F, Tao L X, Fu G F. Sugars, antioxidant enzymes and IAA mediate salicylic acid to prevent rice spikelet degeneration caused by heat stress., 2017, 83: 313-323.
[12] Feng B, Zhang C, Chen T, Zhang X, Tao L, Fu G. Salicylic acid reverses pollen abortion of rice caused by heat stress[J]., 2018, 18(1): 245.
[13] Sousa J S, Calisto F, Langer J D, Mills D J, Refojo P N, Teixeira M, Kuhlbrandt W, Vonck J, Pereira M M. Structural basis for energy transduction by respiratory alternative complex III[J]., 2018, 9(1): 1728.
[14] Li G Y, Zhang C X, Zhang G H, Fu W M, Feng B H, Chen T T, Peng S B, Tao L X, Fu G F. Abscisic acid negatively modulates heat tolerance in rolled leaf rice by increasing leaf temperature and regulating energy homeostasis[J]., 2020a, 13(1): 18.
[15] Chen T T, Li G Y, Islam M R, Fu W M, Feng B H, Tao L X, Fu G F. Abscisic acid synergizes with sucrose to enhance grain yield and quality of rice by improving the source-sink relationship[J]., 2019, 19(1): 525.
[16] 孙永健, 孙园园, 徐徽, 杨志远, 秦俭, 彭玉, 马均. 水氮管理模式与磷钾肥配施对杂交水稻冈优725养分吸收的影响[J]. 中国农业科学, 2013, 46(7): 1335-1346.
Sun Y J, Sun Y Y, Xu H, Yang Z Y, Qin J, Peng Y, Ma J.Effects of water and nitrogen management mode and combined application of phosphorus and potassium fertilizer on nutrient uptake of hybrid rice Gangyou 725[J]., 2013, 46(7): 1335-1346. (in Chinese with English abstract)
[17] 姜红芳, 兰宇辰, 王鹤璎, 徐令旗, 李猛, 赵洋, 李晓蕾, 刘旭莹, 吕艳东, 郭晓红. 氮肥运筹对苏打盐碱地水稻养分积累、转运及分配的影响[J].中国土壤与肥料, 2020, (5): 45-55.
Jang H F, Lan Y C, Wang H Y, Xu L Q, Li M, Zhao Y, Li X L, Liu X Y, LY D, Guo X H.Effects of nitrogen fertilizer operation on nutrient accumulation, transport and distribution of rice in soda-alkali soil[J]., 2020, (5): 45-55. (in Chinese with English abstract)
[18] Fahad S, Hussain S, Saud S, Hassan S, Tanveer M, Ihsan M Z, Shah A N, Ullah A, Nasrullah, Khan F, Ullah S, Alharby H, Nasim W, Wu C, Huang J L. A combined application of biochar and phosphorus alleviates heat-induced adversities on physiological, agronomical and quality attributes of rice[J]., 2016, 103: 191-198.
[19] Xu Y Q, Guan X Y, Han Z Y, Zhou L J, Zhang Y, Asad M, Wang Z W, Jin R, Pan G, Cheng F M. Combined effect of nitrogen fertilizer application and high temperature on grain quality properties of cooked rice[J]., 2022, 13: 874033.
[20] Tang S, Zhang H X, Liu W Z, Dou Z, Zhou Q Y, Chen W Z, Wang S H, Ding Y F. Nitrogen fertilizer at heading stage effectively compensates for the deterioration of rice quality by affecting the starch-related properties under elevated temperatures[J]., 2019, 277: 455-462.
[21] Wang X Q, Wang K L, Yin T Y, Zhao Y F, Liu W Z, Shen Y Y, Ding Y F, Tang S. Nitrogen fertilizer regulated grain storage protein synthesis and reduced chalkiness of rice under actual field warming[J]., 2022, 12: 715436.
[22] Ma J Y, Chen T T, Lin J, Fu W M, Feng B H, Li G Y, Li H B, Li J C, Wu Z H, Tao L X, Fu G F.Functions of nitrogen, phosphorus and potassium in energy status and their influences on rice growth and development[J]., 2022, 29(2): 166-178.
[23] Xu H X, Weng X Y, Yang Y. Effect of phosphorus deficiency on the photosynthetic characteristics of rice plants[J]., 2007, 54(6): 741-748.
[24] Chaudhary M I, Adu-Gyamfi J J, Saneoka H, Nguyen N T, Suwa R, Kanai S, El-Shemy H A, Lightfoot D A, Fujita K. The effect of phosphorus deficiency on nutrient uptake, nitrogen fixation and photosynthetic rate in mashbean, mungbean and soybean[J]., 2008, 30(4): 537-544.
[25] Shabala S. Regulation of potassium transport in leaves: from molecular to tissue level[J]., 2003, 92(5): 627-634.
[26] Britto D T, Kronzucker H J. Cellular mechanisms of potassium transport in plants[J]., 2008, 133(4): 637-650.
[27] Yoshida S, Fornd D A, Cock J H, Gomez K A. Determination of sugar and starch in plant tissue[M]//Yoshida S. Laboratory Manual for Physiological Studies of Rice. Los Baños, Philippines: International Rice Research Institute, 1976: 46-49.
[28] Giannopolitis C N, Ries S K. Superoxide dismutases: I. Occurrence in higher plants[J]., 59(2), 1977, 309-314.
[29] Maehly A C, Chance B. The assay of catalases and peroxidases[J]., 1954, 1: 357-424.
[30] Bonnecarrère V, Borsani O, Pedro Díaz, Fabián Capdevielle, Blanco P, Monza J. Response to photoxidative stress induced by cold inrice is genotype dependent[J]., 2011, 180(5): 726-732.
[31] Brennan T, Frenkel C. Involvement of hydrogen peroxide in the regulation of senescence in pear., 1977, 59(3): 411-416.
[32] Dhindsa R S, Pamela P D, Thorpe T A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase[J]., 1981(1): 93-101.
[33] 赵庆雷, 吴修, 王瑜, 李曰鹏, 陈博聪, 王佳, 马加清. 扬花期高温条件下施肥量对水稻高位分蘖及产量性状的影响[J]. 山东农业科学, 2016, 48(8): 61.
Zhao Q L, Wu X, Wang Y, Li Y P, Chen B C, Wang J, Ma J Q. Effects of fertilizer application on high tillering and yield characters of rice at high temperature during flowering stage[J]., 2016, 48(8): 61. (in Chinese with English abstract)
[34] 闫娜. 增施氮素穗肥对幼穗分化期高温下水稻产量的影响及生理机理研究[D]. 武汉: 华中农业大学, 2021.
Yan N.Effects of increased nitrogen panicle fertilizer on rice yield at high temperature during young panicle differentiation and its physiological mechanism[D]. Wuhan: Huazhong Agricultural University, 2021. (in Chinese with English abstract)
[35] 赵决建. 氮磷钾施用量及比例对水稻抗高温热害能力的影响[J]. 土壤肥料, 2005(5): 13-16.
Zhao J J.Effects of application amount and proportion of nitrogen, phosphorus and potassium on resistance to high temperature heat damage of rice[J]., 2005(5): 13-16. (in Chinese with English abstract)
[36] 胡秋倩. 幼穗分化期高温影响水稻产量形成的机理及氮素调控研究[D]. 武汉: 华中农业大学, 2021.
Hu Q Q. Effect of high temperature on rice yield Formation and Nitrogen regulation during young panicle differentiation[D]. Wuhan: Huazhong Agricultural University, 2021. (in Chinese with English abstract)
[37] 孙丽平, 李笑, 苏雪娇. 中华牛肝菌多糖的性质及抗氧化活性[J]. 食品工业科技, 2016, 37(24): 173-175.
Sun L P, Li X, Su X J.Properties and antioxidant activity of polysaccharides from[J]., 2016, 37(24): 173-175.(in Chinese with English abstract)
[38] 缪乃耀. 氮素粒肥对水稻灌浆前期高温胁迫的缓解效应及其生理机制[D]. 南京: 南京农业大学, 2016.
Miao N Y.Alleviating effect and physiological mechanism of nitrogen granule fertilizer on high temperature stress in early filling stage of rice[D]. Nanjing: Nanjing Agricultural University, 2016. (in Chinese with English abstract)
[39] Xiong D L, Yu T T, Ling X X, Fahad S, Peng S B, Li Y, Huang J L. Sufficient leaf transpiration and nonstructural carbohydrates are beneficial for high-temperature tolerance in three rice () cultivars and two nitrogen treatments[J]., 2015, 42(4): 347-356.
[40] 王慧, 张从合, 严志, 申广勒, 周桂香, 杨韦, 方玉, 黄艳玲, 庞战士, 李方宝. 荃两优系列杂交稻品种耐热性与稻米品质的相关性研究[J]. 中国稻米, 2022, 28(4): 79-83.
Wang H, Zhang C H, Yan Z, Shen G L, Zhou G X, Yang W, Fang Y, Huang Y L, Pang Z S, Li F B.Study on the correlation between heat resistance and rice quality of QuanLiangyou Series hybrid rice[J]., 2022, 28(4): 79-83. (in Chinese with English abstract)
[41] 韩展誉, 吴春艳, 许艳秋, 黄福灯, 熊义勤, 管弦悦, 周庐建, 潘刚, 程方民. 不同施氮水平下灌浆期高温对水稻贮藏蛋白积累及其合成代谢影响[J]. 中国农业科学, 2021, 54(7): 1439-1454.
Han Z Y, Wu C Y, Xu Y Q, Huang F D, Xiong Y Q, Guan X Y, Zhou L J, Pan G, Cheng F M.Effects of high temperature on storage protein accumulation and anabolic metabolism of rice under different nitrogen application levels[J]., 2021, 54(7): 1439-1454. (in Chinese with English abstract)
[42] 郭银燕, 张云康, 蒋美明, 胡秉民, 陈昆荣. 浙江省早籼稻区试品种(系)碾磨品质研究[J].生物数学学报, 1996(1): 85-88.
Guo Y Y, Zhang Y K, Jiang M M, Hu B M, Chen K R.Study on milling quality of early indica rice varieties (lines) in Zhejiang Province[J]., 1996(1): 85-88.
[43] 周培南, 冯惟珠, 许乃霞, 张亚洁, 苏祖芳. 施氮量和移栽密度对水稻产量及稻米品质的影响[J]. 江苏农业研究, 2001(1): 27-31.
Zhou P N, Feng W Z, Xu N X, Zhang Y J, Su Z F.Effects of nitrogen application rate and transplanting density on rice yield and quality[J]., 2001(1): 27-31.
[44] 杨志根. 不同施肥量对水稻产量及经济性状的影响[J].上海农业科技, 2011(4): 94.
Yang Z G.Effects of different fertilizer application rates on rice yield and economic characters[J]., 2011(4): 94.
[45] Chen T T, Yang X Q, Fu W M, Li G Y, Feng B H, Fu G F, Tao L X. Strengthened assimilate transport improves yield and quality of super rice[J]., 2022, 12(4): 753.
[46] Li G Y, Chen T T, Feng B H, Peng S B, Tao L X, Fu G F. Respiration, rather than photosynthesis, determines rice yield loss under moderate high-temperature conditions[J]., 2021, 12: 1287-.
[47] Robaglia C, Thomas M, Meyer C. Sensing nutrient and energy status by SnRK1 and TOR kinases[J]., 2012, 15(3): 301-7.
[48] Rodriguez M, Parola R, Andreola S, Pereyra C, Martínez-Noël G. TOR and SnRK1 signaling pathways in plant response to abiotic stresses: Do they always act according to the "yin-yang" model[J]?, 2019, 288: 110220.
[49] 李志刚, 叶正钱, 杨肖娥, V.V.Virmani. 不同养分管理对杂交稻生育后期功能叶生理活性和籽粒灌浆的影响[J].浙江大学学报: 农业与生命科学版, 2003(3): 31-36.
LI Z G, YE Z Q, YANG X E, Virmani V V.Effects of different nutrient management on physiological activities of functional leaves and grain filling in late growth period of hybrid rice[J].:, 2003(3): 31-36.
Effect of Different Fertilizer Application Rates on Rice Yield and Quality Formation and Its Relationship with Energy Metabolism at High Temperature
WANG Wenting1,#, MA Jiaying1,#, LI Guangyan1,3, FU Weimeng1, LI Hubo1, LIN Jie1, CHEN Tingting1, FENG Baohua1, TAO Longxing1, FU Guanfu1, QIN Yebo2,*
(Yangzhou University, Yangzhou 225009, China; These authors contributed equally to this work; * Corresponding author, email: qyb.leaf@163.com)
【Objective】In recent years, frequent occurrence of extreme high temperature weather severely inhibited the formation of rice yield and quality. Optimizing plant nutritional status and improving energy metabolism can alleviate thermal damage. However, the effects of fertilizer application rates on grain energy metabolism and its relationship with yield, quality and thermal resistance at high temperature have not been reported, and elucidating the mechanism can lay an important theoretical basis for the research and development of rice heat-resistant and risk-resistant cultivation technology.【Method】In this study, a single croppinghybrid rice variety Zhongzheyou 8, which was widely planted in Zhejiang Province in recent years, was pot-cultivated in a climate chamber. Three fertilizer treatments were set: no fertilization (0-NPK), 1/2 normal fertilization (1/2-NPK) and normal fertilization (1-NPK). On the day of flowering, they were moved to the artificial climate chamber for high temperature treatment for 15d (for high temperature treatment, 36℃ in daytime and 28℃ at night; for the normal temperature treatment, 28℃ in daytime and 25℃ at night).【Result】The yield, seed-setting rate and 1000-grain weight of Zhongzheyou 8 followed a trend of increasing with the increaing fertilizer application at normal temperature or high temperature, with a shrinking decreasing amplitude at high temperature. However, head rice rate decreased with the increase of fertilizer application, and chalkiness degree showed an upward trend. The contents of soluble sugar, MDA, H2O2, ATPase and poly(ADP-ribose) polymerase (PARP) in the grains after high temperature treatment were higher than those in the normal temperature control, but contents of starch, non-structural carbohydrate, antioxidant enzyme and ATP were lower than those in the normal temperature control. Both at normal temperature and high temperature, the activities of non-structural carbohydrates, antioxidant enzymes, ATP and ATPase increased with rising fertilizer application level, while contents of MDA, H2O2and PARP decreased.At normal temperature, relative expression levels of energy receptor genesanddecreased with the increase of fertilizer application, whileshowed an increasing trend.Compared with normal temperature treatment, after high temperature treatment, the expression ofwas upregulated, whilewas downregulated with the increase of fertilizer amount.【Conclusion】Increased fertilizer application could effectively alleviate the negative effect of high temperature on seed setting rate and 1000-grain weight of rice during flowering, but appearance quality and processing quality tended to deteriorate. At normal temperature, the quality deterioration caused by increased fertilizer application may be related to the acceleration of grain filling, while the quality deterioration caused by high temperature is mainly due to the lack of energy, which is difficult to meet the demand of yield and quality formation.
rice; heat damage; fertilizer application amount; yield and quality; energy metabolism
10.16819/j.1001-7216.2023.220808
2022-08-25;
2022-12-05。
浙江省重点研发计划尖兵计划项目(2022C02014);浙江省基础公益研究计划资助项目(LY19C130006, LY20C130011);中国水稻研究所所级重点研发项目(CNRRI-2020-05);水稻生物学国家重点实验室开放课题(20210402)。
