水稻种子胚形态与胚乳组成对稻米留胚特性的影响
2023-05-18马兆惠石一涵程海涛宋文雯路连吉刘仁广吕文彦
马兆惠 石一涵 程海涛 宋文雯 路连吉 刘仁广 吕文彦*
水稻种子胚形态与胚乳组成对稻米留胚特性的影响
马兆惠1,#石一涵1,#程海涛1宋文雯1路连吉1刘仁广2吕文彦1,*
(1沈阳农业大学农学院,沈阳 110866;2本溪市农业综合发展服务中心, 辽宁 本溪 117022;*通信联系人,email:lwyzxr@syau.edu.cn)
【目的】稻米留胚能够提高稻米营养品质,但稻米留胚特性与胚性状、胚乳成分的关系等相关生物学机制并不明确。因此,有必要深入研究不同类型稻米品种留胚率差异及其影响因素。【方法】选取粳稻中软米型和普通型两类典型留胚材料及非留胚材料为试材,首先切片观察两类品种籽粒胚发育特点,继而对比表观直链淀粉含量(apparent amylose content, AAC)和离层果胶含量,最后以外源细胞分裂素(cytokinin, CTK)调控,比较胚性状、胚乳组成及留胚率差异。【结果】与非留胚材料相比,软米型和普通型留胚材料具有胚内幅和外幅较大,胚发育速度快,胚体积较大的特点;籽粒充实度好,粒形更宽、厚;同时离层果胶含量也较高。外施CTK后,软米型留胚材料中后期胚发育速度加快,普通型材料胚发育减缓;两类材料的AAC、果胶含量、胚比重及留胚率均提高。【结论】除胚性状对两类品种留胚率均有明显影响外,胚乳组成主要影响软米型材料留胚特性,而粒形是影响普通型材料留胚特性的重要因素。CTK则通过影响籽粒不同性状对不同类型材料留胚率进行调控。这为后续高留胚水稻品种的培育和栽培调控提供了理论依据和指导方向。
留胚率;胚发育;胚乳成分;粒形;细胞分裂素
随着生活水平的提高,人们越来越重视大米的营养品质。稻胚含有脂肪、氨基酸、维生素等丰富的营养元素[1],但在加工过程中大多因碾磨而随胚丢失,导致精米无法实现营养与食味兼备,而糙米口感欠佳。若想在保证稻米优良口感之余进一步提高其营养品质,留胚米毫无疑问成为了一个折衷的选择,留胚品种的研究成为了稻米研究的一个新方向。探索不同类型留胚品种的留胚机制,有助于进一步明确留胚品种高留胚率的特异性及其形成规律,对于选育留胚米具有重要意义。
留胚米通常指糙米经过碾磨加工之后,留胚率在80%以上或胚的质量占整体质量2%以上的稻米[2]。从表型来看,稻米留胚就是胚与胚乳粘连没有被完全破坏,因此,凡是胚体有残留,不论留有多少,均属于留胚米。稻米含胚与加工精度、品种、粒形和胚发育情况等因素有关。一般稻米的留胚率达80%以上时,长宽比小于1.45,即一般短粗形稻米利于留胚[3]。反田嘉博[4]研究发现,通常胚越长,胚的伸出(胚的外侧)大,而嵌入(胚的内侧)小,则胚越容易脱落。籽粒发育过程中,盾片外侧胚乳分化出一层特化细胞(即离层),胚通过离层与胚乳相联系[5],离层组织的粘连强度决定胚与胚乳的结合力[6]。猜测离层(即胚与胚乳间)存在某种黏性物质,而这类黏性物质含量的差异导致水稻留胚率的差异。水稻淀粉可能是胚与胚乳之间粘连物质之一,直链淀粉含量(AAC)较高则黏性较低[7],AAC较低的品种一般黏性较大[8]。植物细胞之间最初主要是由果胶类物质粘连在一起[9],而水稻的胚与胚乳之间是否也存在果胶类的黏性物质?这类报道在谷类研究中尚不多见,值得进一步探讨。已经证实激素可通过调节纤维素酶和果胶酶活性来控制脱落,细胞分裂系(CTK)含量的变化不仅会引起果胶类物质含量和离层特性的改变,从而引发胚脱落[10],而且对胚器官细胞发育、胚乳淀粉积累及籽粒发育也有一定影响[11-12]。因此,细胞分裂素是否也对留胚特性有影响?课题组前期研究发现,灌浆好、胚乳充实度高是提高留胚率的关键因素,据此推测,CTK可能通过对胚乳发育的影响从而间接影响胚的着生、发育和脱落,但水稻上的相关研究还很少。
因此,本研究选择留胚率有显著差异的典型材料,首先通过观察胚及籽粒发育动态,得到两类材料胚形态、体积、结构差异,分析留胚材料的胚发育特性,继而测定籽粒AAC和离层果胶含量,研究胚与胚乳粘连程度和特点,最后以外源激素CTK调控,比较胚性状、理化指标及留胚率差异,以期初步探明水稻高留胚率的生物学特征。本研究旨在揭示影响水稻留胚率的主要因素,并根据不同类型水稻品种的不同留胚影响因素,确定提高稻米留胚率的栽培调控方法,以期应用于水稻高留胚育种和栽培工作。
1 材料与方法
1.1 试验材料
选择2个留胚和2个非留胚粳稻材料为试材,分别为彦粳软玉1号(软米留胚型,8月4日齐穗)、1419-8(软米非留胚型,为彦粳软玉1号后代,8月3日齐穗)、2045-5(普通留胚型,8月5日齐穗)和19-3(普通非留胚型,8月4日齐穗)。
1.2 田间栽培管理
试验于沈阳农业大学试验水田(41°83'N、123°57'E)展开,试验地基本情况为碱解氮65.70 mg/kg,有效磷13.59 mg/kg,速效钾120.40 mg/kg,有机质25.63 g/kg,土壤pH 5.53,电导率(土壤含盐量与水土比为5:1) 274.70mS/m。2020年4月中旬播种,5月下旬插秧。田间采用随机区组排列,3次重复,每个材料3 m行长,6行区,行株距为30 cm×13.3 cm,每穴2株苗。试验田基施硫酸钾型复合肥50 kg/hm2。
在孕穗期时,叶片喷施细胞分裂素(6-苄基氨基嘌呤)。细胞分裂素用95%的乙醇完全溶解后,配制成400 mg/L母液,施用时再稀释成各梯度浓度,对照为纯净水加入等体积的95%乙醇。处理时间选择在16∶00-18∶00,每个处理喷施溶液1 L。经前期预备试验对比,最终采用CK(空白对照)、H(20 mg/L)两个处理。激素处理后,标记同一天开花、长势均匀一致的籽粒,根据稻胚发育进程,分别于标记后9、12、30 d取样,每次取穗中上部强势粒40粒,放入装有FAA固定液(90 mL 70%乙醇+5 mL 38%甲醛+5 mL冰醋酸+5 mL甘油)的离心管后置于4℃冰箱内贮存,用于测定有关性状。
1.3 性状测定
1.3.1 千粒重
稻谷自然干燥后,采用稻谷计数板量出3份稻谷,每份200粒,称量后取平均值,将结果乘以5,获得水稻样品的千粒重。
1.3.2 粒形
用万深SC-E型种子大米外观品质检测仪测定长宽比、粒长和粒宽。随机选取10~15粒籽粒,用游标卡尺测定其厚度,求平均值。
1.3.3 胚比重
将100粒糙米称重(1),将胚部用镊子剥离,称量其重量(2),其胚比重=(2/1)×100%,重复3次,取平均值。
1.3.4 留胚率
稻谷利用韩国KMT-400B6脱壳机脱壳,Keet精米机碾精(碾精度在90%±1%)得到整精米,然后按照四分法取样:10 g整米试样排成一粒米厚度的正方形,用分样板在样品上划两条对角线,分成4个三角形,从每个三角形中,各随机数出25粒,100粒计为一个试样,然后取3份平均试样,将试样摊在白色洁净的搪瓷盘里,在明亮处(应采用背向光)逐粒鉴别,并按水稻留胚级别[13]分类,半胚及半胚以上米粒数之和计为留胚米,即该试样留胚率。
1.3.5 米粉的制备
整糙米采用FSⅡ旋风式粉碎磨制成米粉,分成两等份,一份过80目筛,用于总果胶含量的测定;另一份过100目筛,用于表观直链淀粉含量的测定。
1.3.6 直链淀粉含量的测定
参照《ISO 6647-2:2007直链淀粉含量测定方法》,其中标准样品来自于中国水稻研究所(CRRI)。
1.3.7 总果胶含量的测定
总果胶含量测定参照张雪丹[14]的方法。
1.3.8 胚形态的观察
胚形态观察采用石蜡切片和徒手切片相结合的方法,石蜡切片制作参照李娜[15]的方法;徒手切片方法具体如下:将糙米放在蒸馏水中浸泡15 min,取出后放在吸水纸上吸干水分。将整个籽粒从中部截断,然后将含有胚的半个籽粒立在桌面上,从籽粒上端中间与胚横截面水平角度用双面刀片切开,取两侧第一片,用于显微镜观察,测量胚部各性状。每个材料做25~30片,除去异常值(|测量值-平均值|/平均值>15%),选择形态、大小类似的作为最终计算数值。
1.3 数据分析
运用DPS 15.10数据处理系统进行多重比较和相关分析,用Sigmaplot作图软件绘图。
2 结果与分析
2.1 不同类型材料胚性状差异比较
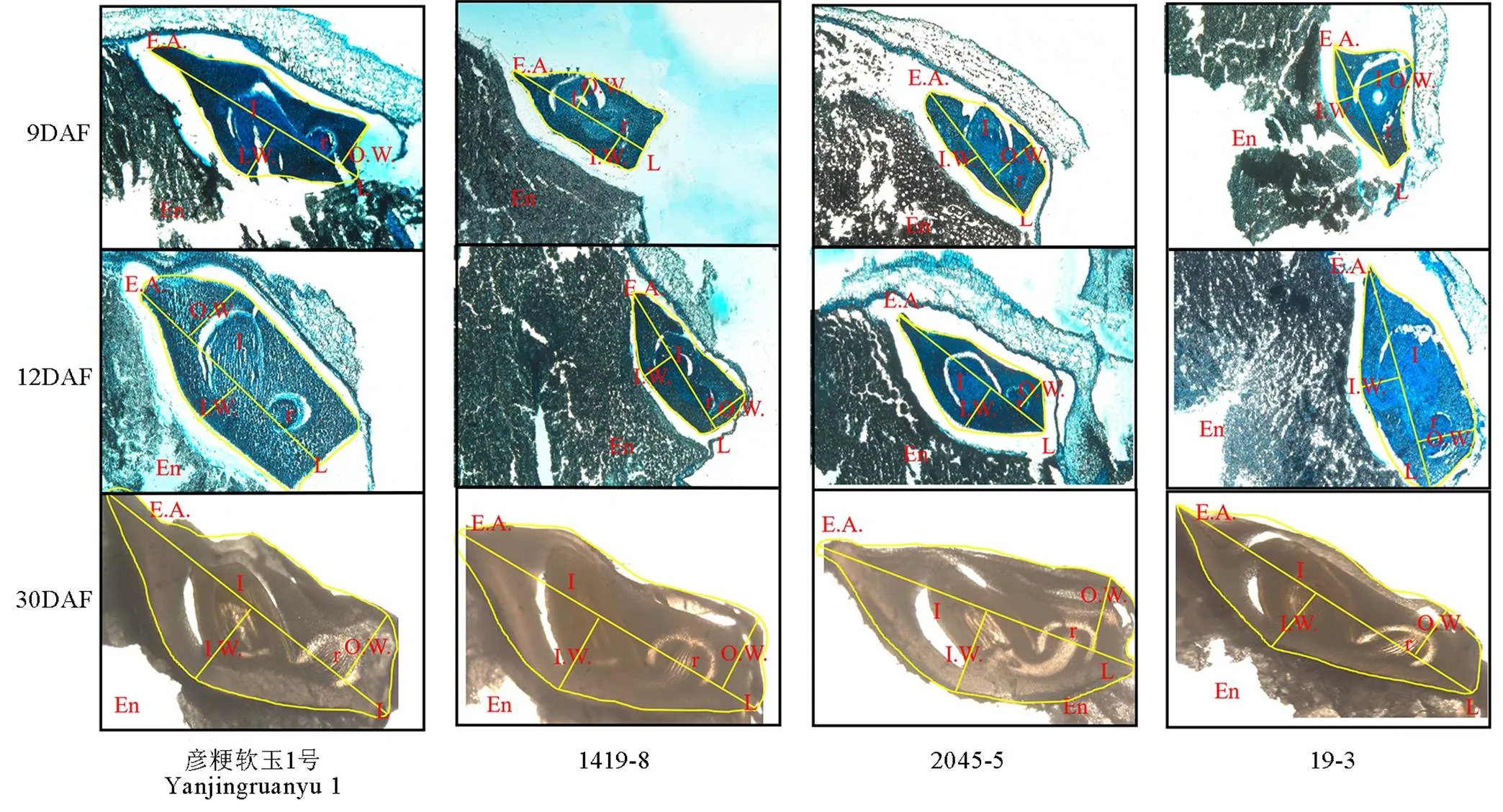
切片观察软米型和普通型两类材料花后9 d、12 d和30 d胚部形态特征(图1,表1),结果表明,在胚发育前期与中期,软米型留胚材料彦粳软玉1号的内幅、胚长、外幅、胚面积及胚发育速度均显著大于非留胚材料1419-8,并在整个时期保持了较大的胚体积(图1)。普通型留胚材料2045-5在花后9 d胚部各指标显著小于非留胚材料19-3,但其在整个生育期的胚发育速度较19-3快,因此在花后12 d至成熟期,2045-5胚的内幅、外幅和面积显著大于19-3,表现出大胚的特点。
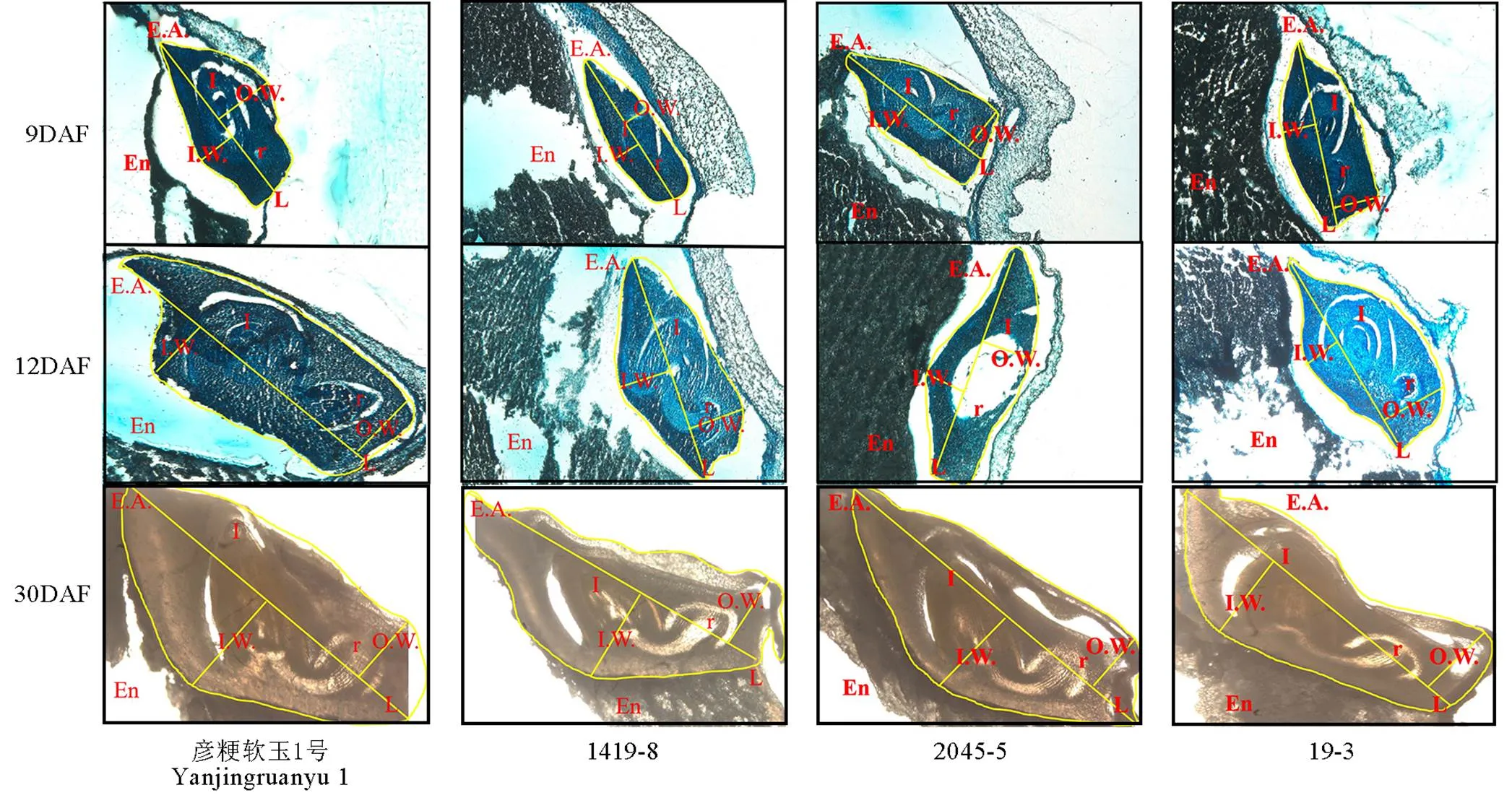
EA-胚面积;En-胚乳;L-胚长;I-胚芽;IW-内幅;OW-外幅;R-胚根。DAF-花后天数。
Fig. 1. Embryo-specific morphologies of different materials at different developmental stages.
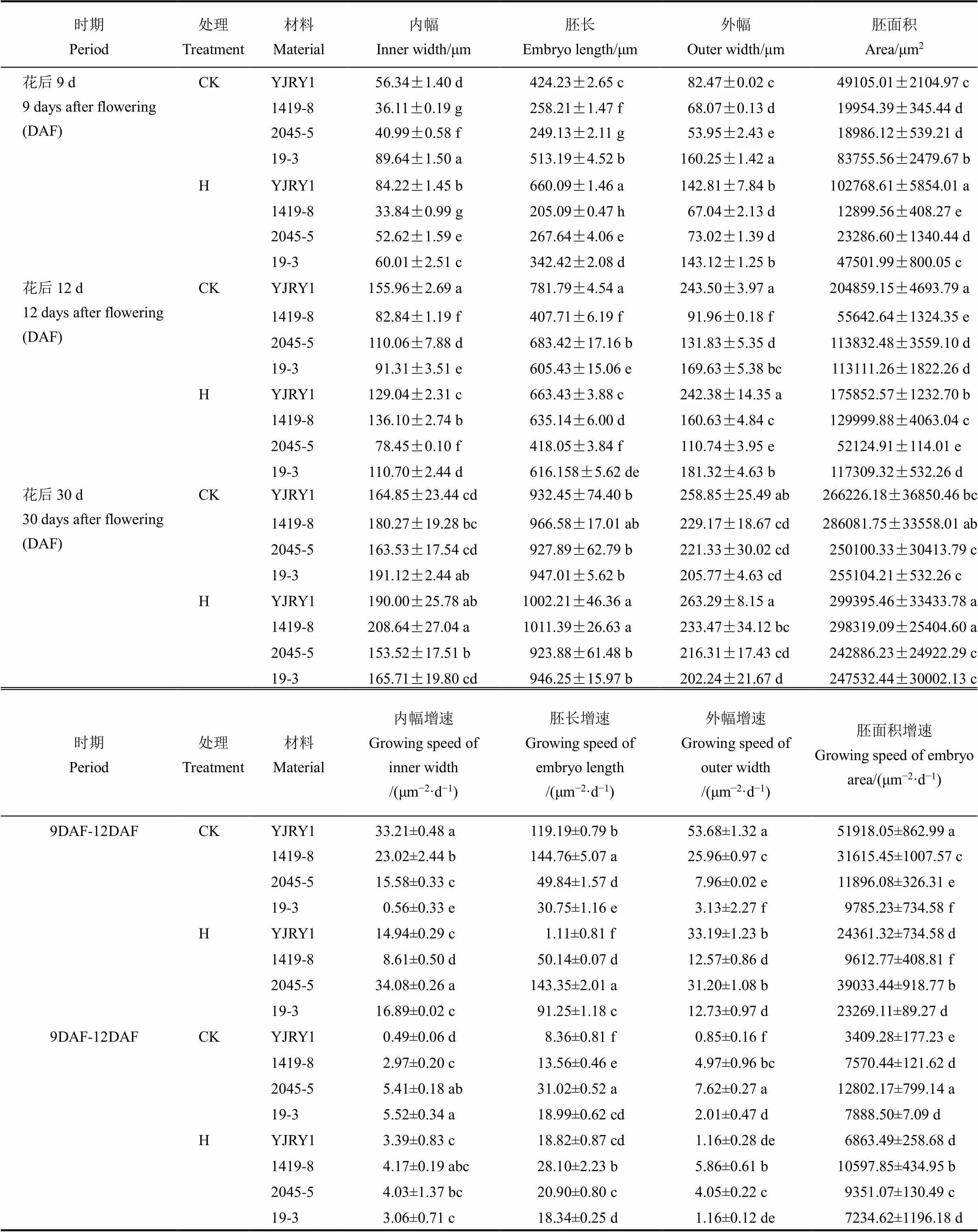
表1 施用细胞分裂素后不同时期留胚和非留胚材料胚部性状及发育速率对比
不同小写字母表示在0.05水平上差异显著。CK-对照;H-施用细胞分裂系。
Values flanked by various lowercase letters are significantly different at the level of 0.05. CK, Control; H, Cytokinin application.
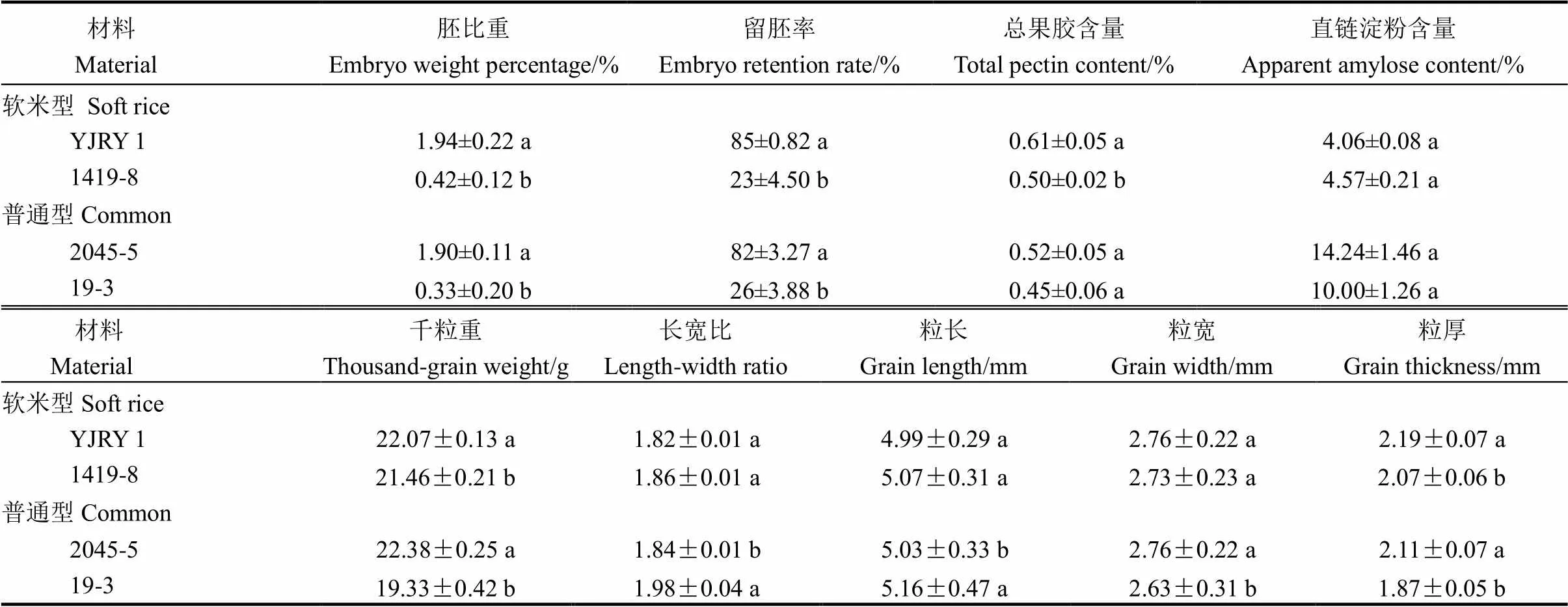
表2 不同类型材料留胚特性及籽粒性状比较
YJRY 1-彦粳软玉1号;不同字母表示同一类型品种间在5%水平上差异显著。下同。
YJRY 1, Yanjingruanyu 1. Different letters indicate significance at the 5% level. The same below.
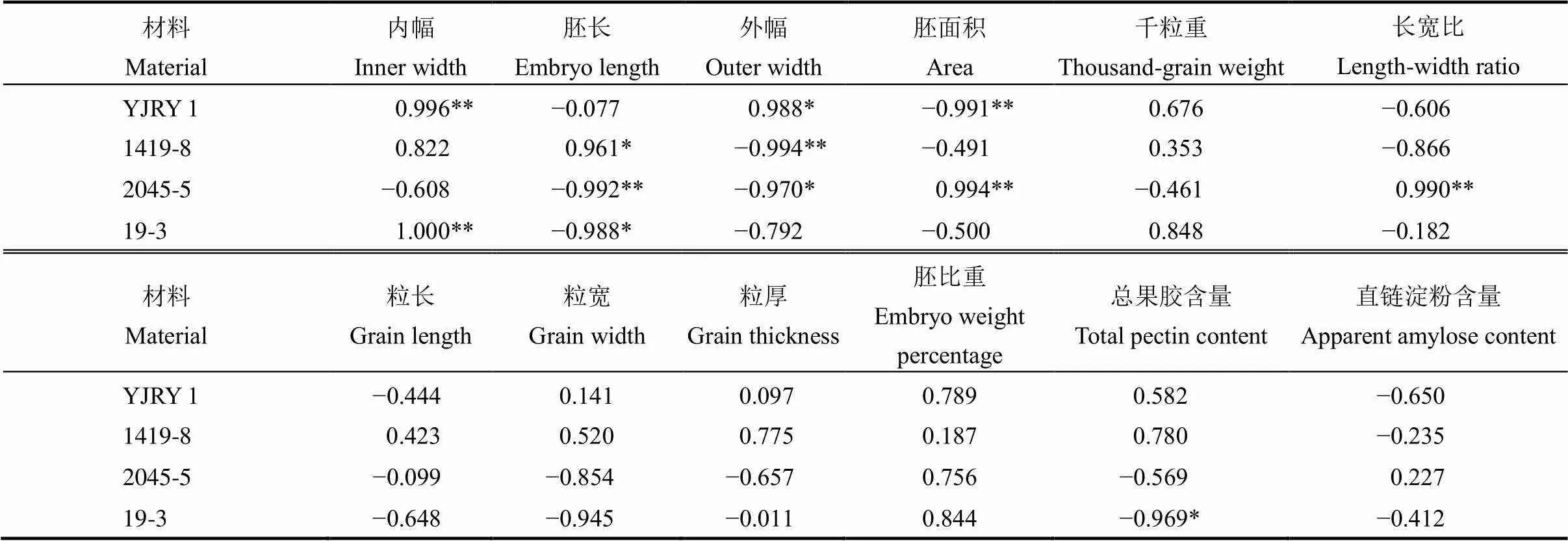
表3 不同类型材料籽粒性状与留胚率的关系
=5。**表示该性状与留胚率达极显著相关;*表示显著相关。
=5. **,<0.01; *,<0.05.
2.2 不同类型材料离层物质含量及留胚特性比较
两类留胚材料的胚比重和留胚率均显著高于两类非留胚材料(表2),其中软米型留胚材料的胚比重和留胚率最大;在离层物质含量上,留胚材料的总果胶含量均大于同类型的非留胚材料,其中软米型材料间的果胶含量差异达显著水平。从直链淀粉含量看,材料间差异不显著,但软米型留胚材料的直链淀粉含量较非留胚材料低,因此胚乳黏性较高,这可能是其高留胚率的原因之一。
2.3 不同类型材料粒形及千粒重差异比较
研究结果表明,留胚材料与非留胚材料之间千粒重差异均显著(表2),说明无论是软米型还是普通型,留胚材料的籽粒充实度高于非留胚材料。从粒形看,留胚材料的籽粒更宽、厚,但粒长和长宽比值较非留胚材料低,这种差异在普通型材料间达到显著,说明留胚材料更趋近于短圆粒形。
2.4 不同类型材料籽粒性状与留胚率的关系
籽粒性状中,胚形态与留胚率关系最密切(表3)。软米型留胚材料的内幅、外幅与留胚率达到显著正相关;普通型留胚材料的胚面积与留胚率极显著正相关。此外,增加胚比重对提高留胚率起促进作用。从粒形看,长宽比对普通型留胚材料的留胚率影响更明显;粒厚增加能提高软米型材料留胚率。从离层成分看,软米型材料果胶含量与留胚率呈正相关,直链淀粉含量与留胚率呈负相关,而普通型材料果胶含量与留胚率呈负相关,说明离层成分对留胚率的影响因材料类型而异。
EA-胚面积;En-胚乳;L-胚长;I-胚芽;IW-内幅;OW-外幅;R-胚根。
Fig. 2. Comparison of embryo morphology of different types of embryo-remaining material after cytokinin application.
2.5 外施CTK对籽粒各性状及留胚特性的影响
与对照相比,外施CTK后使发育前期(花后9 d)留胚材料的胚各性状值均有所增加(图2,表1),胚发育速度加快(花后9~12 d)。在发育中后期(12~30 d),CTK抑制普通型材料的胚发育速度,加速软米型材料胚发育,因此最终成熟时,软米材料胚各性状值较高,普通型材料胚性状值降低。此外,外施CTK增加了胚比重和留胚率(表4),其中两类非留胚材料增幅较大,增幅达36.1%和46.5%。果胶含量因外施CTK而增加,其中普通型留胚材料的增幅明显(30.9%)。外施CTK使各材料的千粒重降低,软米型材料粒厚减小,普通型材料粒厚增加,留胚材料粒宽下降而长宽比增加,非留胚材料粒宽增加而长宽比下降。
2.6 CTK施用量与留胚特性的相关性分析
外施CTK对不同类型材料的胚发育特性、籽粒性状及胚乳成分含量有不同程度的影响(表4)。CTK施用量与留胚材料各胚性状正相关,但CTK施用量与非留胚材料的胚性状负相关。从粒形看,留胚材料的粒长、粒宽、粒厚均与CTK施用量负相关,其中粒长、粒厚受CTK负影响较大。离层成分中,果胶含量和直链淀粉含量与CTK用量均呈显著或极显著正相关。留胚特性中,留胚率和胚比重均与CTK施用量表现为正相关,其中留胚材料与CTK施用量的相关更显著。
3 讨论
3.1 不同类型留胚材料胚性状及籽粒性状差异比较
前人对胚的发育进程研究虽已基本明确[16-19],但不同类型、不同留胚率品种间胚发育是否存在差异?本研究结果表明,软米型留胚材料在早期胚发育速度快,后期发育速度慢,且胚体积始终大于非留胚材料,这不仅符合早期胚迅速生长发育的特征[20],更证明早期是软米型材料胚发育及胚体积形成的关键时期。而普通型留胚材料在整个生育期均表现出较非留胚材料快的发育速度,因此最终成熟期时,二者胚大小近乎一致,说明普通型材料的胚建成贯穿整个胚发育时期,这验证了前人的结论[17-18]。同时,还发现留胚米的胚质地较为紧密,非留胚米胚极易碎裂,难以取得完整胚。因此,进一步研究可以从留胚米胚部质地方向着手。前期研究发现,留胚率高的籽粒具有较低的AAC[21],说明低AAC有助于留胚,而本研究中只有软米型材料符合这一特征,因此AAC作为判断留胚率高低的指标应因材料类型而异。Michal等[22]通过红醋栗实验证明了果胶含量与黏度特性之间存在关系,但关于果胶与黏性的报道主要集中在浆果类,在谷物类研究中尚属空白。我们的研究发现,留胚材料的总果胶含量较大,故此推测总果胶含量差异使胚与胚乳粘连程度不同,是影响留胚的原因之一,此结果与张雪丹[14]的研究结果相似。
表4 不同类型材料外施细胞分裂素后籽粒性状及理化性质的变化
不同字母表示在5%水平上差异显著。
Different letters indicate significance at the 5% level.
高留胚品种充实度好,千粒重大,籽粒宽厚,这与前期研究结果相同[23],且这种特点在普通型留胚品种表现明显,说明粒形和充实度是影响普通型品种间留胚率的重要因素。此外,通过分级碾精加工发现(图3),随碾磨时间增加,留胚材料的留胚类型由全胚(A)和平胚(B),转变成少量半胚(C)和大部分的残胚(D);而非留胚材料的留胚类型由半胚(C)转变成残胚(D)和无胚(E)。这种留胚类型的差异,证明留胚材料的胚与胚乳粘连紧密且抗碾磨。而本研究中发现该类材料在胚发育、籽粒性状及胚乳成分方面与非留胚材料相比,存在一定特点和差异,从而具有高留胚率的特性。
3.2 影响不同类型品种留胚率的因素分析
影响稻米留胚的因素多样[24],留胚首先与胚形态结构有关,当胚嵌入米粒较浅且自身体积大时,则受碾磨能力弱,易脱落[4]。本研究发现留胚材料具有米胚外形钝圆,且胚整体向粒内伸展长度/向外伸展长度比例较大的特点,因此更抗碾磨。从相关分析看,软米型留胚材料的胚嵌入方式是影响留胚率的主要因素,而普通型留胚材料的留胚率主要由胚面积决定,说明不同类型留胚材料的留胚率由不同的胚形态性状影响。胚乳的主要组成成分是淀粉,其中AAC是影响米粉黏性的主要因子[25-26],通常AAC低更有利于胚乳与胚相粘连。本研究中,与非留胚材料相比,软米型留胚材料的AAC低且其与留胚率负相关,进一步证明较低的AAC是保证软米型材料高留胚率的因素之一。果胶作为植物细胞之间最初的粘合物质,其浓度越大则黏性越强[27]。本研究也发现总果胶含量高是导致软米型材料具有高留胚率的重要因素,但普通型材料总果胶含量高不利于留胚率,是否因果胶种类及占比不同而造成差异,还有待进一步研究。
水稻粒形一般指的是粒长、粒宽、粒厚以及长宽比[28],稻米加工时,粒形影响籽粒碾磨的接触面积,因此是留胚的一个决定因素。有研究表明,水稻留胚率与粒长、长宽比显著负相关,与粒厚、粒宽正相关[21],也有研究得出留胚率与粒长、宽、长宽比不显著负相关的结果[29]。课题组前期研究发现,对于一般材料而言,粒形越长的稻米,耐磨性越差[23],长宽比增加,留胚率则下降[30]。而本研究中,普通型留胚材料的长宽比与留胚率呈极显著正相关,说明对于较籼稻粒形更短圆的粳稻品种而言,适当提高长宽比可以提高留胚率。此外,粒重对胚比重有显著影响[31],本研究中两类留胚材料的胚比重与留胚率均正相关,这说明,粒重通过影响胚比重进而影响留胚率,即籽粒充实度是影响留胚率的基本因素之一。
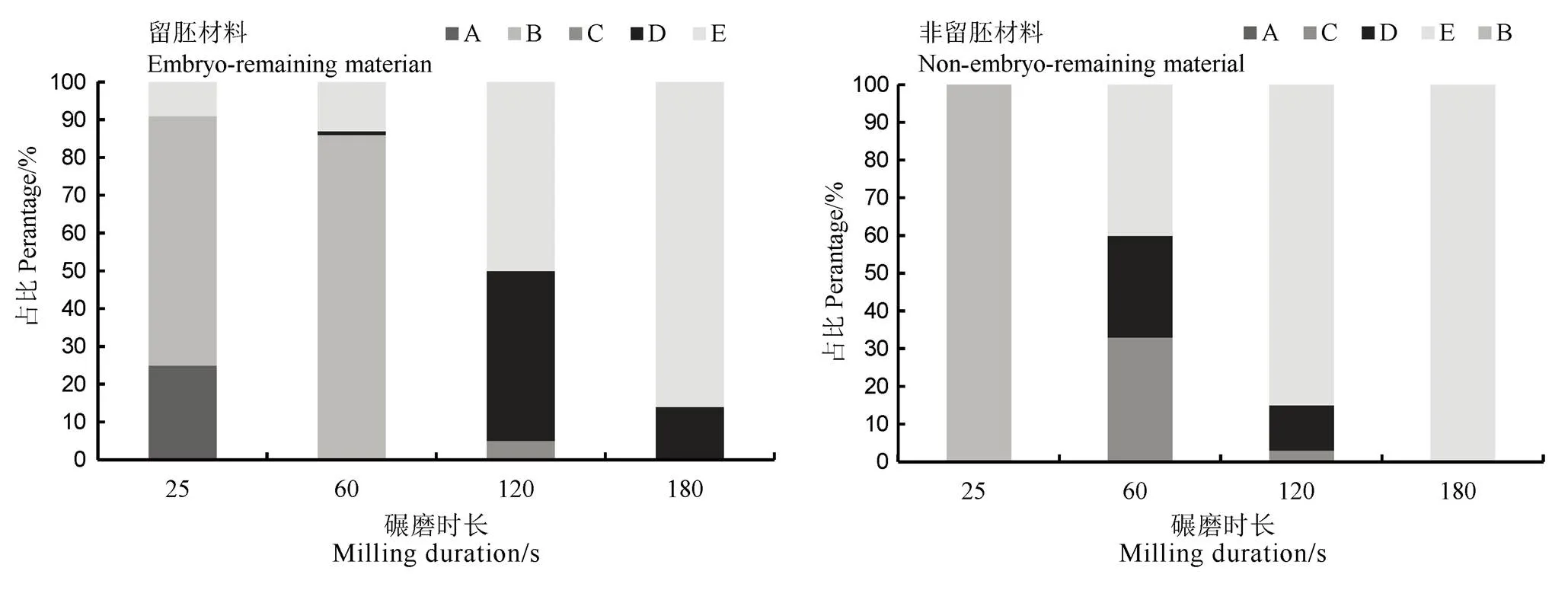
A-全胚, 糙米研磨后,胚几乎没有磨损;B-平胚,糙米碾碎后,剩下的胚与米尖相切;C-半胚, 糙米磨碎后,将一半胚磨掉;D-残胚, 糙米研磨后,只剩下部分胚;E-无胚, 糙米研磨后,所有的胚都被研磨掉,没有任何残留物。
Fig. 3. Changes of embryo remaining types of embryo-remaining rice and non-embryo-remaining rice under different milling time.
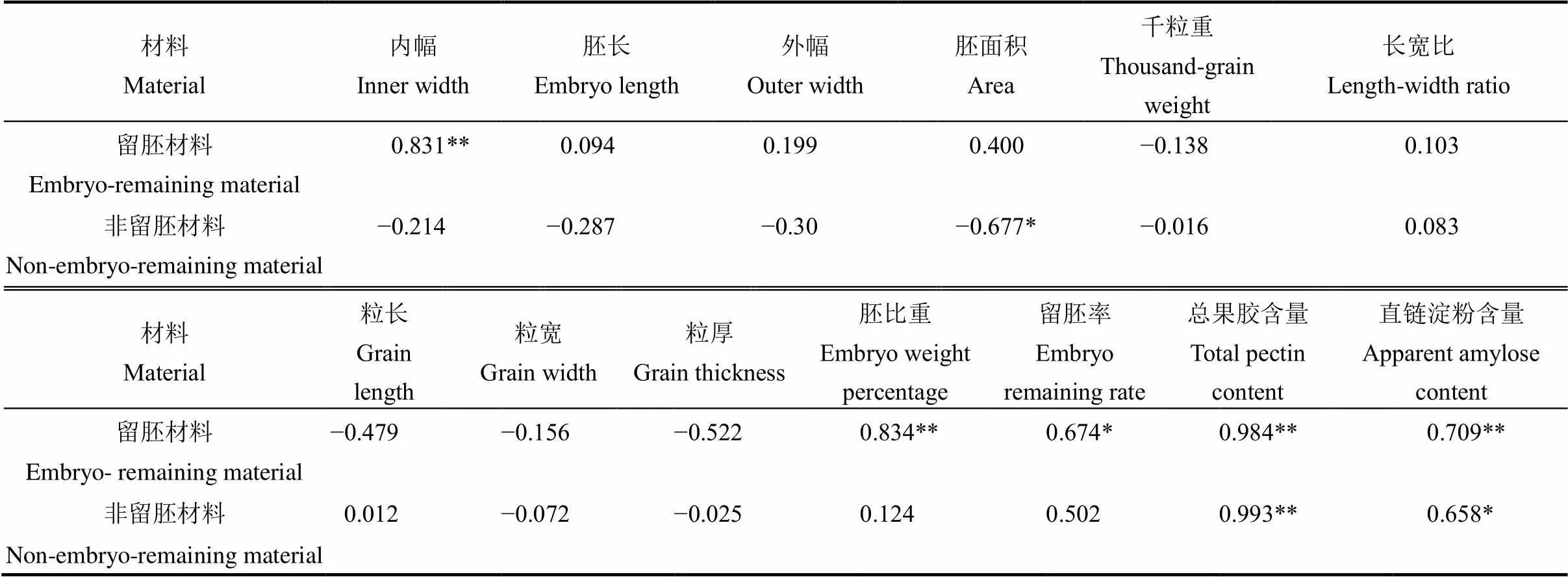
表5 CTK施用量与不同类型材料籽粒性状及留胚率的相关性
**表示极显著相关;*表示显著相关。
**,<0.01;*,<0.05.
3.3 外施CTK对不同类型材料籽粒性状及留胚特性的影响
细胞分裂素(CTK)对籽粒发育有重要影响[32],可使胚增大[33]。本研究也发现,与对照相比,CTK处理加速了软米型材料中后期的胚发育,使其形成大胚;但普通型材料在CTK处理下整个生育期胚发育速率下降,胚值减小,说明CTK处理可能抑制普通型材料胚细胞的分生组织[34]。即外源施用CTK不仅对胚发育的影响时期不同,影响结果也因材料类型而异。CTK诱导的水稻基因Cga1下调表达时,籽粒变得细长[35-36],即当CTK浓度较低时,籽粒发育会趋于细长形。本研究中,与对照相比,CTK处理提高了留胚材料的长宽比,而普通型留胚米的长宽比与留胚率呈极显著正相关,因此,普通型材料可通过CTK调控粒形进而提高留胚率。
前人研究已经证实,内源激素能通过调节纤维素酶和果胶酶活性引起果胶类物质含量和离层黏性的改变,进而影响留胚[37-39]。对于留胚米的高留胚率机理,有研究推测其胚和胚乳间具有较高含量的果胶,导致离层粘连特性变化。本研究发现外施CTK对果胶含量提高有明显的促进作用,软米类材料可通过外施CTK,显著提高果胶含量,进而提高留胚率。此外,CTK有调控同化物积累和胚乳发育[40],促进淀粉积累[41]等作用,但其对水稻留胚特性的影响方面尚未见报道。本研究中,两类材料的AAC与CTK量显著正相关,证明CTK在水稻中也具有促进淀粉积累的作用,并通过调控AAC含量影响不同类型材料的留胚率。
4 结论
本研究结果表明,留胚材料总体上胚发育速度快,胚体积较大,胚质地紧密。软米型材料胚嵌入胚乳部位较深,并具有较高果胶含量。普通型材料的胚面积大,粒形宽厚。通过外源CTK调控发现,施用CTK可以促进软米型材料胚内幅、外幅发育,并提高离层黏性物质含量;而对于普通型材料,CTK主要增加胚比重和粒长。因此,CTK对不同类型材料留胚率的调控是通过影响水稻籽粒不同性状实现,可根据材料特性进行留胚率调控。
[1] 朱一帆, 冯亚斌, 林轩, 周美圆, 王中华. 胚芽米营养成分研究[J]. 科技通报, 2013,29(9): 51-54.
Zhu Y F, Feng Y B, Lin X, Zhou M Y, Wang Z H. Nutritional components analysis of germ-remaining rice[J].,2013,29(9): 51-54. (in Chinese with English abstract)
[2] 宋幼良, 吴殿星. 高留胚糯稻突变体原胚糯的选育与营养成分分析[J]. 核农学报, 2017, 31(9): 1672-1677.
Song Y L, Wu, D X. Breeding and nutrient content analysis of glutinous rice mutant Yuanpeinuo with high remained germ[J].,2017,31(9): 1672-1677. (in Chinese with English abstract)
[3] Liang J, Li Z, Tsuji K. Milling characteristics and distribution of phytic acid and zinc in long-, medium-and short-grain rice[J]., 2008, 48(1): 83-91.
[4] Tanda Y. Research on embryo detachment by polishing early cultivated rice[J].1962, 30: 9-13. (in Japanese)
[5] Yang W, Gao M, Yin X, Liu J, Xu Y, Zeng L, Li Q, Zhang S, Wang J, Zhang X, He Z. Control of rice embryo development, shoot apical meristem maintenance, and grain yield by a novel cytochrome P450[J]., 2013, 6(6): 1945-1960.
[6] Wan Z, Xia J, Kang D. The Research progress and prospect analysis of germ-remaining rice whitener[J].:,2008, 15(2): 84-87.
[7] Kong X, Zhu P, Sui Z, Bao J. Physicochemical properties of starches from diverse rice cultivars varying in apparent amylose content and gelatinization temperature combinations[J]., 2015,172: 433-440.
[8] Zhou L, Sheng W, Jun W, Wang C, Liu Q, Deng Q. Differential expressions among five Waxy alleles and their effects on the eating and cooking qualities in specialty rice cultivars[J]., 2015, 14(6): 1153-1162.
[9] Müller-Maatsch J, Bencivenni M, Caligiani A, Tedeschi T, Bruggeman G, Bosch M, Petrusan J, Droogenbroeck BV, Elstf K, Sforza S. Pectin content and composition from different food waste streams[J]., 2016,201: 37-45.
[10] Yuan R C, Burns J K. Temperature factor affecting the abscission, response of mature fruit and leaves to CMN-Pyrazole and ethephon in Hamlin oranges[J].,2004,129(3): 287-293.
[11] Jameson P E, Song J. Cytokinin: A key driver of seed yield[J]., 2015, 67: 593-606.
[12] Han Y, Yang H, Jiao Y. Regulation of inflorescence architecture by cytokinins[J]., 2014,5: 669.
[13] 李爱华. 米胚的营养价值和胚芽米的加工技术[J]. 粮食与饲料工业, 1997(5): 9-12.
Li A H. The nutritive value of rice germ and the processing technology of germ-remaining rice[J]., 1997(5): 9-12. (in Chinese with English abstract)
[14] 张雪丹. 苹果果胶制备工艺及研究进展[J]. 落叶果树, 2009, 41(2): 22-25.
Zhang X D. Preparation technology and research progress of apple pectin[J]., 2009, 41(2): 22-25. (in Chinese with English abstract)
[15] 李娜.基因对小穗花器官发育的影响研究[D]. 沈阳: 沈阳农业大学, 2015.
Li N. Functional analysis ofgene in spikelet development[D]. Shenyang: Shenyang Agricultural University, 2015. (in Chinese with English abstract)
[16] 杨弘远. 水稻生殖生物学[M]. 杭州: 浙江大学出版社, 2005: 102-125.
Yang H Y. Rice Reproductive Biology[M]. Hangzhou: Zhejiang University Press, 2005: 102-125 (in Chinese).
[17] 王敏. 关于水稻颖界发育的研究[D]. 扬州: 扬州大学, 2011.
Wang M. Caryopsis development of rice[D]. Yangzhou: Yangzhou University, 2011. (in Chinese with English abstract)
[18] Itoh J I, Nonomura K I, Ikeda K. Rice plant development: From zygote to spikelet[J]., 2005,46(1): 23-47.
[19] Xu H, Zhang W, Gao Y, Zhao Y, Guo L, Wang J. Proteomic analysis of embryo development in rice () [J]., 2012, 235(4): 687-701.
[20] 章清杞, 陈健勇, 黄荣华, 张书标. 巨胚稻胚发育的解剖学观察[J]. 核农学报, 2008(2): 122-126.
Zhang Q Q, Chen J Y, Huang R H, Zhang S B. Anatomical observations on the developmental process of embryo in giant embryo rice[J]., 2008(2): 122-126(in Chinese with English abstract)
[21] 陈恒雪, 石一涵, 吕文彦, 马兆惠, 程海涛, 陈云. 稻米留胚率测定方法筛选及留胚米相关特性研究[J]. 沈阳农业大学学报, 2018, 49(3): 337-341.
Chen H X, Shi Y H, LW Y, Ma Z H, Cheng H T, Chen Y. Determination methods of rice plumule ratio and related characteristics of germ-remained rice[J]., 2018, 49(3): 337-341.
[22] Michal P, Joanna K, Marcin L, Magdalena W, Mateusz K, Jolanta J, Anna P. Red currant pectin: The physicochemical characteristic of pectin solutions in dilute and semi dilute regimes[J]., 2021, 113: 106420.
[23] Ma Z H, Chen H X, LW Y, Cheng H T, Chen Y, Wang Y B. Comparison of the chemical and textural properties of germ-remaining soft rice grains from different spikelet positions[J]., 2019, 96: 1137-1147.
[24] 熊海铮, 张宁, 孙健, 王寅, 舒小丽, 吴殿星. 水稻留胚米的营养价值、加工技术及产品开发研究进展[J]. 核农学报, 2012, 26(7): 1031-1036.
Xiong H Z, Zhang N, Sun J, Wang Y, Shu X L, Wu D X. Research progress on nutritional value, processing technology and product exploration of embryo-retaining milled rice[J]., 2012, 26(7):1031-1036. (in Chinese with English abstract)
[25] Ahmed N, Tetlow I, Nawaz S, Iqbal A, Mubin M, Rehman M, Butt A, Lightfootc D, Maekawa M. Effect of high temperature on grain filling period, yield, amylase content and activity of starch biosynthesis enzymes in endosperm of basmati rice[J]., 2015, 95: 2237-2243.
[26] Teng B, Zeng R, Wang Y, Liu Z, Zhang Z, Zhu H, Ding X, Li W, Zhang G. Detection of allelic variation at thelocus with single segment substitution lines in rice (L) [J]., 2012, 30 (1):583-595.
[27] Xing F, Owen G J, Bradley L R, Osvaldo H C. Soluble pectin acts as a particle stabilizer of tomato suspensions: The impact on tomato products rheological characterization[J]., 2021, 139(3): 110508.
[28] 周海庆. 水稻粒形及千粒重的遗传研究[D]. 吉林省延边市: 延边大学, 2014.
Zhou H Q. Genetic studies on the rice grain shape and 1000-grain weight[D]. Yanbian: Yanbian University, 2014. (in Chinese with English abstract)
[29] 赵飞. 粳稻稻米品质及其与产量关系的遗传分析[D]. 沈阳: 沈阳农业大学. 2014.
Zhao F. Genetic analysis on relationship between quality and yield of Japanese rice[D]. Shenyang: Shenyang Agricultural University, 2014.(in Chinese with English abstract)
[30] 王少元, 孟庆虹, 严松, 张志宏, 高扬, 袁超, 卢淑雯, 河野元信. 北方粳稻留胚米碾磨工艺的研究[J]. 中国稻米, 2014, 20(5): 31-35.
Wang S Y, Meng Q H, Yan S, Zhang Z H, Gao Y, Yuan C, Lu S W, Motonibu K. Research on the milling processing with germ- remained japonica rice of northern China[J]., 2014, 20(5): 31-35. (in Chinese with English abstract)
[31] 张标金, 魏益华, 张祥喜, 罗林广. 育成巨胚稻与其亲本的糙米粒型性状和矿物质含量的比较分析[J]. 中国农学通报, 2014, 30(6): 182-185.
Zhang BJ, Wei YH, Zhang XL, Luo LG. Comparative analysis on grain traits and mineral elements of brown rice from bred giant embryo rice and its parents[J]., 2014, 30(6): 182-185.
[32] 李志康, 严冬, 薛张逸, 顾逸彪, 李思嘉, 刘立军, 张耗, 王志琴,杨建昌, 顾骏飞.细胞分裂素对植物生长发育的调控机理研究进展及其在水稻生产中的应用探讨[J]. 中国水稻科学, 2018, 32(4): 311-324.
Li Z K, Yan D, Xue Z Y, Gu Y B, Li S J, Liu L J, Zhang H, Wang Z Q, Yang J C, Gu J F. Regulations of plant growth and development by cytokinins and their applications in rice production[J]., 2018, 32(4): 311-324. (in Chinese with English abstract)
[33] Werner T, Schmülling T. Cytokinin action in plant development[J]., 2009, 12(5): 527-538.
[34] 孙贝贝. 生长素和细胞分裂素的内源合成控制根从头发生的机制研究[D]. 上海: 上海师范大学, 2017.
Sun B B. Mechanism of specific function of plant hormones auxin and cytokinin during adventitious root regeneration[D]. Shanghai: Shanghai Normal University. 2017. (in Chinese with English abstract)
[35] Hudson D, Guevara D R, Hand A J. Rice cytokine in GATA transcription Factor1 regulates chloroplast development and plant architecture[J]., 2013, 162(1): 132-144.
[36] Li J, Nie X, Tan J L. Integration of epigenetic and genetic controls of seed size by cytokine in[J]., 2013, 110 (38): 15479-15484.
[37] Ashikari M, Wu J, Yano M. Rice gibberellin-in-sensitive dwarf mutant geneencodes the alpha-subunit of GTP-binding protein[J]., 1999, 96(18): 10284-10289.
[38] Fujisawa Y, Kato T, Ohki S. Suppression of the hetero trimetric G protein causes abnormal morphology, including dwarfism, in rice[J]., 1999, 96(13): 7575-7580.
[39] Nayar S, Sharma R, Tyagi A, Kapoor S. Functional delineation of rice MADS29 reveals its role in embryo and endosperm development by affecting hormone homeostasis[J]., 2013, 64: 4239-4253.
[40] Zhao Y, Hu Y, Dai M, Huang L, Zhou D. The WUSCHEL-Related homeobox geneis required to activate Shoot-Borne crown root development in rice[J]., 2009, 21(3): 736-748.
[41] 朱艳梅, 罗兴录, 樊吴静.木薯内源细胞分裂素含量对块根淀粉积累的影响[J]. 南方农业学报,2016, 47(8): 1279-1284.
Zhu Y M, Luo X L, Fan W J.Effects of endogenous cytokinin content on starch accumulation in root tuber of cassava[J]., 2016, 47(8): 1279-1284.(in Chinese with English abstract).
Effects of Embryo Morphology and Endosperm Composition on Embryo-remaining Characteristics in Rice
MA Zhaohui1,#, SHI Yihan1,#,CHENG Haitao1, SONG Wenwen1, LU Lianji1, LIU Renguang2, LÜ Wenyan1,*
(Agronomy College, Shenyang Agricultural University, Shenyang 110866, China; Benxi Agricultural Comprehensive Development Service CenterBenxi 117022, China; Corresponding author, email:)
【Objective】The embryo of embryo-remaining rice has high nutritional value and ensures the taste of rice. However, related biological mechanisms, such as the relationship among embryo-remaining characteristics, embryo traits, and endosperm components remains unclear. Therefore, it is necessary to further study the difference of embryo-remaining rate among different types of rice varieties and its influencing factors.【Method】Therefore, typical embryo-retaining and non-embryo-remaining varieties were selected from softand common rice as experimental materials to investigate rice embryo-remaining characteristics. Here, the embryo development characteristics of the two types of varieties were observed by slicing; then, apparent amylose content (AAC) and abscission pectin contents were compared. Finally, the differences in embryo-remaining rate, embryo traits and endosperm composition were studied under external cytokinin (CTK) application. 【Result】Compared with non-embryo-remaining varieties, embryo-remaining softvarieties and common rice had larger inner and outer embryo width, faster embryo development, large embryo volume, better grain plumpness, wider and thicker grain shape and higher abscission pectin content. After the application of CTK, the development speed of the soft rice varieties was accelerated in the middle and late stages of development, while the embryo development of the common varieties slowed down. The AAC, pectin content, embryo weight percentage and embryo-remaining rate of the two types were increased.【Conclusion】In general, apart from embryo traits, endosperm composition mainly affected the embryo-remaining characteristics of soft rice varieties, whereas grain shape was an important factor influencing the embryo-remaining characteristics of common varieties.CTK regulates the embryo-remaining rate of different types of rice varieties by influencing different traits of rice seeds. These results will lay a new theoretical basis for the breeding and cultivation regulation of high embryo-remaining rice varieties.
embryo-remaining rate; embryo development; endosperm composition; grain shape; cytokinin
10.16819/j.1001-7216.2023.220809
2022-08-25;
2022-12-26。
国家自然科学基金资助项目(32201892);辽宁省教育厅面上项目(LJKMZ20221016);辽宁省科技重大专项(2019JH1/10200001);沈阳农业大学成果转化项目(H2017035)。
