冬季和夏季达乌尔黄鼠下丘脑c-Fos、nNOS和NPY表达及血清激素水平比较
2023-05-16郑杨慧马小梅王建礼
郑杨慧,王 恒,马小梅,王建礼
(北方民族大学生物科学与工程学院,国家民委黄河流域农牧交错区生态保护重点实验室,银川,750021)
食欲是中枢和外周信号系统相互作用调节个体对营养摄入的反应[1]。营养物质引起的机械或化学信号可刺激胃肠激素分泌,肠道激素、胰岛素和脂肪激素通过血液循环或迷走神经运输,影响调控食物相关行为的关键脑区继而调节食欲。下丘脑被认为是食欲调节和能量稳定的关键结构,其中下丘脑弓状核(arcuate nucleus,ARC)、下丘脑室旁核(paraven⁃tricular nucleus,PVN)、下丘脑腹内侧核(ventrome⁃dial hypothalamic nucleus,VMH)、下丘脑外侧区(lat⁃eral hypothalamic area,LHA)以及下丘脑背内侧核(dorsomedial hypothalamic nucleus,DMH)合称为食欲调节中枢[2−5]。ARC 是控制食欲的中心单元,其他参与调节摄食行为的脑区位于ARC 下游[6]。PVN 分泌的促肾上腺激素释放激素(corticotropin releasing hormone,CRH)和促甲状腺素释放激素(thyrotropin releasing hormone,TRH)能够抑制食物摄入[1,5];LHA为饥饿或摄食中枢,VMH 为饱食中枢,受到刺激会抑制进食;DMH 与情绪调节有关,也能促进食欲。下丘脑通过整合来自血液循环或迷走神经的代谢、激素和神经元信号,调节神经肽诸如NPY(neuropep⁃tide Y)、食欲素(orexin,OX)等物质的释放继而调节食欲。同时,这些信号在更高级大脑中心被进一步整合,根据能量实时需求,产生进食或节食行为,维持能量摄入或支出平衡[3]。
NPY 是一种很强的促食因子,在机体摄食活动中发挥重要调节作用。当机体处于不同能量需求时,下丘脑NPY 能神经元可以直接调节摄食与消化功能,也可通过胃肠神经系统调节[4]。调节摄食的许多神经肽可通过上调或下调内源性一氧化氮(ni⁃tric oxide,NO)调节食物摄入量,因此,NO 成为神经肽调节食欲的核心成分[7−8]。由于NO 定位定量较难,可借助于神经元型一氧化氮合酶(neuronal ni⁃tric oxide synthase,nNOS)作为其生成标记酶。CRH除了在应激反应方面具有重要作用以外,也参与能量代谢,调节食欲[3],作为一种抗食欲调节因子,CRH 减少将会增加动物摄食行为。胃肠道激素也是食欲调节的重要信使,如胃饥饿素(ghrelin,GHRL)主要产生于胃部,是唯一的肠源性增食激素,具有促进食欲的作用,在调节食欲和能量稳态中扮演了关键角色[9]。c-Fos 是一种被广泛使用的中枢神经系统神经元激活的标志蛋白,对动物摄食也有一定指示作用[10]。研究表明外周和中枢注射GHRL 能提高下丘脑ARC、PVN 及DMH 脑区c-Fos表达[4]。
冬眠是一些动物在冬季出现的非活动状态,是长期进化过程中适应冬季寒冷和食物短缺等条件的行为与生理策略。达乌尔黄鼠(Spermophilus dauri⁃cus)是一种典型的贮脂类冬眠动物,在研究冬眠、肥胖及肌萎缩机制等方面被广泛使用,已成为研究适应极端条件生理机制的天然动物模型[11−14]。达乌尔黄鼠春季静止代谢率(resting metabolic rate,RMR)最高,秋季次之,夏季最低,冬眠期RMR 降到极低水平,仅为春季3%[15],体脂量在冬眠前大幅增加,体重增加近1倍[16]。研究阿片肽在达乌尔黄鼠冬眠调控的作用发现,脑中阿片肽含量具有明显季节差异[17],说明达乌尔黄鼠生理机能具有季节性适应。动物在冬眠时自发停止进食,能量需求降低,为了探明与食欲相关的神经内分泌因子在动物冬眠期和活动期发生了怎样的适应性变化,本研究检测了达乌尔黄鼠冬季(冬眠期)和夏季(活动期)下丘脑核团c-Fos、nNOS 和NPY 表达,同时检测了血清GHRL 和CRH浓度,以探讨中枢和外周信号在食欲调节方面的季节性适应。
1 材料与方法
1.1 试验动物
达乌尔黄鼠捕自宁夏盐池县惠安堡,分为夏季组(7 月上旬试验,共15 只)和冬季组(12 月下旬试验,共11只),皆为雄性成体。夏季组于6月捕获,冬眠组于冬眠前(10月)捕获,提供食物和饮水,在室内温度逐渐降为3~5 ℃,光照变为全黑条件下,达乌尔黄鼠会自发地进入冬眠状态[18−20]。
1.2 c-Fos、nNOS和NPY免疫组织化学实验
试验于09:00—11:00 进行,冬眠组在达乌尔黄鼠处于冬眠时取材,麻醉后开胸暴露心脏,左心室灌注0.01 mol/L PBS 缓冲液约150 mL,再以4%多聚甲醛灌注,取脑组织置于4%多聚甲醛液固定6 h,待脑组织沉底后换至30%蔗糖溶液中,待沉底后取出,做c-Fos、nNOS和NPY免疫组织化学试验。c-Fos 和nNOS 试验夏季用8 只达乌尔黄鼠,冬季用6 只,NPY试验夏季用7只,冬季用5只。
沉底的脑组织用组织包埋剂包埋,并在−24 ℃冷冻切片机上做厚度为30 μm 的冠状连续切片。用盛有0.01 mol/L PBS 缓冲液培养皿收集切片。将收集好的脑组织切片取出并放入反应槽后,滴加3% H2O2室温封闭10 min。去除H2O2后,用蒸馏水、0.01 mol/L PBS 缓冲液各洗1 次,每次5 min。滴加5%BSA,37 ℃封闭孵育1 h 后,加入一抗c-Fos(ab190289,1∶5 000,Abcam(香港)有限公司)或nNOS(ab5583,1∶500,Abcam(香港)有限公司)或NPY(ab30914,1∶1 000,Abcam(香港)有限公司),4 ℃孵育72 h。随后用0.01 mol/L PBS 缓冲液漂洗 5 min×3 次,滴加生物素化羊抗兔IgG(博士德生物 工程有限公司,武汉),37 ℃孵育1.5 h。再用 0.01 mol/L PBS 缓冲液漂洗5 min×3 次,滴加SABC试剂(武汉博士德生物工程有限公司),37 ℃孵育2.5 h,0.01 mol/L PBS 缓冲液漂洗10 min×4 次。用DAB 显色剂(武汉博士德生物有限公司)显色,最后使用梯度乙醇脱水,二甲苯透明,中性树胶封片。
各脑区参照Paxinos等[21]的小鼠脑图谱定位,使用Motic Panthera U 数码显微镜在10×10 倍镜下观察、计数和拍照。每只鼠各核区选择3 张连续切片量化,利用显微测微尺,计算相同面积内单侧脑区核团免疫活性神经元数目,包括下丘脑ARC、PVN、VMH、LHA及DMH脑区。
1.3 血清激素检测
达乌尔黄鼠麻醉后心脏取血,血液收集于离心管中,4 ℃静置1 h 后,3 000 r/min 离心15 min,吸取上清液放置在标记好的EP 管中,冰箱−20 ℃保存。购买GHRL(CEA991Ra)和CRH(CEA835Mu)ELISA检测试剂盒(武汉云克隆科技股份有限公司),按照试剂盒操作说明检测。由于部分达乌尔黄鼠采血量不足及各激素检测所需血清量不同,因此实际检测激素样本量有所不同(GHRL:夏季n=8,冬季n=8;CRH:夏季n=6,冬季n=6)。
1.4 数据统计分析
所有数据用SPSS 20.0处理,冬季和夏季免疫活性神经元数目及血清激素浓度差异采用独立样本t检验比较,p<0.05为差异显著。
2 结果
2.1 c-Fos-IR、nNOS-IR和NPY-IR神经元表达
冬季与夏季相比,c-Fos-IR 神经元密度在ARC(t12=2.863,p<0.05)、PVN(t12=7.455,p<0.001)、VMH(t12=4.266,p<0.01)、LHA(t12=2.424,p<0.05)和DMH(t12=3.232,p<0.05)表达减少(图1A,图2)。nNOS-IR 神经元密度在PVN(t12=6.474,p<0.001)、VMH(t12=3.461,p<0.05)、LHA(t12=4.332,p<0.001)和DMH(t12=3.896,p<0.05)显著减少,但在ARC 表达没有显著差异(t12=1.562,p>0.05)(图1B,图3)。NPY-IR 神经元在ARC(t10=7.056,p<0.001)和VMH(t10=9.253,p<0.001)表达显著减少,但在LHA 的NPY-IR 神经元数目显著高于夏季(t10=5.125,p<0.001)(图1C,图4)。
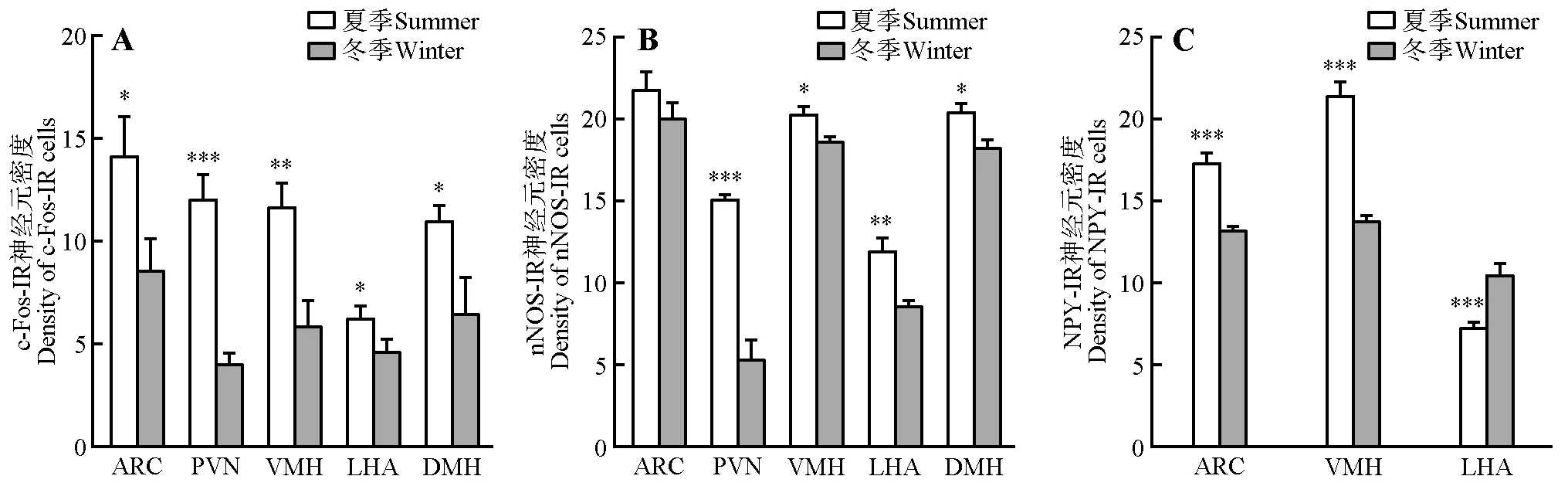
图1 冬季和夏季达乌尔黄鼠下丘脑c-Fos-IR、nNOS-IR和NPY-IR神经元密度Fig.1 Density of c-Fos-IR,nNOS-IR and NPY-IR neurons in the hypothalamus of Daurian ground squirrels in winter and summer
图2 冬季和夏季达乌尔黄鼠c-Fos免疫组织化学染色Fig.2 Immunohistochemical localization of hypothalamic c-Fos in Daurian ground squirrels in winter and summer
图3 冬季和夏季达乌尔黄鼠下丘脑nNOS免疫组织化学染色Fig.3 Immunohistochemical localization of hypothalamic nNOS in Daurian ground squirrels in winter and summer
图4 冬季和夏季达乌尔黄鼠下丘脑NPY免疫组织化学染色Fig.4 Immunohistochemical localization of hypothalamic NPY in Daurian ground squirrels in winter and summer
2.2 血清GHRL和CRH水平
冬季达乌尔黄鼠血清GHRL 浓度显著低于夏季(t14=5.295,p<0.001),但CRH 浓度高于夏季(t10=3.558,p<0.01)(图5)。
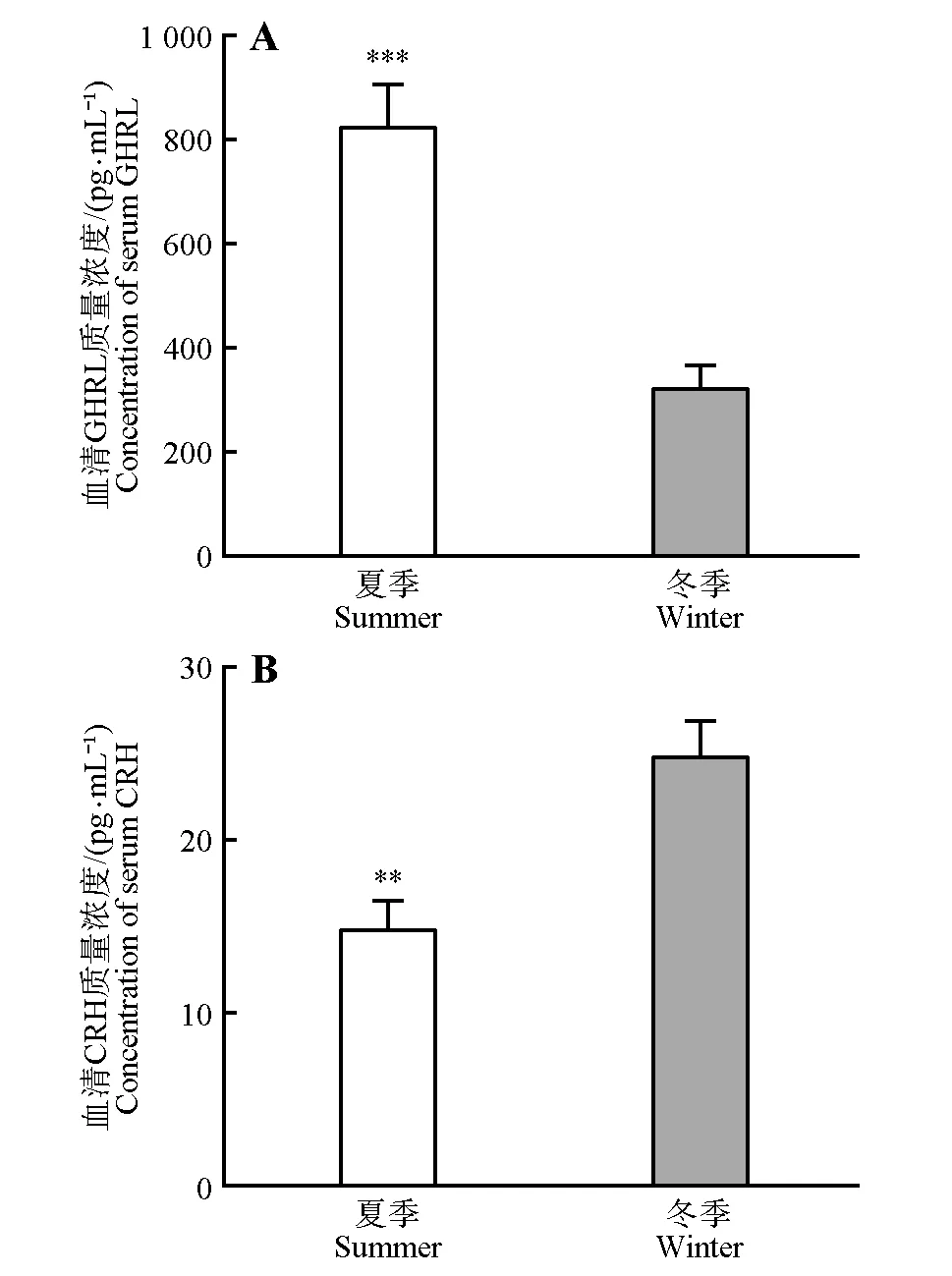
图5 冬季和夏季达乌尔黄鼠血清GHRL和CRH质量浓度Fig.5 Concentrations of serum GHRL and CRH in Daurian ground squirrels in winter and summer
3 讨论
c-Fos、nNOS 和NPY 神经元广泛分布于下丘 脑各区[4,22−23]。本研究结果显示,达乌尔黄鼠冬季 c-Fos-IR神经元密度在ARC、PVN、VMH、LHA及DMH区域低于夏季活动期;除ARC外,冬季nNOS-IR神经元密度在这些核团也明显减少;NPY-IR 神经元密度在ARC和VMH明显低于活动期,这些结果暗示了达乌尔黄鼠下丘脑神经元活动、NO及NPY合成对冬眠期的适应。
c-Fos 作为神经元激活标志,冬季达乌尔黄鼠下丘脑各核团c-Fos-IR 神经元较少,暗示了下丘脑神经元活动变弱,这与冬季处于休眠状态有关。与此结果相似的是,冬季达乌尔黄鼠嗅球神经元活动也低于其他季节[20]。NO参与中枢神经系统摄食调节,增加NO 会增加食欲[24]。NPY 具有增强食欲和调控脂肪合成和储存的作用,主要分布于下丘脑ARC,并有神经纤维投射到VMH 和LHA 形成的NPY 网络,在机体摄食活动中发挥重要调节作用。当机体处于不同能量需求时,下丘脑NPY 能神经元可以直接调节摄食与消化功能,也可通过胃肠神经系统调节[25]。李艳莉等[26]对棕色田鼠(Lasiopodomys mandarinus)的研究发现,VMH 区NPY 对动物摄食和能量平衡具有重要调节作用,在能量需求增加或能量供应不足时,可激活下丘脑NPY 能神经元,刺激食欲,增加摄食量。达乌尔黄鼠在冬眠时能量需求降低,nNOSIR和NPY-IR神经元表达的降低暗示了食欲的抑制;相反,夏季能量需求增加,通过兴奋下丘脑内NPY能神经元可提高食欲,增加摄食量,如对达乌尔黄鼠和大鼠尾蝠(Rhinopoma microphyllum)的研究表明,NPY 表达增加有助于冬眠前脂肪和体重增加[16,27]。本研究发现冬季LHA 区NPY-IR 表达量高于夏季,由于NPY 还参与胃肠功能调节,具有抑制小肠蠕动和胃肠转运的作用[28−29],进入冬眠后达乌尔黄鼠几乎不摄食,高表达的NPY 可能与抑制胃肠功能有关,考虑到冬眠期达乌尔黄鼠胃几乎是空的,LHA活动增加也是饥饿状态的一种暗示[30]。在整个冬眠期,冬眠动物从入眠、深冬眠、激醒再到体温恢复正常为一个冬眠阵,之后再进入下一个冬眠阵[12]。达乌尔黄鼠冬眠期的冬眠阵平均为7.55 个,阵间激醒时长平均为1.36 d[19]。LHA 虽是摄食中枢,同时也与觉醒有关[31−32],LHA 的OX 参与睡眠-觉醒调节。NPY 对觉醒和睡眠具有双向调节作用[33],NPY 通过突触前和突触后机制能够抑制OX神经元活动,继而调节觉醒反应[34]。因此,冬季LHA 区NPY-IR 高表达不能排除与阵间激醒的相关性。
GHRL 是一种能够促进食欲的胃肠激素,通过血脑屏障作用于中枢系统,具有调节能量平衡、睡眠、食欲和胃肠道蠕动作用[35]。GHRL 通过与中枢食欲调节神经元受体或外周代谢器官受体结合发挥作用。Alamri等[9]研究表明禁食会使血浆GHRL 升高,继而刺激摄食行为,贮存脂肪。本研究结果显示,夏季GHRL 含量高于冬季。作为一种促食因子,GHRL 可促进活动期达乌尔黄鼠摄食。冬季达乌尔黄鼠自发地停止摄食,处于冬眠状态,与胃肠活动减弱相适应,GHRL 合成和分泌也相应地减少。CRH是一类抑食因子,同时也参与能量调节[5]。CRH 与受体结合后可通过抑制胃排空,增加饱腹感,降低食欲[36]。中枢或外周注射CRH,摄食量会显著减少[37−38]。ARC 增食和抑食神经元投射到PVN,PVN产生抑食因子,如CRH 和TRH 等抑制食物摄取[1,5]。尽管外周激素水平不能表示其在中枢的水平,但在一定程度上能够反映对环境刺激的响应[39]。CRH作为一种抑食因子,冬眠期达乌尔黄鼠血清CRH 浓度高于活动期,可能与冬眠期停止摄食有关。先前研究冬眠和觉醒过程中达乌尔黄鼠和多纹黄鼠(Spermophilus tridecemlineatus)甲状腺素发现,血清T3和T4水平明显高于非冬眠鼠[40−41]。此外,由于CRH在应激反应中的突出作用,高水平CRH 也可能与冬眠阵中冬眠激醒有关。
下丘脑监测机体稳态并调节不同行为,诸如摄食、产热和睡眠等。冬季和夏季达乌尔黄鼠处于不同能量需求时,下丘脑NO 和NPY 能神经元激活程度不同。夏季达乌尔黄鼠处于活动状态时摄食频繁,下丘脑神经元活跃,作为促食因子,NO与NPY高表达促进了食欲并有利于体内脂肪积累。冬季与夏季外周GHRL 和CRH 浓度变化并不一致,提示了二者在季节适应中的作用不同,该结果有助于认识冬眠动物摄食生理的季节性调节。
