太行山猕猴寰椎桥的类型与特征
2023-04-29赵晓进郭梦丹张梦琪鹿雪涵
赵晓进 郭梦丹 张梦琪 鹿雪涵

摘要:为了确定太行山猕猴(Macaca mulatta) 的椎动脉沟上3 种骨桥(腹桥、侧桥和背桥)的分布特征,以期理解在人类寰椎上观察到的变异,并推测灵长类动物可能的进化趋势。本文分别测量57 例(雄性17 例,雌性40 例)猕猴寰椎标本(成年骨骼标本)3 个寰椎桥的宽度。并采用SPSS 统计软件进行数据处理,性别之间差异采用方差分析,观察寰椎桥的出现率、形态特征和组合类型。统计结果显示,猕猴寰椎桥变量性差显著(P<0.05),猕猴的腹桥和背桥是恒定出现(100%),侧桥基本恒定,但也有一些缺失个案(90.3%),表明腹桥和背桥是猕猴稳定的特征;猕猴寰椎桥的主要类型为原始的A 型,与主要类型为D 型的人科动物发生分离。我们基于以上特征后得出结论,猕猴的3 个寰椎桥是一种稳定性状,其出现率明显高于类人猿。
关键词:寰椎;骨桥;猕猴;灵长类
1 引言
人类寰椎的正常变异和临床意义受到国内外学者的关注,寰椎作为颈部的重要解剖结构也愈加受到重视。欧海宁等[1] 通过大数据分析了人类寰椎后桥的出现率和临床表现后得出结论:发生在人类寰椎后桥是颈椎骨质结构的解剖学变异,寰椎后桥的出现及解剖学特征与临床症状有一定的相关性。了解寰椎桥的结构、特征、起源和种群差异,有助于了解灵长类动物的个体发育和系统演化,探讨灵长类动物可能的进化趋势[2]。
薛德明等曾对太行山猕猴寰椎和枢椎进行了初步研究[3,4]。肖洪文等针对人类寰椎桥的测量、分型、变异和临床意义等展开系列的研究,丰富了寰椎桥的研究内容,推进了寰椎桥的研究进展[5]。Le Minor 等观察了部分灵长类动物的寰椎标本,探讨寰椎桥在灵长类动物中的分布和类型,试图解释人类观察到的寰椎桥变异[6]。目前,对于在人类种群中出现寰椎桥变化,出现寰椎桥变化的真正原因尚不清楚,提出的解释以理论性为主。Yammoto 等对328 只日本猕猴寰椎桥的研究表明,日本猕猴的寰椎主要类型是背桥,侧桥的出现率略低于背桥[7]。推测寰椎背桥是日本猕猴的一个稳定性状,其背桥出现率明显高于人类。尽管人们已经认识到寰椎桥结构在人类进化过程中的重要性,但迄今为止,国内关于非人灵长类动物寰椎桥的研究一直较少。
本文研究目的在于观察太行山猕猴寰椎桥的形态学特征和分布,描述性统计其性别差异,探讨不同灵长类动物寰椎桥的种群差异和进化趋势。
2 材料与方法
2.1 研究材料
本文的研究材料为太行山猕猴寰椎标本57 件(雄17,雌40),均按照标准制备方法完成。标本完整、对称、无任何病例征象(图1)。根据齿式和骨骺愈合可以确认标本是成年的,性别是已知的。标本收集时间是1998~2022 年,标本全部来自河南省太行山猕猴自然保护区。目前保存在郑州工商学院商学院。
2.2 寰椎桥的定义和分类
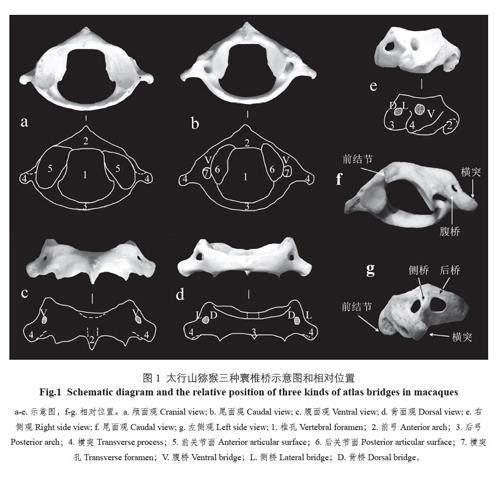
根据文献对寰椎桥进行定义,理论上寰椎椎动脉沟上可能存在3 种骨桥,分别是腹桥、侧桥和背桥[6,7](图1: a-e)。腹桥由寰椎横突孔的腹侧结构构成,椎动脉通常在此通过,部分对应于横突的前根,是颅面横突孔与尾面横突孔在腹面观的直线距离,腹桥如果缺失一般认为是横突前根缺失。侧桥位于寰椎侧块的外侧,椎动脉沟上方,文献中常常会出现各种不同的命名(侧骨桥、外侧桥等);从寰椎的颅面观上看得清楚,也容易测量,是横突孔与横向上孔之间的直线距离。背桥(人类寰椎上通常称为后桥)位于寰椎侧块的背侧,椎动脉沟的上方,在寰椎的背面观上看得很清楚,是横向上孔与侧块上缘的直线距离。三种寰椎桥的相对位置见图1: f-g。
依据是否出现寰椎桥,可以有8 种组合类型,记作A、B、C、D、E、F、G、H(表1)[6]。
2.3 研究方法
本文对寰椎桥进行定量分析,采用人类体质学骨骼直接测量法。测量仪器为国际标准电子数显卡尺,单位是mm,精确到0.01 mm。所有标本的测量均由本文第一作者独立完成。变量选择是参照文献资料[8],测量变量为三种寰椎桥的桥宽。数据处理采用IBMSPSS 26.0 统计软件。如果侧别之间没有统计学差异,性别组间统计检验可以采用两侧合并数据 [9,10]。性别组间检验采用单因素方差分析(Variance analysis, VA)。
3 结果
3.1 寰椎桥描述性统计
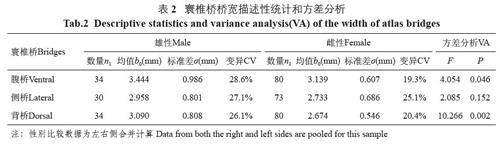
太行山猕猴3 种寰椎桥测量性状性别组间的比较结果表明,腹桥桥宽的F=4.054,P=0.046;背桥桥宽的F=10.266, P=0.002, 性别组之间腹桥和背桥有统计学差异(P<0.05);侧桥桥宽的F=2.085, P=0.152, 性别组间没有统计学差异;3 个寰椎桥变量都是雄性大于雌性(表2)。
3.2 猕猴寰椎桥的出现率
太行山猕猴3 种寰椎桥的出现频数和百分比统计结果显示:猕猴的3 种寰椎桥出现率有一定的规律。腹桥和背桥出现稳定,出现率为100%;侧桥出现率略低,双侧合并后出现率为90.4%,双侧同时出现率为86.0%(表3)。
从非测量性状来看,首先是寰椎桥的数目不同,猕猴一般在寰椎侧块上有3 个骨桥,分别称为腹桥、侧桥和背桥。寰椎桥的数目差异是区别人类与猿类的关键。从形状和大体解剖结构来看,人类与猿类的寰椎桥非常相似,但有一个明显的差异,即猕猴的寰椎颅面观一般看不到横突孔,尾面观横突孔显著,基本上是圆形。在猕猴的寰椎腹面观可以看到一个比较小的横突孔(图1)。综上所述,太行山猕猴的横突孔有两个特征,一是横突孔在颅面观和尾面观不是贯通的;二是颅面观侧块上的横突孔一般比尾面观上的横突孔要小,而且形状不规则,有时侧桥和腹桥会发生融合,导致该孔消失。
猕猴腹桥在颅尾侧也不是扁平的,腹桥的宽几乎比横突上向尾面伸出的脊还要小。其重要原因是位于颅面观上的横突孔比较小且不规则,和人类的腹桥形态完全不同。猕猴侧桥形状极不规则,有时很难辨认,只能通过横突侧面上的两个孔来识别,桥宽用两个孔之间的最短距离来表示。本研究中侧桥的缺失率较高,说明变异性较大。侧桥缺失的原因可能是侧桥在颅面观的横突孔一般大小不等,两侧一般不对称,有时会在单侧或双侧消失,所以,侧桥和腹桥会融合形成较大的腹桥,猕猴寰椎背桥是猕猴类的典型特征,形状规则、扁平、稳定,容易测量和识别。
3.3 不同种群灵长类动物寰椎桥比较
由表4 可知,寰椎桥的出现率有明显的差异。婆罗洲猩猩(Pongo pygmaeus) 腹桥出现率为95.8%,侧桥出现率为0,背桥为66.7%。人类(Homo sapiens) 腹桥出现率为92.6%, 侧桥为1.0%,背桥为10.1%。在原猴猴亚目(Strepsirhine)、蛛猴亚科(Atelinae) 和绢毛猴科(Callitrichine) 中,侧桥却是恒定出现,或者几乎稳定出现(99.0%~100%)。猩猩和人类中,后桥出现率明显减少,分别为66.7% 和10.1%;侧桥的出现率更低,分别为0.0% 和1.0%。太行山猕猴(Macaca mulatta) 中,腹桥出现率为100%,侧桥为90.3%,背桥为100%。在猕猴亚科(Cercopithecinae) 中,腹桥出现率为100%,侧桥为91.5%,背桥为98.9%。在所有的灵长类种群中,除了猩猩和人类外,背桥出现率都非常高,范围在98.9 %~100%(表4)。
不同灵长类种群寰椎桥类型比较见表5。灵长类动物中的寰椎桥类型有8 种,其中主要类型有3 种,分别是A 型、C 型和D 型。
A 型 3 种寰椎桥同时存在,是原猴亚目(Strepsirhini) 和猴科(Cercopithecines) 的主要类型,在黑猩猩(Pan troglodytes)、长臂猿(Hylobates) 和人类(Homo sapiens) 中偶然出现,而在婆罗洲猩猩(Pongo pygmaeus) 和大猩猩(Gorilla gorilla) 中不曾出现。太行山猕猴主要类型是A 型。
C 型 腹桥和背桥同时存在,侧桥不存在,是婆罗洲猩猩(Pongo pygmaeus) 和黑猩猩(Pan troglodytes) 的主要类型,在长臂猿(Hylobates) 的出现率为33.3%,在人类(Homosapiens) 和猴科(Cercopithecines) 中偶然出现(出现率分别是9.0% 和5.7%),在大猩猩(Gorilla gorilla) 中不存在。
D 型 只存在腹桥,不存在侧桥和背桥,通常仅在一些类人猿物种中占主导地位。猕猴(Macaca mulatta) 和猴科(Cercopithecines) 动物中的D 型十分罕见或缺失(0.0%~0.3%)。
H 型 3 种寰椎桥同时消失,在大猩猩(Gorilla gorilla)、黑猩猩(Pan troglodytes) 和人类(Homo sapiens) 中偶然出现,出现率在6.9% ~11.9% 之间(表5)。
4 讨论
将寰椎桥简单地确定为存在或缺失,显然不能完全反映出骨骼形态特征所表达的生物学意义[6]。孙静宜等对人类寰椎后桥进行了形态学分析后认为,背桥是人类进化过程中的退化痕迹,狭窄的寰椎动脉沟环可能是造成颈性眩晕的主要原因[2]。Yammoto 等研究了日本猕猴寰椎桥个体变化和地理变异,同时证明日本猕猴寰椎桥与年龄之间的对应关系[7]。研究结果表明,成年日本猕猴背桥和侧桥普遍存在。Kikuchi 等研究发现,不同灵长类动物颈椎的形态不同,通过寰椎的体型指数和形态指数可以推测其种群的个体大小和运动类型,进一步可以推测他们的进化程度[11]。
从测量性状来看,寰椎桥侧别之间没有统计学差异;性别之间腹桥和背桥有统计学差异,差异最大的变量是背桥宽。寰椎桥性差的特征可以解释为种群遗传因素和系统进化的结果,是其种群生物学属性之一,是长期性别选择和青春期后雌雄两性异速生长的结果[12]。侧桥宽性差不显著,可能与侧桥的变异性较大有关。侧桥宽的变异系数是25.90%,极差是3.66 mm;对应背桥宽的变异系数是23.58%,极差是3.13 mm。关于寰椎骨桥的测量性状的意义,可以解释为这是解剖学和人类学常用的方法。很多文献资料也对人类和非人灵长类寰椎的形态进行定量和定性研究[2,4-6]。尽管大多数骨桥的测量是以人类寰椎为对象,但猕猴是人类的近亲,寰椎桥的结构,尤其是背桥是相似的,资料也显示人类的寰椎后桥与非人灵长类的寰椎背桥是同源的。另外,通过对骨桥宽度的定量分析,还可以得到很多有价值的信息。例如,可以进行判别分析、多元回归分析和种间定量分析等。
从系统发育的角度来看,这三个寰椎桥在灵长类动物和其他哺乳动物中具有一定的同源特征,深入研究不同动物类型的寰椎骨桥,对于进一步理解动物的系统发育具有重要意义。目前,国内这一领域研究结果相对较少,有待进一步加强。
从非测量性状来看,太行山猕猴通常具有稳定的3 种寰椎桥结构,其中背桥和腹桥的出现率均为100%。侧桥偶有缺失,缺失率往往超过10.0%;这一现象表明,太行山猕猴侧桥的结构并不稳定。侧桥缺失的主要原因在于寰椎颅面观的横突孔一般大小不等,两侧的横突孔也总是不对称的。另外,侧桥的形状极不规则,一般是通过寰椎横突脊侧面的两个小孔来确认,在寰椎横突截面上的侧桥会构成锥形。有时由于侧桥和腹桥融合,形成较宽的腹桥,这种特殊的寰椎腹桥有待进一步观察。太行山猕猴寰椎后桥是猕猴属的稳定结构,这一点与日本学者Yammoto 等对日本猕猴寰椎桥的研究结果是一致的[7]。
探讨不同灵长类动物寰椎桥的起源、种间差异、影响因素和进化趋势是有意义的。参照以前的研究资料可以推测,太行山猕猴寰椎桥的结构是一种相对原始的结构,其主要依据是:1) 许多哺乳动物中出现了类似猕猴寰椎桥的结构,与人类的寰椎桥结构完全不同[5];2) 从表4 可以看出太行山猕猴(Macaca mulatta) 的3 种寰椎桥的出现率与一些低等灵长类动物的十分相似,如原猴亚目(Strepsirhini)、蜘蛛猴亚科(Atelines) 和绒猴亚科(Callitrichines) 等[5]。
非人灵长类动物的三个寰椎桥同时存在为原始类型(A 型)。在灵长类动物中能够观察到的从原始类型(A 型)以及派生出来的6 种寰椎桥的类型(表1)。A 型主要出现在原猴亚目(Prosimians) 的一些科和属、阔鼻类(Platyrrhines) 的一些科、猴科(Cercopithecinae)以及猕猴属(Macaca mulatta) 等。由原始类型(A 型)派生出来的其它类型包括3 个层次,第一层次是C 型(只有腹桥和背桥),这一类型主要包括跗猴科(Tarsiids, 100%)、疣猴属(Colobus, 85%) 和猩猩属(Pongo pygmaeus, 62.5%);第二层次是D 型,这一层次主要包括大猩猩(Gorilla gorilla, 89.5%)、人科(Homo sapiens, 82.8%) 和长臂猿属(Hylobates,63.9%) 等;第三层次是H 型,这一类型只在人猿超科中出现,大猩猩(10.5%)、黑猩猩(Pantroglodytes, 11.9%) 和人科(6.9%)。
从进化趋势上来看,可以将所有的灵长类动物大致分为3 个组。1) 原猴亚目- 阔鼻类-猴科组,主要是原始类型A 型;2) 疣猴属- 猩猩属- 黑猩猩属组,主要是由A 型派生出来的C 型;3) 长臂猿- 大猩猩- 人科组,主要是由C 型派生出来的更高级别的D 型。从整体结构上看,与人类最近似的猿类就是大猩猩,这点确定无疑[13]。总之,寰椎桥的逐渐消失似乎构成了灵长类动物的一种进化趋势。
考虑到灵长类的系统发育,各种寰椎桥类型之间也存在着内在的演变关系。例如,原猴类被公认是现存灵长类动物中最原始的,寰椎桥表现出原始类型(A 型)。眼镜猴科(Tarsiidae) 属灵长目简鼻亚目跗猴型下目的一个科,出现于距今6000 万年以前,一直延续到现代,是一支高度特化的灵长目动物。简鼻亚目与原猴亚目约于6300 万年前演化分开,所以眼镜猴科会有一些原猴亚目的特征,同时也混合了类人猿的一些特征。例如,眼镜猴的婴儿相较于原猴更大,但较依赖母亲,这可能是因它们复杂的行为及自然历史所致。眼镜猴科的寰椎桥也确实表现出介于原猴类与类人猿之间的模式(C 类)。
本文对观察到的太行山猕猴寰椎桥的分布也可以进行解释。寰椎桥主要类型为原始的A 型,同时,出现少量的C 型,没有B 型,因此推断太行山猕猴寰椎桥介于原始灵长类动物与高等灵长类动物之间,与Le Minor 等和Yammoto 等研究结果一致[6,7]。类人猿亚目也明显表现出寰椎桥的多态性,黑猩猩的寰椎桥的多态性似乎比人类要更大一些。寰椎桥的多态性曾一直被人们所忽视,实际上寰椎桥多态性对于物种的形成或进化具有特殊的意义,是物种进化和生物多样性形成的基础。
有关寰椎桥的起源一直存在争议。部分学者认为是哺乳动物先天正常发育的结果,特别是翼手目和食虫目颈椎上斜突的同源物。Yamamoto 等却认为寰椎背桥是由寰枕后膜骨化形成,很明显这种骨化过程与年龄增长有关[14]。Traize 等的研究进一步证明青年人似乎是以部分不完整的桥为主,而老年人主要以完整桥为主;部分桥可能是完整桥的前体[8]。目前,还没有证据证明,成年人的寰椎背桥会随着年龄的不同而发生变异。本研究观察发现,幼年太行山猕猴的寰椎标本上的确可以看到不完整的背桥,但更像是动物正常发育的过程;成年太行山猕猴有着完整稳定的背桥结构(100%),而这似乎与衰老过程无关;寰椎背桥应是猕猴的一种正常结构,具有种遗传属性,是系统演化和长期适应环境的结果。
本文依据Finnegan 等提出的寰椎桥分类方法进行统计描述[15]。这是一种新的尝试,也产生了一些新的认识,概述如下:1) 简单地将寰椎桥分为存在与缺失似乎与实际情况不符,因为在寰椎桥存在一些中间过渡类型(例如半桥)。Taitz 等的研究结果显示人类不同种群的部分桥出现率不同,白人是1.4%~5.2%,黑人是6.0%~10.4%,印度人是37.4%,中东人是55.2%,总体上来看,部分桥或不完整桥的出现率是25.9%[8]。太行山猕猴寰椎桥的观察中也发现了这种情况,值得深入研究。2) 将非人灵长类动物的寰椎桥定义为8 种类型是为了进行研究对比,并非唯一的选择;一些寰椎桥根本不出现,或者一些桥类很难归类。 3) 除了考虑是否存在外,还要考虑寰椎桥非测量性状的形态特征。
例如,太行山猕猴3 种寰椎桥的形态各异,腹桥宽扁且向上隆起,侧桥形状极不规则,桥面向上突起构成横突,这些桥之间的空间非常小,形成了一个微小的孔。4) 关于寰椎桥出现率是否存在性别差异,尚无定论的文献资料。有资料显示人类的侧桥不存在性别差异。对于背桥的研究,不同学者的研究结果不同[3-5]。5) 全面研究灵长类动物的寰椎形态特征,可以让我们得到更多有价值的信息。例如,人类寰椎背桥的出现很可能是一种返祖现象,因为这种结构在低等的原始灵长类动物的寰椎上普遍存在;另外,通过寰椎一些变量可以计算出体型大小指数,可用于推算灵长类动物的体型大小,还可以计算出相应的形状指数,去推测不同灵长类动物的形状差异,包括行为和运动类型[8]。最后,本文作为一项基础性研究,可以为国内同行进行形态比较和种间比较,为考古工作者鉴定灵长类动物化石等提供相关参考资料[14]。
参考文献
[1] 欧海宁,黄志明,刘向华,等.寰椎后桥X 线表现及临床意义[J].现代医用影像学,2018, 27(3): 705-709
[2] 孙静宜,张琼珍,叶静.寰椎椎动脉沟环与颈性眩晕[J].中华外科杂志,1990, 28 (10): 592-594
[3] 薛德明,赵晓进,郭玫,等.太行猕猴寰椎的测量及相关性研究[J].河南师范大学学报,1996, 24(4): 101-103
[4] 薛德明.太行山猕猴寰椎和枢椎的初步研究[J].动物学杂志,2003, 38(2): 74-76
[5] 肖洪文,吴国雄,张西北,等.寰椎变异的观测I 中国人寰椎后桥和侧桥的观测[J].泸州医学院学报,1990, 12(6): 416-422
[6] Le Mino JM, Trost O. Bony bridge of the atlas (C1) over the groove for the vertebral artery in humans and primates: Polymorphism
and evolutionary trends[J]. American Journal of Physical Anthropology, 2004, 125: 16-29
[7] Yamamoto A, Kunimatsu Y. Geographic variation of atlas bridging of Japanese monkey (Macaca fuscata) [J]. Primate Research,
2002, 18: 215-224
[8] Taitz C, Nathan H. Some observations on the posterior and lateral bridge of the atlas[J]. Acta Anatomica, 1986, 127: 212-217
[9] 赵晓进,王凤产,田华香,等.猕猴掌骨和指骨长度性差[J].解剖学报,2017, 46(5): 673-678
[10] Barrio PA, Trancho GJ, Sanchez JA. Metacarpal sexual determination in a Spanish population[J]. Journal of Forensic Sciences,
2006, 51(5): 990-994
[11] Kikuchi Y, Nakano Y, Nakatsukasa M, et al. Functional morphology and anatomy of cervical vertebrae in Nacholapithecus kerioi, a
middle miocene hominoid from Kenya[J]. Journal of Human Evolution, 2012, 62: 677-695
[12] 赵晓进,韩霄帆,王凤产.猕猴胸骨柄性差与锁骨、股骨长相关性分析[J].解剖学报,2019, 50(2): 123-126
[13] Huxley TH.人类在自然界的位置[M].蔡重阳,王鑫,傅强(译).北京:北京大学出版社,2010, 41-42
[14] Yamamoto A, Kunimatsu Y. Ontogenetic change and geographical variation of atlas bridging in Japanese Macaques (Macaca
fuscata) [J]. Anthropological Science, 2006, 114: 153-160
[15] Finnegan M. Non-metric variation of the intracranial skeleton[J]. Journal of Anatomy, 1978, 125: 23-37
