雌雄异株树种山杨、水曲柳和东北红豆杉光合特性对比
2023-03-30孙晓伟王兴昌孙慧珍全先奎杨青杰
孙晓伟,王兴昌,孙慧珍*,全先奎,杨青杰
(1.东北林业大学林学院生态研究中心,森林生态系统可持续经营教育部重点实验室,黑龙江 哈尔滨 150040;2.东北林业大学功能性草业工程技术研究中心,黑龙江 哈尔滨 150040)
树木为了适应自然界,逐渐演化出多种繁育系统。其中雌雄异株树木由于繁殖成本的不同形成性别特化及其各自的形态、生长和空间分布特征,这对于维持森林生态系统结构和功能稳定性发挥着重要作用[1]。光合作用是树木将太阳能转为化学能并储存在有机物中的唯一自然途径,不仅是雌雄异株树木生长、繁殖及防御等一切生命活动的物质基础和能量来源,也是评判不同性别树木生长适应性的重要指标。因此雌雄异株树木光合特性作为最基本的生理现象之一,一直被学者广泛关注。
雌雄植株光合特性受到诸多内部和外部因素共同制约,因树种和环境而异[2]。目前雌雄异株树木光合特性研究多以杨属(Populus)和柳属(Salix)等繁殖前期的温室盆栽幼苗幼树为材料。对比温室幼苗在最适温湿度、大气CO2浓度和饱和光强时净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)或水分利用效率(EWUE)等指标,呈现雌株高于雄株[3-4]、雌株低于雄株[5-6]、两者结果相似[7-8],或性别间同一光合指标比较结果截然相反[3,6]等情况。产生以上分歧的原因可能与幼苗光合指标变异较大有关,因此尚未进入繁殖期的幼苗气体交换特征很难代表成龄树性别遗传型之间表现出来的差异。已有研究表明,在野外自然生境条件下,除檀香科(Viscaceae)植物Phoradendronjuniperinum[9],成龄树木如北极柳(S.arctica)[10]、梣叶槭(Acernegundo)[11-12]、窄叶杨(P.angustifolia)[13]、杞柳(S.integra)[14]、中国沙棘(Hippophaerhamnoidessubsp.sinensis)[15]、连香树(Cercidiphyllumjaponicum)[16]、鼠李(Rhamnusdavurica)[17]等均为雌株的气体交换参数不同程度地高于雄株。这说明雌雄异株树木光合生理过程除了受控于树木本身生长发育阶段,还可能与温室和野外环境条件不同有关[18]。例如生长于地中海气候区的乳香黄连木(Pistacialentiscus)在室内适宜条件下,雌雄株光与CO2响应曲线得到的参数相似(P>0.05);然而野外自然生长、处于水分胁迫的雌株EWUE高于雄株44.4%,雌株Pn、Gs和Tr的最大值低于雄株40.4%~52.4%(P<0.05)[19]。有些研究者为了尽量减少试验条件或材料的影响,将原状土上着生的野生幼苗装入盆中,在原地经过1个生长季后,于休眠期运回生长室[20]或采集海拔2 300 m处枝条,盆栽后置于不同海拔的自然生境中开展试验[21]。
山杨(Populusdavidiana)、水曲柳(Fraxinusmandshurica)和东北红豆杉(Taxuscuspidata)是东北东部山区地带顶极群落——阔叶红松林不同演替时期的重要伴生树种。以上雌雄异株树种的种群性别比、年龄结构、树干径向生长和生殖力[22-25]的差异可能与光合特性有关。虽然前人对以上树种的光合特征进行了相关研究[26-29],但因树木生长发育早期或野外成龄树未处于开花结果期,无法通过形态特征鉴定其性别。为此,本研究以野外自然生长的成龄树为研究对象,通过开花标识性别,比较不同树种雌雄植株气体交换参数。研究结果不仅有助于深入了解山杨、水曲柳和东北红豆杉雌雄异株光合生理生态方面的生物学特性,还可为正确评价该类树种对环境的适应机制以及确定影响种群建成的因素提供理论参考。
1 材料与方法
1.1 研究区概况
山杨(Populusdavidiana,简写为Pd)和水曲柳(F.mandshurica,Fm)的采样地位于黑龙江省尚志市帽儿山森林生态系统定位站(126°20′~126°25′E,45°20′~45°35′N),东北红豆杉(T.cuspidata,Tc)的采样地位于黑龙江省穆棱东北红豆杉自然保护区(130°00′~130°28′E,43°49′~44°06′N)。以上两个采样地主要土壤类型均为暗棕壤,气候类型均属大陆性温带季风气候。帽儿山生态系统定位站平均海拔400 m,年平均气温2.8 ℃,最高和最低气温分别为32、-31 ℃,无霜期130 d;年平均降水量629 mm,降水主要集中在7—8月,全年平均蒸发量为884 mm。主要植被为阔叶红松林经过不同程度的干扰(采伐、经营、火烧和开垦等)后形成的天然次生林和人工林。穆棱东北红豆杉自然保护区平均海拔600 m,年平均气温3.5 ℃,最高和最低气温分别为21.8、-18.3 ℃,无霜期126 d;年平均降水量530 mm,降水主要集中在7月,主要植被是以红松为主的温带针阔混交林。
1.2 试验材料
山杨和水曲柳均为先花后叶植物,东北红豆杉大、小孢子叶球同期开花。观察山杨、水曲柳(4—5月)和东北红豆杉(5—6月)繁殖器官,确定雌雄植株。选取生境、胸径相似的同一树种雌树和雄树各6株作为样树,山杨、水曲柳和东北红豆杉胸径范围分别为(23.9±0.7)、(27.0±0.9)和(55.0±3.6)cm。在生长旺盛的7—8月晴朗清晨,人工爬树截取样树树冠南向中上部枝条后,立即放入水桶,于水下环剥去除切口处3 cm左右的韧皮部并剪去大部分叶片,用黑色塑料袋罩住带回实验室,以保持离体枝条上叶片水分供应和光合活力[30-33]。
1.3 气体交换参数测定
选择枝条上叶位一致的健康成熟叶片,于9:00—11:30采用Li-6400XT光合仪(Licor,USA)的开放式气路,控制叶室内温度26 ℃左右,相对湿度60%左右,利用小钢瓶控制CO2含量为400 μmol/mol,应用仪器自带的红蓝光光源控制光照强度为1 500 μmol/(m2·s),分别测定3个树种雌雄植株的气体交换参数,包括:净光合速率[Pn,μmol/(m2·s)]、气孔导度[Gs,mol/(m2·s)]、胞间二氧化碳浓度(Ci,μmol/mol)和蒸腾速率[Tr,mmol/(m2·s)],并计算水分利用效率(EWUE,μmol/mmol),EWUE=Pn/Tr。对于不能充满2 cm×3 cm标准叶室的东北红豆杉针叶,每次完成光合测定后,剪下叶片,利用Photoshop软件确定其叶面积,并重新计算各参数。
在饱和光下充分诱导后,控制CO2含量为400 μmol/mol,光合有效辐射(PAR)梯度设置为1 600、1 400、1 200、1 000、800、600、400、300、200、150、100、50、0 μmol/(m2·s),测定光响应曲线。利用直角双曲线修正模型得到最大净光合速率[Pn,max,μmol/(m2·s)]、暗呼吸速率[Rd,μmol/(m2·s)]、光饱和点[PLSP,μmol/(m2·s)]和光补偿点[PLCP,μmol/(m2·s)],计算表观量子效率(ηAQY)[34-35]。
在各树种饱和光强下,控制CO2含量依次为400、250、200、100、50、150、400、800、1 000、1 200、1 800 μmol/mol,测定CO2响应曲线。利用直角双曲线修正模型得到潜在光合能力[Amax,μmol/(m2·s)]、CO2补偿点(CCP,μmol/mol)和光呼吸速率[Rp,μmol/(m2·s)],计算羧化效率[CE,mol/(m2·s)][34],利用FvCB模型得到最大羧化速率[CE,max,μmol/(m2·s)]和最大电子传递速率[RET,max,μmol/(m2·s)][36-37]。由于3个树种Ci值为CCP的2.4~4.6倍,推测CO2不是其光合过程的限制因素,因此本研究不关注Ci和CCP两个指标。
1.4 叶性状参数测定
将光合测定的叶片及其附近的成熟叶片取下,通过扫描仪(LiDE120,Canon,中国)确定叶面积后,泡水至饱和,称量叶片饱和鲜质量,随后在65 ℃烘干至质量恒定。利用Photoshop软件确定叶面积(leaf area,LA,cm2),计算比叶重(leaf mass area,LMA,g/m2),叶干物质含量(leaf dry mass content,LDMC)。采用AQ400全自动间断化学分析仪(SEAL Analytical,USA)测定叶氮质量分数,通过比叶重换算单位面积叶片氮含量(LNC,g/m2)。
1.5 数据处理
运用Microsoft Excel 2007和SPSS 20.0进行数据分析。采用单因素方差分析(One-way ANOVA)检验3个树种性别间叶性状参数和光合参数的差异,并用最小显著差异法(LSD)进行多重比较,显著水平均为0.05。利用SigmaPlot 12.5作图。图表中数据均为平均值±标准误。
2 结果与分析
2.1 3个树种雌雄株叶性状参数特征
同一树种雌株叶性状参数较雄株相应性状指标相差-5.5%~20.6%(P>0.05)(表1)。不同树种性别间叶性状指标差异显著(P<0.05)。LA从高到低依次为水曲柳、山杨、东北红豆杉(P<0.05)。东北红豆杉LMA和LNC显著高于山杨和水曲柳(P<0.05),且后两者间差异不显著。山杨LDMC显著高于水曲柳和东北红豆杉(P<0.05),且后两者间差异不显著。
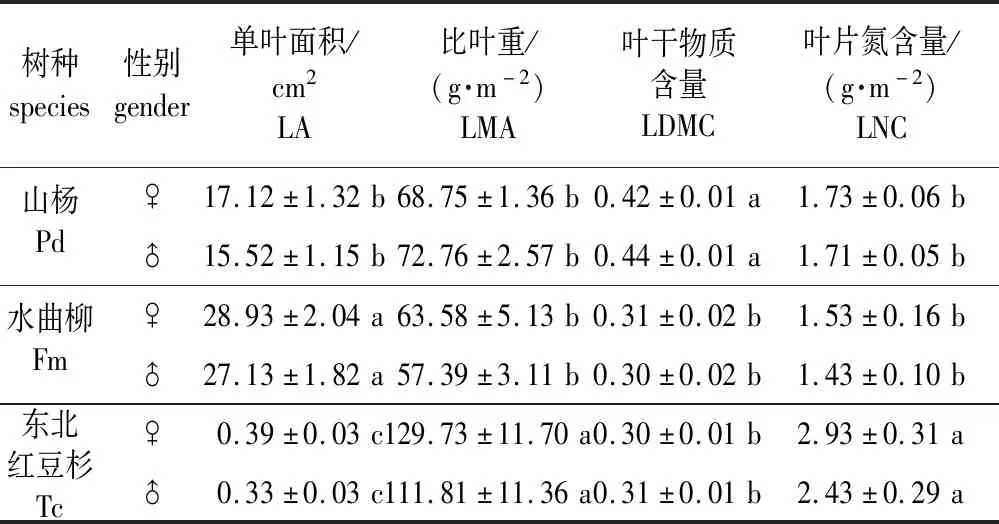
表1 3个树种雌雄株叶性状参数Table 1 Leaf traits parameters of three female and male tree species
2.2 3个树种雌雄株气体交换参数比较
同一树种雌雄株光响应曲线并不相同(图1A)。PAR在0~400 μmol/(m2·s)范围内,山杨雌株Pn略低于雄株;当PAR大于400 μmol/(m2·s)后,雌株Pn高于雄株。水曲柳雌雄株Pn在不同光强下均相似。而东北红豆杉雌株和雄株Pn在0~200 μmol/(m2·s)范围内相似;当PAR超过200 μmol/(m2·s)后,雌株Pn略低于雄株。在饱和光强下,同一树种雌雄植株Pn随着Ci升高而增加,在同一Ci浓度下,3个树种雌株Pn均高于相应的雄株(图1B)。通过描绘CO2含量低于200 μmol/mol的Pn与Ci散点图可见(图1C),3个树种雌株斜率均大于相应的雄株。3个树种Pn对光和Ci浓度的响应顺序由高到低均为水曲柳、山杨、东北红豆杉。
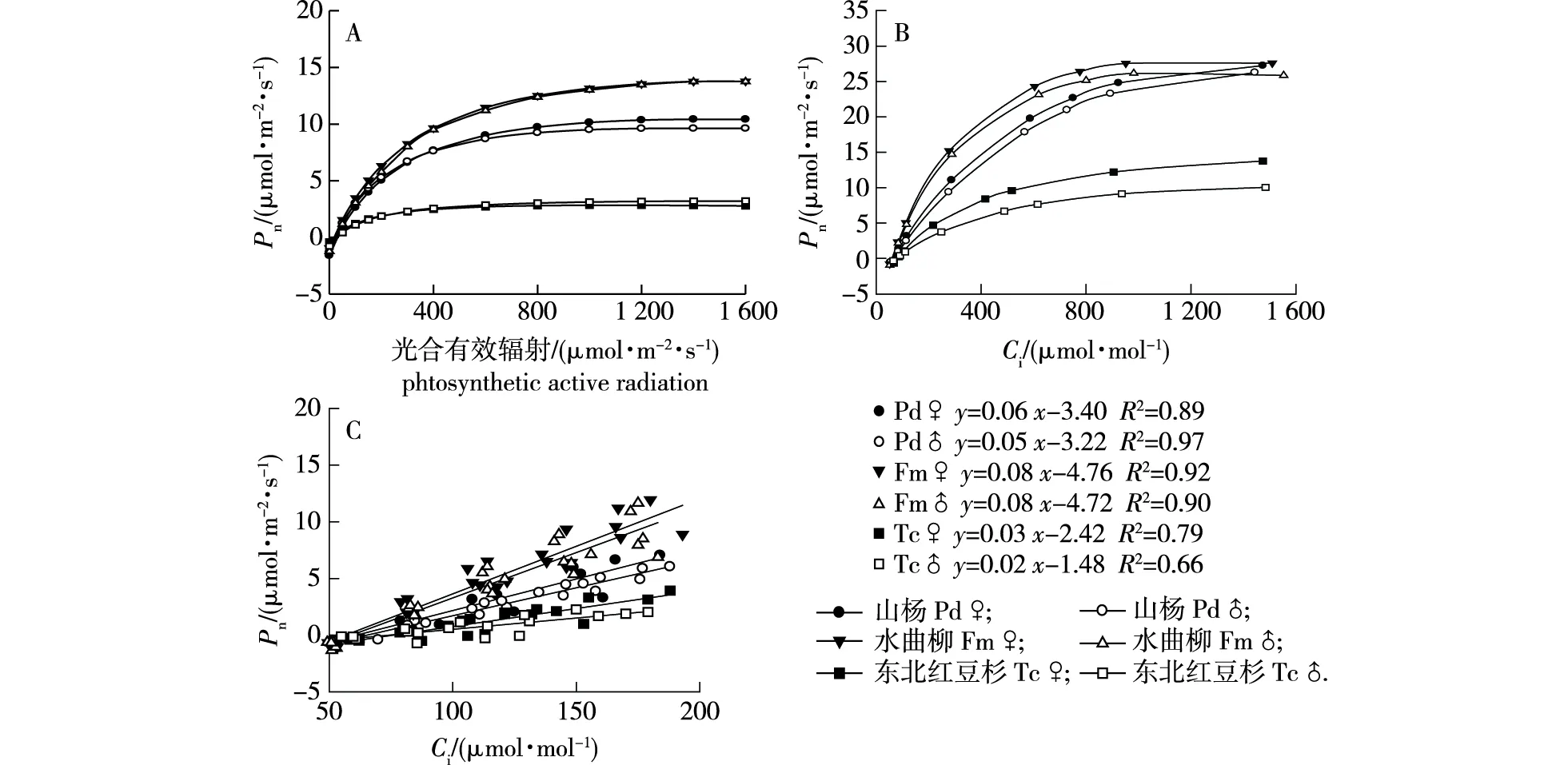
图1 3个树种雌雄株光、CO2响应曲线和Pn对低Ci的响应Fig.1 Light (A),CO2 (B) response curves and Pn response to low Ci concentration (C) of three female and male tree species
同一树种雌雄株间仅有几个气体交换参数差异显著(表2)。其中雌株山杨PLCP和CE,雌株水曲柳EWUE,雌株东北红豆杉CE和RP分别高于雄株32.3%、18.2%、26.0%、60.0%和59.7%,而雌株东北红豆杉PLSP和PLCP较雄株低24.1%和41.7%(P<0.05)。其余参数雌雄植株相差-48.1%~49.3%(P>0.05)。除Pn、Pn,max、Amax、Gs、Tr、ηAQY、CE,max和RET,max外,同一树种其余6个雌雄株参数改变了树种间比较结果。其中EWUE、Rd、PLCP影响3个种间比较结果;PLSP、CE和Rp影响山杨和东北红豆杉种间比较结果。

表2 3个树种雌雄株气体交换参数Table 2 Gas exchange parameters of three female and male tree species
3 讨 论
雌雄异株树木由于生殖成本的不同必然在光合生理生态方面有所体现。山杨、水曲柳和东北红豆杉雌雄植株间虽然仅有少数气体交换参数达到差异显著水平(P<0.05),但雌株光合特性总体优于雄株;另外雌株叶性状参数并不低于雄株。这可能与雌株存在着获取资源的补偿机制有关,雌株通过这种生存策略同时满足营养生长和生殖生长的需求。
叶片气体交换参数能够判定不同性别树木对光、CO2的利用以及对生长环境的适应能力。以往雌雄异株树木光合特性的研究多集中在温室幼苗,而且在光温水适宜条件下研究结果并不一致[3-8,38]。例如同为青杨(P.cathayana),Zhang等[7]认为雌雄株Pn、Gs、Tr和EWUE相差不足5.0%(P>0.05);而Liu等[6]表明该种雌株Pn、Gs和Tr低于雄株21.6%~38.8%(P<0.05),且两者EWUE相同(P>0.05);Liao等[3]却发现该种雌株Pn、EWUE、Rd和PLCP较雄株高27.0%~52.5%(P<0.05)。与温室扦插苗相比,野外自然生境成年雌雄植株关键光合特性参数对比结果均为雌株不同程度地优于雄株[10-11,13-14,16-17]。例如处于河岸带、雄雌比为0.51的连香树雌株Gs、Tr和PLCP较雄株低20.8%~56.1%(P<0.05);前者Pn,max、EWUE、ηAQY和CE,max较后者高20.1%~158.7%(P<0.05)[16]。本研究野外成年树种也表现为雌株以不同方式达到较高的光合生产和水分利用效率。相对于雄株,山杨和东北红豆杉雌株分别仅对强光和弱光适应能力强;而水曲柳雌株对强光和弱光均可充分利用。虽然3个树种雌株对光照条件的适应范围有所不同,但对光能吸收转化利用能力与雄株相似,保证了较强的同化力,同时具有较高的CE、CE,max和RET,max,提高了1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)活性、1,5-二磷酸核酮糖(RuBP)再生能力和Rubisco催化的最大反应速率,促进碳反应,通过少量水分消耗达到有机物生产[39-40]。以上3个树种雌株较高的RP值可能是通过耗散过剩光能,对光合机构进行保护的重要途径[41]。
由于雌株繁殖投入较高,它们会在其他生活史特征上表现出权衡,如雌株叶片在营养或繁殖生长期增强营养器官的生长和光合作用,获得更多的可利用资源,这样既满足了繁殖的资源需求,又没有减弱营养生长的资源投入,同时储存提供先花后叶树种例如水曲柳和山杨翌年开花需要的物质。这种较强的光合能力是雌株高繁殖压力下进化的一种额外补偿机制[14,42-43]。本研究山杨、水曲柳和东北红豆杉雌株的叶片生物量投入、捕获光能和获取资源能力并不低于雄株也证实了以上论点。另外,山杨雌株花序和水曲柳种子等繁殖器官是碳同化贡献重要的非叶光合器官[44],一方面制造有机养分,来有效弥补繁殖成本的消耗;另一方面,正是由于开花结实,起到增强碳汇强度的作用,反而加速了叶枝花果实等的碳同化速率。雌株这种稳健的适应性保证了整个种群的繁衍和发展。3个树种雌雄株仅有几个光合指标差异显著的原因可能与帽儿山地区土壤肥沃,生长季水热同步,生长环境优越,树木未受到水分、养分等因素胁迫有关;或与山杨、水曲柳和东北红豆杉种子小而多,繁殖成本较低有关。
3个树种的雌和雄株光合能力顺序均为水曲柳>山杨>东北红豆杉。与落叶阔叶树种山杨高生产、高消耗和水曲柳高生产、低消耗的光合特征相比,常绿针叶树种东北红豆杉虽然充分利用光合时间开始早、结束晚,即延长光合时间积累有机物用于生长与繁殖,但较低的光能利用效率、羧化效率和RUBP再生速率呈现明显的低生产、低消耗。导致该种繁殖更新难,或幼苗数量虽多,但生长缓慢,难以维持进入幼树阶段,从而形成目前不合理的种群年龄结构[45],这可能是东北红豆杉处于濒危状态的直接原因。
通过分析东北东部山区野外自然生长的雌雄异株成树山杨、水曲柳和东北红豆杉的光合以及叶性状指标,结果表明,虽然3个树种雌雄株仅有少数光合特性参数达到差异显著水平(P<0.05),但3个树种雌株整体表现出相对较高的光能、CO2和水分利用能力及光合生产力。同时雌株叶性状指标并不低于雄株。雌株具有一定的补偿生长,获得更多的可利用资源用于营养生长和生殖生长。3个树种中,东北红豆杉光合能显著低于山杨和水曲柳(P<0.05),这可能是东北红豆杉处于濒危状态的直接原因。在野外条件允许情况下,调整种群时应尽量考虑3个树种生理生态指标的性别因素。
致谢:黑龙江帽儿山森林生态系统国家野外科学观测研究站提供支持和帮助。
