转基因小黑杨根际土壤微生物群落特征研究
2023-03-30顾宸瑞杨蕴力
王 阳,王 伟,姜 静,顾宸瑞,杨蕴力
(林木遗传育种国家重点实验室(东北林业大学),黑龙江 哈尔滨 150040)
我国杨树基因工程始于20世纪80年代末,继选育获得首例抗虫转基因欧洲黑杨(Populusnigra)后[1],抗虫转基因毛白杨(P.tomentosa)、美洲黑杨(P.deltoides)、小黑杨(P.simonii×P.nigra)等陆续培育成功[2]。此外,杨树抗旱、耐盐或抗病等基因工程育种研究也取得了一定的成效,相继获得了不同转基因株系。这些转基因杨中仅有转抗虫欧洲黑杨获批商品化[3],大多数转基因杨树还处于中间试验或环境释放试验阶段,该阶段的主要研究内容之一就是进行转基因杨的环境生态安全性评价[4-8]。其中,转基因杨树对根际微生物数量及群落组成产生的影响愈发引起研究者们的重视。
转基因植物在生长发育过程中通过根系与土壤进行着频繁的物质交换,作为土壤生态系统重要组成部分的土壤微生物,尤其是根际微生物则极有可能受到影响。前人对其他转基因植物的研究发现部分转基因植物会通过根系残茬或分泌物对周围土壤产生一定影响,如抑制土壤中某些微生物生长的同时促进另一些微生物的生存与繁殖,致使土壤中营养成分发生改变与转化,从而使土壤最适宜于植物的生长[9];在关于磷高效转基因水稻(Oryzasativa)连续种植对土壤微生物影响的研究中发现,土壤微生物多样性确实受到了影响,但这种影响主要因施肥条件和植株发育时期的不同而变化[10]。另一方面,一些研究也发现转Bt基因玉米(Zeamays)和棉花(Gossypiumhirsutum)与非转基因植株相比,土壤微生物多样性及其活性没有显著差异[11-12]。
对转基因杨树根际微生物研究显示,转抗虫基因欧洲黑杨、转Bt基因‘南林895’杨(Populusdeltoides×P.euramericana‘Nanlin 895’)、转DREB毛白杨、转基因库安托杨(Populus×euramericana‘Guariento’)等杨树在外源基因导入后对根际细菌或真菌群落组成几乎没有影响或影响非常有限[13-19]。例如,转PeTLP基因‘南林 895’杨根际土壤中的细菌、真菌及放线菌数量在不同月份的变化趋势基本一致,且不同微生物的数量在转基因株系与非转基因株系之间的变化趋势也基本保持一致,转基因杨树与非转基因杨树之间土壤微生物的菌落数量间无显著性差异[20];7年生转Bt基因欧洲黑杨林地和土壤微生物数量调查显示,转基因植株根系与非转基因植株根系的土壤细菌、放线菌和霉菌数量无显著差异,转基因林地与非转基因林地间的土壤细菌、放线菌和霉菌数量也无显著差异[6]。
小黑杨(Populussimonii×P.nigra)是黑龙江省的主要人工造林树种之一,为了扩大其种植范围,提高在盐碱及干旱地区造林成活率,东北林业大学林木遗传育种国家重点实验室姜静教授研究团队自2001年开始,采用基因工程育种技术先后获得了转betA基因[21]、转LEA基因[22]及转WRKY70基因的小黑杨[3]。研究显示,betA基因及LEA基因在植物基因组中过表达后,能够显著提高植物甜菜碱、晚期胚胎蛋白的表达量,进而通过渗透调节提高植物的抗旱耐盐性[23-25]。WRKY70基因属于WRKY转录因子超家族第Ⅲ类亚家族成员,在植物的生物及非生物胁迫中均起着至关重要的作用,PsnWRKY70转录因子在盐胁迫应答信号转导网络中具有负调控作用,PsnWRKY70干扰表达小黑杨其耐盐性显著高于对照株系[3]。目前,通过转基因小黑杨的中间试验初步选择获得了生长适应性好的优良株系,是后续推广应用的首选转基因株系[26-28]。为了解上述转基因小黑杨对土壤微生物产生的影响,试验以转TaLEA、betA、WRKY70基因小黑杨根际土壤为试材,分析其根际土壤细菌及真菌群落组成特征,明确转基因活动对小黑杨根际群落微生物的影响,为后续的推广应用和产业化奠定基础。
1 材料与方法
1.1 试验地概况
研究地点位于黑龙江省哈尔滨市东北林业大学育种基地(126°64′E,45°72′N),属中温带大陆性季风气候,全年平均气温5.6 ℃,最热月平均气温23.6 ℃,最冷月平均气温-15.8 ℃,平均年降水量569.1 mm,主要降水月份为6—9 月,无霜期168 d。
1.2 试验材料
试验所用材料分别为:转TaLEA、转betA、转WRKY70-RNAi(图表中相应简称为TaLEA、betA、WRKY70)小黑杨及野生型(WT)小黑杨根际土壤。
2018年4月初分别采集参试小黑杨当年生枝条,剪切长度为12 cm左右的插穗,于温室中进行扦插,基质为V草炭土∶V河沙∶V黑土= 2∶2∶1,5月中旬待插穗生根、展叶后,每个株系选择生长一致的30株小苗,移栽于30 cm(高)×36 cm(径)花盆中,基质为V草炭土∶V河沙∶V黑土=2∶1∶1,每个花盆中加入等量基质,盆下放置塑料托盘,置于东北林业大学育种基地进行常规管理。
2020年6月中旬,选取株高为(190±5)cm的健康株系,各转基因小黑杨及野生型小黑杨分别选3株,共计12株苗木。分别采集植株根系,抖落较松散的土壤,将每种转基因小黑杨及野生型小黑杨的3株植株的根系分别进行混样,随后置于装有质量分数0.85% NaCL无菌溶液的三角瓶中,200 r/min震荡15 min,取出根系得到土壤悬浊液,将每组土壤悬浊液3等分,即为3次重复。每次取悬浊液于2 mL离心管中,4 ℃条件下10 000 r/min离心10 min,弃上清,反复5~6次进行悬浊液离心,将获得的土壤放入-80 ℃冰箱保存备用。采用QIAGEN公司的DNeasy PowerSoil Kit试剂盒提取土壤微生物总DNA。
将得到的DNA样品送至上海派森诺生物科技有限公司进行扩增,根际细菌扩增区域为V3—V4区,采用正向引物(5′-ACTCCTACGGGAGGCAGCA-3′)和反向引物(5′-GGACTACHVGGGTWTCTAAT-3′),扩增片段大小为500 bp;根际真菌扩增区域为ITS1区,采用正向引物(5′-GGAAGTAAAAGTCGTAACAAGG-3′)和反向引物(5′-GCTGCGTTCTTCATCGATGC-3′),扩增片段大小为280 bp。25μL的扩增体系中加入:5×反应缓冲液5 μL,5×GC buffer 5 μL,2 μL 2.5 mmol/L的 dNTP,1 μL 10 μmol/L的正向引物,1 μL 10 μmol/L的反向引物,2 μL DNA模板,8.75 μL ddH2O,Q5 DNA聚合酶0.25 μL。扩增参数为98 ℃ 2 min;98 ℃ 15 s,55 ℃ 30 s,72 ℃ 30 s,25~30个循环;72 ℃ 5 min。扩增完成后进行细菌16SrRNA、真菌ITS建库及高通量测序。
1.3 数据处理
用DADA2法[29]进行去引物,质量过滤,去噪(denoise),拼接和去嵌合体等。此法不再以相似度聚类,只进行去重(dereplication),即相当于以100%相似度聚类。质控后产生的每个去重序列称为ASVs (amplicon sequence variants,即特征序列)。用QIIME2 (2019.4)进行Alpha多样性、物种组成及差异分析。
用Excel、Origin 9.0、Heml等软件进行数据处理和作图。
2 结果与分析
2.1 土壤细菌及真菌样品测序深度评估
根据转不同基因小黑杨及野生型小黑杨的根际土壤细菌及真菌稀释曲线(图1),可知随着测序深度的增加,细菌及真菌稀释曲线逐渐趋于平缓,可观察的ASVs变化不大,说明测序数据量合理,能够真实反映土壤细菌及真菌的群落组成。
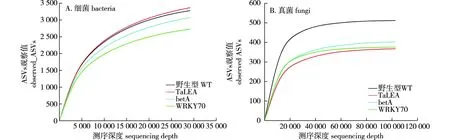
图1 参试小黑杨根际土壤细菌及真菌稀释曲线Fig.1 Rare faction curve of bacteria and fungi in rhizosphere soil of Populus simonii × P. nigra
2.2 根际土壤细菌、真菌群落丰度和Alpha多样性变化
土壤样品微生物群落Alpha多样性指数如表1所示。覆盖度(coverage)为测序深度指数,数值越高则说明样品中序列被测出的概率越高,参试样品细菌及真菌的覆盖度均在97%以上,表明绝大部分细菌及真菌种群被测出,测序结果可以反映根际土壤细菌及真菌群落多样性变化(表1)。
观察到的物种数(observed species)和Chao1指数是估算细菌、真菌群落丰度的参数,而Shannon和Simpson指数是估算群落多样性的参数。由表1可知,参试的转TaLEA、转betA、转WRKY70小黑杨及WT小黑杨,根际土壤细菌及真菌群落丰度、多样性不尽相同。在细菌群落丰度方面,转TaLEA、转betA小黑杨与WT小黑杨根际土壤细菌群落丰度无显著差异,而转WRKY70小黑杨显著低于WT小黑杨根际土壤细菌群落丰度(P<0.05);在细菌群落多样性方面,转TaLEA小黑杨与WT小黑杨差异不显著,而转betA及转WRKY70小黑杨均显著低于WT小黑杨(P<0.05)。

表1 参试小黑杨根际土壤细菌及真菌Alpha 多样性分析Table 1 Alpha diversity of bacteria and fungi in rhizosphere soil of Populus simonii × P.nigra
根际真菌群落丰度及多样性分析显示,3种转基因小黑杨的真菌群落丰度及多样性均显著低于WT小黑杨(P<0.05)。
2.3 根际土壤细菌及真菌群落结构差异性分析
基于参试小黑杨根际土壤细菌和真菌的ASV(特征序列)分别绘制了韦恩图(图2)。由图2可知,在根际土壤细菌方面,WT、TaLEA、betA和WRKY70 4组样品中特有的ASV数量分别为4 823、4 985、4 027和4 148个,各占总数的21.9%、22.6%、18.2%和18.8%。其中参试小黑杨共有的ASV有824个,占总数的3.7%,说明各组样品间ASV存在差异。根据与WT小黑杨共有ASV数量分析显示,与WT小黑杨根际土壤细菌群落相似程度最高的是转TaLEA小黑杨(1 996个),其次是转betA小黑杨(1 836个),相似度最小的是转WRKY70小黑杨(1 344个)。
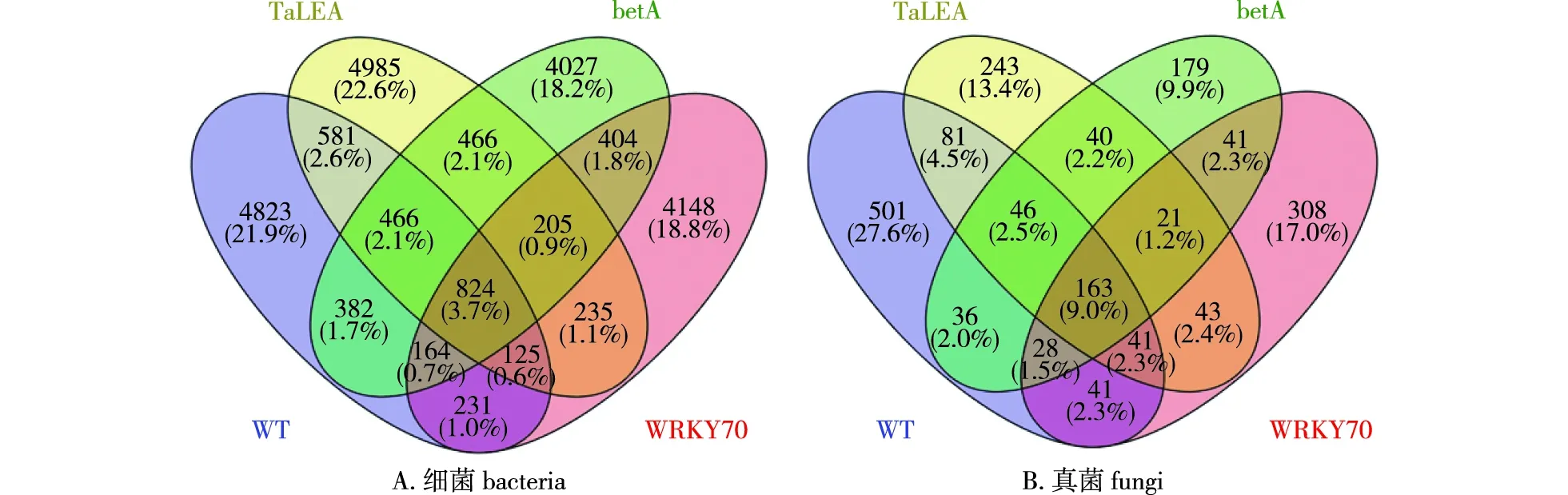
图2 参试小黑杨根际土壤细菌及真菌的ASV韦恩图Fig.2 ASV Venn diagram of bacteria and fungi in rhizosphere soil of Populus simonii × P. nigra
在根际土壤真菌方面,WT、转TaLEA、betA和WRKY704组样品中特有的ASV分别为501、243、179、308个,分别占总数的27.6%、13.4%、9.9%、17.0%。参试小黑杨共有的ASV有163个,占总数的9.0%,同样说明各组样品间ASV存在差异。根据与WT小黑杨共有ASV数量分析表明,与WT小黑杨根际土壤真菌群落相似度最高的是转TaLEA小黑杨(331个),其次是转betA小黑杨(273个)及转WRKY70小黑杨(273个)。
为了进一步比较样本间的根际土壤细菌及真菌属水平组成差异,用平均丰度位于前50属的丰度数据绘制了聚类热图(图3),结果显示,在根际土壤细菌及真菌群落组成上,转TaLEA小黑杨与WT小黑杨更接近,转betA小黑杨与转WRKY70小黑杨更接近,这也与PCA(图4)结果相符。

A:Gemmatimonas.芽单胞菌属;Rhodanobacter.产黄杆菌属;Lactobacillus.乳(酸)杆菌属;Sphingomonas.鞘氨醇单胞菌属;Pseudomonas.假单胞菌属;Arenimonas.砂单胞菌属;Novosphingobium.新鞘氨醇杆菌属;Flavobacterium.黄杆菌属;Phenylobacterium.苯基杆菌属;Hyphomicrobium.生丝微菌属;Pseudolabrys.假双头斧形菌属;Amycolatopsis.拟无枝酸菌属;Devosia.德沃斯氏菌属;Bradyrhizobium.慢生根瘤菌属;Nocardioides.类诺卡氏属。测序结果未能将Subgroup_7、Burkholderia-Caballeronia-Paraburkholderia、Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium、Subgroup_6、Subgroup_2准确归类。Saccharimonadales、Ellin6067、SC-I-84、Occallatibacter、Bryobacter、Candidatus_Solibacter、KD4-96、Ramlibacter、IMCC26256、MND1、IS-44、Haliangium、Dokdonella、Mucilaginibacter、Rhizobacter、Granulicella、GOUTA6、Bauldia、Dongia、Jatrophihabitans、67-14、Reyranella、Acidibacter、Candidatus_Koribacter、A4b、SWB02、WPS-2、Micropepsis、Chloroplast、Rhodovastum尚未确定中文名称。B:Tomentella.棉革菌属;Clavulina.锁瑚菌属;Neurospora.链孢霉属;Pseudogymnoascus.假裸囊菌属;Mrakia.木拉克酵母属;Coniochaeta.锥毛壳属;Penicillium.青霉属;Byssochlamys.丝衣霉属;Talaromyces.踝节霉属;Paecilomyces.拟青霉属;Fusarium.镰刀菌属;Trichoderma.木霉属;Pseudeurotium.假散囊菌属;Plenodomus.丰屋菌属;Inocybe.丝盖伞属;Hebeloma.粘滑菇属;Sphaerosporella.小球孢盘菌属;Geopora.地孔菌属;Laccaria.蜡蘑属;Didymella.亚隔孢壳属;Cadophora.背芽突霉属;Thanatephorus.亡革菌属;Aureobasidium.短梗霉属;Galerina.盔孢伞属;Plectosphaerella.小不整球壳属;Mycosphaerella.球腔菌属;Filobasidium.线黑粉菌属;Mortierella.被孢霉属;Epicoccum.附球霉属;Paraglomus.类球囊霉属;Leptosphaeria.小球腔菌属;Rhodotorula.红酵母属;Aspergillus.曲霉属;Peziza.盘菌属;Gibberella.赤霉属;Alternaria.链格孢霉属;Septoria.壳针孢属;Olpidium.油壶菌属。Serendipita、Capronia、Protostropharia、Fibulochlamys、Archaeorhizomyces、Paraphaeosphaeria、Cystofilobasidium、Ceratobasidium、Echria、Naganishia、Tausonia、Neoascochyta尚未确定中文名称。Z值为该样品在属分类水平的相对丰度与所有样品在该水平的平均相对丰度的差除以所有样品在该水平上的标准差所得到的值。Z value represents the data obtained by dividing the difference between the relative abundance of the sample at the genera classification level and the average relative abundance of all samples at that level by the standard deviation of all samples at that level.图3 参试小黑杨根际土壤细菌及真菌群落属水平排序前50的丰度聚类热图Fig.3 Cluster heat map at genus level(top 50) of abundance of bacteria and fungi in rhizosphere soil of Populus simonii × P.nigra
主成分分析(PCA)显示,PC1和PC2对根际土壤细菌群落结构的贡献率分别为54.1%和24.8%(图4A),累积贡献率为78.9%,认为是造成细菌群落结构变化的主要来源;同时在根际土壤真菌群落PCA分析中,PC1和PC2的贡献率分别为63.9%和21.6%(图4B),累积贡献率为85.5%,说明这两个主成分可以解释真菌群落差异。结合图2可知,无论是根际土壤细菌群落结构还是真菌群落结构,转TaLEA小黑杨与WT小黑杨的群落差异较小,而转WRKY70小黑杨与WT小黑杨的群落差异相对较大。
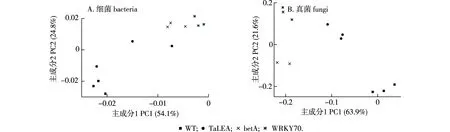
图4 参试小黑杨根际土壤细菌及真菌群落属水平主成分分析Fig.4 PCA(principal component analysis) at genus level of bacteria and fungi communities in rhizosphere soil of Populus simonii × P.nigra
2.4 根际土壤细菌及真菌群落组成分析
小黑杨根际土壤样品细菌群落结构在门水平和属水平排序前20的相对丰度见图5、图6。
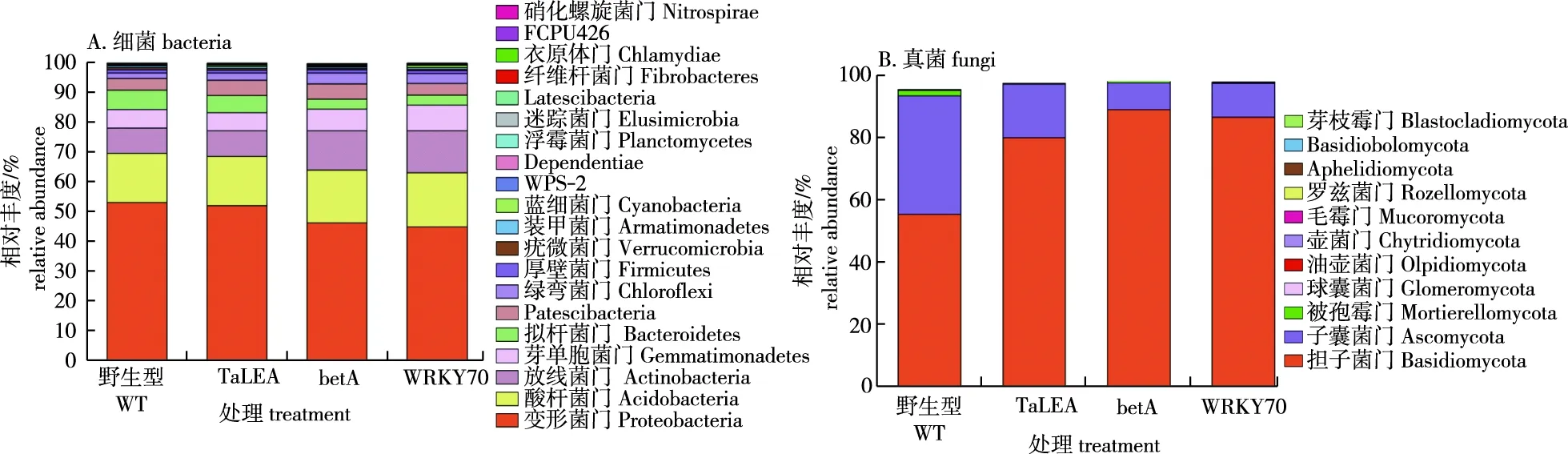
图5 参试小黑杨根际土壤细菌、真菌群落门分类水平排序前20的相对丰度Fig.5 Relative abundance at phylum level(top 20) of bacteria and fungi in rhizosphere soil of Populus simonii × P. nigra

图6 参试小黑杨根际土壤细菌、真菌群落属分类水平排序前20的相对丰度Fig.6 Relative abundance at genus level(top 20) of bacteria and fungi in rhizosphere soil of Populus simonii × P. nigra
由图5可知,门水平上主要包括由变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、放线菌门(Actinobacteria)、芽单胞菌门(Gemmatimonadetes)、拟杆菌门(Bacteroidetes)、Patescibacteria和绿弯菌门(Chloroflexi)(图5A),占细菌总数的 96.37%~96.54%,其中变形菌门为优势菌门,占细菌总数的44.78%~53.00%。在4个参试样品中,WT与TaLEA根际细菌群落结构在门水平更接近,betA与WRKY70更接近。真菌群落结构在门水平上则是担子菌门(Basidiomycota)和子囊菌门(Ascomycota)为优势菌门(图5B),占真菌总数的93.46%~97.64%,其中,转TaLEA、转betA和转WRKY70小黑杨根际担子菌门相对丰度显著高于WT株系。
小黑杨根际土壤样品细菌群落结构在属水平上的组成和丰度如图6A所示。参试样品土壤细菌群落前20占细菌总量的33.13%~38.74%,由图6可知参试样品在组成上比较接近,同时在几个属的丰度上存在差异。与WT小黑杨比较,在3种转基因小黑杨根际土壤中假双头斧形菌属(Pseudolabrys)、产黄杆菌属(Rhodanobacter)及德沃斯氏菌属(Devosia)丰度均有不同程度的提高,而鞘氨醇单胞菌属(Sphingomonas)丰度则呈现不同程度的降低;在转betA小黑杨和转WRKY70小黑杨中,芽单胞菌属(Gemmatimonas)和Ramlibacter丰度有不同程度的提高;Occallatibacter仅在转TaLEA小黑杨中丰度更高;同时Ellin6067丰度仅在betA小黑杨中升高。
根际土壤真菌群落结构在属水平上排名前20的物种相对丰度分析显示(图6B),参试样品真菌群落前20占真菌总量的82.21%~93.75%,不同小黑杨之间的真菌群落多样性和丰度存在差异。与WT小黑杨比较,3种转基因小黑杨根际土壤中Serendipita丰度均升高,亚隔孢壳属(Didymella)和背芽突霉属(Cadophora)丰度则降低了85%以上;转TaLEA小黑杨、转WRKY70小黑杨的棉革菌属(Tomentella)丰度降低,粘滑菇属(Hebeloma)则高于WT小黑杨;另外,转TaLEA及betA小黑杨根际土壤中的丝盖伞属(Inocybe)丰度有不同程度的提高,小球孢盘菌属(Sphaerosporella)和蜡蘑属(Laccaria)仅在TaLEA根际土壤中丰度较高。
3 讨 论
植物生长的土壤中存在大量微生物,主要包括细菌、真菌、放线菌和土壤原生动物。其中细菌及真菌对土壤生态系统的作用至关重要[30]。研究发现,根际土壤细菌及真菌群落组成非常敏感,受植物种类、植株生长时期、耕种管理方式及外源基因的导入等生物和非生物因素的影响[31]。为了解转TaLEA、转betA及转WRKY70小黑杨对土壤微生物群落组成产生的影响,本研究采用高通量测序技术分别对转TaLEA、转betA及转WRKY70小黑杨与野生型小黑杨的根际土壤细菌及真菌群落差异进行分析。结果发现,转betA及WRKY70小黑杨影响了根际土壤细菌及真菌群落多样性、群落丰度,低于或显著低于WT小黑杨,转TaLEA小黑杨在真菌群落丰富度和多样性方面显著低于WT小黑杨,表明外源基因的导入或过量表达对根际微生物群落产生一定影响。根据参试样品的韦恩图、主成分分析和聚类热图显示,转TaLEA小黑杨与WT小黑杨的群落组成非常相似,转betA小黑杨与转WRKY70小黑杨群落组成更接近,说明转betA及转WRKY70小黑杨的根际土壤细菌和真菌群落结构与WT小黑杨明显不同。前人研究发现,转基因植物可能对土壤微生物群落产生影响,但该影响远远小于作物的生育及生长季节、田间管理、肥料的施加等因素对土壤微生物产生的影响[32-33]。同样,Gyamfi等[34]对转抗除草剂基因油菜(Brassicanapus)的研究也证明,转基因油菜根际微生物群落较野生型发生了轻微变化,认为这种影响只是暂时的,与油菜发育阶段引起的微生物群落变化相比要小。对转cry1Ac基因和转cry1Ab基因水稻安全性分析显示,在生长发育的旺盛期转基因水稻对土壤细菌数量、多样性及功能多样性产生一定的影响,但这种影响并不持续,随着转基因水稻的生长发育直至成熟期而逐渐消失[35]。本实验的季节正是处于杨树生长最旺盛的季节,转betA、转WRKY70小黑杨的根际土壤细菌和真菌群落结构与WT小黑杨的差异能否随着生长发育而减弱,还需后续实验验证。
参试转基因小黑杨根际土壤细菌群落结构在属水平上的组成和丰度分析显示,与WT小黑杨比较,在3种转基因小黑杨根际土壤中假双头斧形菌属、产黄杆菌属及德沃斯氏菌属丰度均有不同程度的提高;Ellin6067丰度在转betA小黑杨土壤中明显升高,较其余样品均值高20.44%,而在转TaLEA小黑杨土壤中则较其余小黑杨样品的均值低19.83%;对于Occallatibacter丰度则仅在转TaLEA小黑杨土壤中显著提高,较其余样品均值提升31.52%。
研究已证明假双头斧形菌属是植物根际的有益细菌,能够促进植物对激素、生物碱以及氮的代谢等,进而促进植物的生长发育,提高植物抵抗病原菌的能力,可具有抑制土壤病原菌并促进植物生长、提升土壤酶活性及分解土壤中有害物质的作用,被称为“烃类降解剂”[36-38]。产黄杆菌属有能够降解氯化芳香族化合物及各种芳香族化合物的功能[39-40],Ellin6067为氨氧化细菌,参与分解利用土壤中几丁质的Occallatibacter、能够分解有机质提升土壤肥力的芽单胞菌属等土壤有益菌的生长情况也在不同转基因小黑杨中得到提升[41-43]。
真菌群落结构在属水平上的组成分析显示,致病真菌背芽突霉属(Cadophora)和亚隔孢壳属(Didymella)[44-45]在3种转基因小黑杨中均被明显抑制,同时一些有益的菌根真菌丰度明显高于WT小黑杨。这其中丝盖伞属丰度在转TaLEA、betA小黑杨中显著高于另外两个样品,分别为13.97%和15.44%;小球孢盘菌属、粘滑菇属、蜡蘑属丰度仅在转TaLEA小黑杨中显著提高,其丰度分别较其余样品均值提升了6倍、18倍和1倍。这些菌根真菌属的某些种已经被鉴定为杨树外生菌根真菌主要类群[46-50]。
本实验中所涉及的betA、TaLEA外源基因是两个广泛应用于植物基因工程的功能基因,其表达产物分别为甘氨酸甜菜碱及晚期胚胎富集蛋白,导入小黑杨基因组后能够不同程度的提高受体的耐盐性[8-9],WRKY70属于WRKY转录因子超家族第Ⅲ类亚家族成员,在植物的生物及非生物胁迫调控网络中起着关键作用,转WRKY70-RNAi小黑杨耐盐性明显提高[11]。上述转基因小黑杨在生长发育过程中可能通过根系分泌物或其他途径富集根际土壤中潜在益生菌及菌根真菌,这些细菌与真菌可能参与抑制有害微生物的生长,提高了转基因植株抗逆能力,进而增强了转基因小黑杨的抗非生物胁迫能力,这种推测还需后续试验进行证明。
实验虽然发现转TaLEA、转betA及转WRKY70小黑杨对根际土壤细菌及真菌多样性及丰度产生了一些影响,但这些影响主要表现为转基因小黑杨根际潜在益生菌的增加、致病菌的减少。笔者认为仅这一方面的变化并不足以评判其对环境而言安全与否,还需深入分析根际微生物群落结构是否合理、微生物间的是否具有拮抗作用,以及这种影响是否是因为本研究转基因杨树正处于生长季,或这些影响是否只是暂时性等。因此,后续还需对3种转基因小黑杨在不同发育时期及不同年份进行根际土壤微生物群落结构研究,进一步评价其对环境微生物群落组成的影响。
