6种竹子叶器官的解剖结构比较
2023-03-30金点坤王曙光王声翰
金点坤,吕 卓,王曙光,龙 昊,张 冲,王声翰
(西南林业大学生命科学学院,云南 昆明 650224)
竹类植物叶器官包括营养叶和茎生叶/秆箨,具有不同的生理功能[1]。竹子的营养叶主要包括叶片、叶柄和叶鞘,秆箨主要包括箨鞘和箨片两部分,叶柄则是竹类植物与其他草本植物的重要区别[2];秆箨是禾本科植物重要的形态特征,是竹类植物分类学上鉴定竹种的主要依据[2]。在竹类植物中,箨器官又分为笋箨/秆箨、鞭箨和叶箨[1-3]。Wong[1]研究发现,由于秆箨与营养叶的基本组成一致,箨鞘的上部都具有细长的叶,所以认为秆箨是叶子的另一种存在形式。秆箨通常为绿色或者其他颜色,有的能够在竹秆成熟后仍然宿存一段时间,具有明显的种间差异性[1-4]。
在竹子高生长的过程中,箨鞘包裹着幼嫩的节间,起到关键的机械支持作用[4]。近年来,箨鞘的生理功能受到越来越多的重视,Wang等[5-6]对云南箭竹快速生长进行研究,发现秆箨的横向切除会影响竹子的高生长,纵向切除会导致竹子的弯曲生长,认为秆箨是竹笋高生长过程中的水分调节器,证实了水分在竹子高生长中的重要作用;Chen等[7]研究发现竹子箨鞘衰老与糖分累积相关。
Wang[3]观察到秆箨没有主脉,但其维管束解剖结构与叶枝相似,表皮气孔密度和形态与叶枝更加接近,因此认为秆箨与枝同源,是枝的变态,但并没有比较其解剖结构与叶柄之间的差异,忽略了箨鞘与叶鞘基部均包裹着腋芽的现象。丁雨龙等[8]在植物形态学描述中,定义了完全叶是指具叶片、叶柄和托叶3个部分的叶,叶柄为连接叶片与枝的部分。此外,在其他植物中,也有学者提出叶鞘起源于叶柄,如伞形科植物叶的叶鞘都是由叶柄基部或全部扩大成鞘状形成[9]。传统的观点认为竹子的箨鞘是一种变态的叶片,但在叶子中两片叶之间不可能存在叶柄,而叶片的基部往往是叶柄,从这个角度看,箨鞘应该属于叶柄的变态。竹类植物中的箨鞘与箨片,以及叶片、叶柄、叶鞘与枝之间的形态与解剖结构仍然需要深入研究,以更好地解释叶鞘与箨鞘的起源问题。本研究选取6个云南常见竹种,以其营养叶与秆箨等为研究对象,对其解剖结构进行比较,分析不同叶器官间的解剖学与功能差异,探究不同叶器官在竹类植物生长发育过程中所扮演的角色及箨鞘的起源,为竹类植物基础生物学研究提供新的理论信息。
1 材料与方法
1.1 供试材料
选择6种云南常见竹种包括勃氏甜龙竹(Dendrocalamusbrandisii)、慈竹(Bambusaemeiensis)、绵竹(Lingnaniaintermedia)、香竹(Chimonocalamusdelicatus)、云南箭竹(Fargesiayunnanensis)、美竹(Phyllostachysmannii),以其营养叶、秆箨及叶枝器官为研究对象。所有试验材料均采自西南林业大学竹园内(102°45′40″E,25°3′41″N),研究地海拔1 936 m。
为减少采样位置导致的误差,试验样品均取自各竹种形态位置一致(即形态学中部位置)的30片/段新鲜且成熟的叶片、叶柄、叶鞘、秆上部箨片(地上部分10节以上,下同)、秆下部箨片(地上部分3节以下,下同)、秆上部箨鞘、秆下部箨鞘以及叶枝共8个部位,将采集的样品剪成约0.5 mm×0.5 mm(长×宽),放入FAA(体积分数50%乙醇、甲醛、乙酸体积比18∶1∶1)中固定抽气备用。
1.2 石蜡切片制作
将固定好的材料用梯度叔丁醇(从体积分数50%开始)脱水[10]、石蜡包埋。旋转切片机连续切片(厚7~8 μm),番红-固绿染色,加拿大树胶封片制成永久装片,放置在显微镜(凤凰牌PH100-3B41l-IPL)下观察,并用二维测量软件DS-3000拍摄测量。
1.3 指标测定及数据处理
记录各竹种叶片的叶宽、叶长和一级脉数量,并测量同一竹种叶片、叶柄、叶鞘、秆上部箨片、秆下部箨片、秆上部箨鞘、秆下部箨鞘以及叶枝的厚度、近轴面表皮厚度、远轴面表皮厚度、维管束面积、木质部面积(原生导管与后生导管总面积)、韧皮部面积和泡状细胞面积。每个部位测量30片/段,每片/段测量30次,如所测量的部位缺失则不进行测量。用Excel 2019和SPSS 22.0进行数据处理、单因素方差分析(One-way ANOVA)及Duncan多重比较。
2 结果与分析
2.1 6种竹子营养叶解剖结构比较
2.1.1 叶片形态
6种竹子间叶形差异较大(图1、表1)。勃氏甜龙竹的叶片长度与宽度均最大,分别为28.08 cm和4.67 cm,长宽比为6.01;其次为慈竹;香竹叶片长度与宽度均最小,分别为8.76 cm和1.23 cm,其长宽比为7.12,与绵竹近等宽。美竹的叶片厚度最大,为124.56 μm,其次为勃氏甜龙竹和香竹的叶片,云南箭竹叶片厚度最小,为81.19 μm。

A.勃氏甜龙竹Dendrocalamus brandisii;B.慈竹Bambusa emeiensis;C.绵竹Lingnania intermedia;D.香竹Chimonocalamus delicatus;E.云南箭竹Fargesia yunnanensis;F.美竹Phyllostachys mannii。下同。The same below.图1 6种竹子叶外部形态特征Fig.1 The external morphological characteristics of six bamboo species leaves
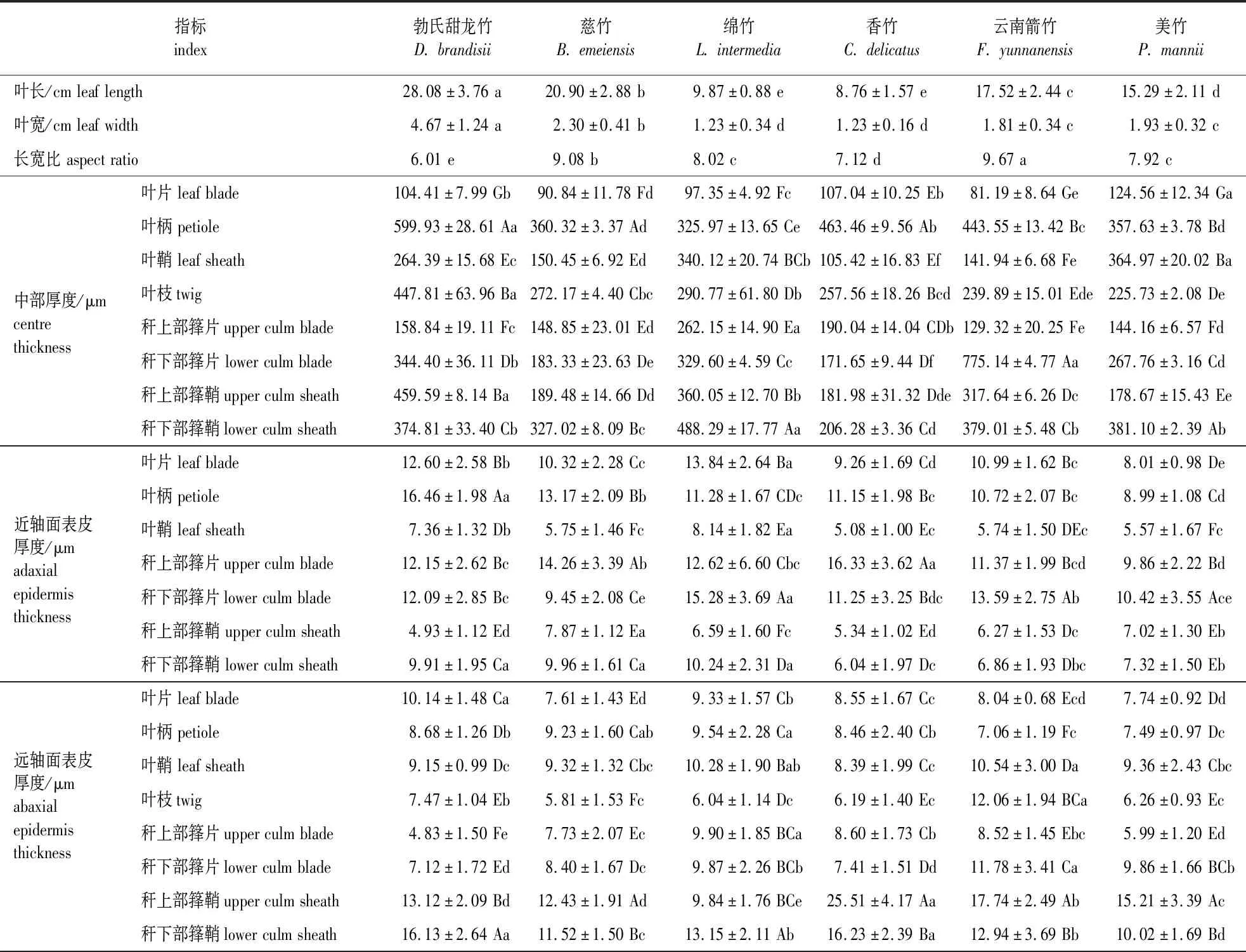
表1 6种竹子营养叶、叶鞘及枝条解剖特征Table 1 The anatomical characteristics of the leaves,sheath and branch of six bamboo species

表1(续)
6种竹子叶片均为长卵状披针形,体视显微镜下观察发现,叶片叶缘均具有叶尖朝向的透明硅质针刺,不同竹种叶片基部的形态和叶缘的针刺大小、形态及分布密度均不相同,叶缘的不同位置也有较大差别(图2)。总体而言,竹子叶片内缘的叶缘刺细小且数量少,刺尖均朝向叶尖;外缘部分的叶缘刺长而坚硬,分布数量多,叶缘触感更为粗糙。不同竹种叶片叶缘刺大小、数量和形态均有较大的差异,叶缘刺能否作为竹种分类依据,还需要进一步研究。

a.叶基leaf base;b.中部内缘central inner edge;c.中部外缘central outer edge;d.上部内缘upper inner edge;e.上部外缘upper outer edge.图2 6种竹子叶片外部形态特征Fig.2 The external morphological characteristics of six bamboo species leaves
2.1.2 叶片解剖结构
不同竹种叶片的解剖结构基本一致,均包括表皮、基本组织和维管束3个部分。表皮分为近轴面表皮和远轴面表皮,基本组织主要包括叶肉和梭形细胞,维管束分为一级平行脉和二级平行脉维管束。
1)叶片表皮。6种竹子种叶片的近轴面表皮均为单层细胞,厚度均大于远轴面,细胞透明近圆形,排列紧密,多为长圆形,外表面有高度角质化的圆锥状乳突(图3 Aa—Fa)。6个竹种中,近轴面表皮厚度最大的为绵竹,厚度为13.84 μm,其次为勃氏甜龙竹叶片,厚度为12.60 μm,厚度最小的为美竹,为8.01 μm(表1)。竹种叶片近轴面两个维管束之间分布着大小不一的泡状细胞组成扇形泡状细胞复合体,外壁不加厚(图3 Aa—Fa)。
不同竹种间泡状细胞复合体的大小、形状都存在差异。勃氏甜龙竹、绵竹与美竹的泡状细胞复合体的中部最大、两侧较小,整体呈扇形,由3~5个细胞组成(图3Aa、3Ca、3Fa)。慈竹与云南箭竹的泡状细胞复合体整体呈马蹄形,由2~3个细胞组成,但慈竹的泡状细胞复合体向外凸出,高于近轴面表皮(图3 Ba、3Ea)。香竹泡状细胞复合体整体下陷呈月牙形或马鞍形,由4~6个泡状细胞组成(图3 Da)。不同竹种叶片的远轴面表皮均为单层细胞,厚度小于近轴面表皮。不同竹种叶片远轴面表皮厚度也有差异,勃氏甜龙竹远轴面表皮厚度最大,为10.14 μm;慈竹最薄,为7.61 μm。
2)基本组织。竹类植物叶片基本组织以梭形细胞收缩后的空穴为界,将叶肉组织分为近轴面和远轴面两部分,靠近近轴面表皮的叶肉细胞呈指状,靠近远轴面表皮的叶肉细胞呈梅花状。梭形细胞收缩后的空穴对称地分布在维管束两端,且不超过泡状细胞所在的位置,故空穴大小受限于维管束与泡状细胞之间的距离。
梭形细胞将叶肉组织分为近轴面和远轴面两部分,不同竹种两部分的细胞层数也不同,其中勃氏甜龙竹叶片近轴面叶肉组织有2~3层细胞,远轴面仅有1层细胞(图3 Aa);慈竹近轴面2~3层细胞,远轴面有2层细胞(图3 Ba);绵竹与香竹、云南箭竹叶片表现一致,均表现出近轴面3~4层细胞、远轴面2层细胞(图3 Ca—Fa),近轴面叶肉组织更加发达。
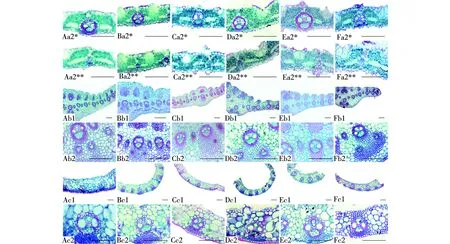
a.叶片leaf blad;b.叶柄petiole;c.叶鞘leaf sheath.1.100×;2.400×.*.大维管束macro-vascular bundle;**.小维管束small-vascular bundle.比例尺为100 μm。下同。Bar is 100 μm.The same below.图3 6种竹子营养叶解剖形态特征Fig.3 The anatomical and morphological characteristics of six bamboo species vegetative leaves
3)维管束。竹子叶片纵向平行脉维管束分为一级平行脉维管束和二级平行脉维管束(图3 Aa—Fa)。一级平行脉和二级平行脉维管束鞘细胞层数不同,一级平行脉维管束鞘细胞有2层,外层为薄壁鞘细胞,内层为厚壁鞘细胞。二级平行脉维管束鞘细胞仅为1层薄壁鞘细胞。各维管束鞘的近轴面和远轴面均由厚壁组织连接着近、远轴面表皮。维管束内部由初生木质部和初生韧皮部构成,木质部由1个原生导管和2个后生导管构成,分布在叶片近轴面,韧皮部分布在叶片远轴面。不同竹种维管束的面积有一定的差异,其中香竹叶片维管束面积最大,而绵竹叶片维管束面积最小。不同竹种维管束面积与叶片的形态也并不完全一致(表1)。不同竹种的叶片维管束,无论韧皮部面积还是韧皮部面积占比,均大于木质部。
2.1.3 叶柄
竹类植物的叶柄位于叶片与叶鞘之间,起到连接叶片与叶鞘的功能,是竹类植物与其他草本植物的重要区别之一。根据其横切面的解剖结构,将其分为表皮、基本组织和维管束3个部分。
1)叶柄表皮。竹类植物的叶柄形态略扁,并向远轴面突起,形态与叶片的主叶脉相似。叶柄表皮也可分为近轴面表皮与远轴面表皮。所有竹种叶柄的近轴面表皮均为单层细胞,表皮细胞厚度最大的为勃氏甜龙竹[(16.46±1.98)μm] ,最小的为美竹[(8.99±1.08)μm](表1)。表皮细胞为长圆形或近圆形,与其内部的基本组织薄壁细胞相比,形状较小、排列整齐紧密。此外,多数竹种叶柄近轴面表皮在维管束两侧位置发生显著下陷而呈波浪状凸起,而美竹叶柄整体较为平整。在近轴面表皮凸起部位的细胞,其近轴面细胞壁发生加厚并且角质化,下方具有发达密集的纤维鞘,下陷部位的细胞则均为薄壁细胞。所有竹种叶柄远轴面表皮均为单层细胞,细胞呈长圆形或近圆形,排列整齐,形态与近轴面表皮类似,细胞大小小于近轴面表皮。勃氏甜龙竹、慈竹、绵竹和云南箭竹远轴面表皮细胞相对平整,香竹与美竹远轴面有圆锥状的刺毛,长短不一(图3 Ab—Fb)。
2)基本组织与维管束。竹类植物叶柄的基本组织主要由薄壁细胞构成。与叶片显著差异的是其维管束呈上、下两行,平行排列于叶柄的基本组织中。靠近轴面的维管束木质部纤维鞘更发达,靠远轴面的维管束韧皮部纤维鞘更加发达。近轴面维管束的发育程度低于远轴面的维管束,且在远轴面的维管束呈现出一大一小间隔排列的方式。叶柄中部维管束形态与组成类似于叶片主脉,说明叶片的主脉维管束起源于该部位的维管束。叶柄中的大维管束发育完善,由初生导管、次生导管及韧皮部组成,而小维管束多发育不完善,仅由韧皮部和1个导管或无导管组成。在6种竹子的叶柄中,维管束鞘通常均为1层鞘细胞,少数为双层鞘细胞,鞘细胞壁发生显著加厚,不具有叶脉维管束的薄壁鞘细胞。从位置上看,叶柄维管束的韧皮部和木质部纤维鞘,相当于叶脉维管束近轴面与远轴面的厚壁组织,对叶柄起机械支持功能(图3 Ab—Fb)。
不同竹种叶柄的维管束面积也存在显著差异,最大的为香竹,其维管束面积为3 111.31 μm2,最小的为慈竹,只有1 670.84 μm2(表1)。除慈竹外,其他竹种的叶柄维管束面积均大于叶片维管束。与叶片的维管束相同,叶柄维管束的韧皮部均朝向远轴面,木质部均朝向近轴面,而且维管束中的韧皮部面积及其面积占比,均显著高于木质部(表1)。小维管束也几乎被韧皮部占满,或仅有1个导管,说明叶柄的有机物运输组织更为发达。
2.1.4 叶鞘
竹类植物的叶鞘着生在叶枝的节部,起到连接叶柄与叶枝的功能,叶鞘包裹着节芽和幼嫩的枝节。在横切面上,叶鞘也由表皮、基本组织和维管束组成,与叶片不同是,叶鞘不具有明显的主脉。不同竹种叶鞘厚度具有较大的差异,由大到小分别为美竹>绵竹>勃氏甜龙竹>慈竹>云南箭竹>香竹,其中美竹叶片最厚,叶鞘也最厚(表1)。
1)叶鞘表皮。叶鞘的近轴面表皮光滑,为单层细胞组成,排列整齐紧密,细胞壁加厚不显著,不具有泡状细胞和梭形细胞。美竹叶鞘近轴面表皮细胞形态为近圆形,其余竹种均为横向的长方形或长圆形,厚度小于远轴面表皮(图3 Ac—Fc)。不同竹种叶鞘近轴面表皮厚度存在差异,表现为绵竹叶鞘近轴面表皮最厚,为8.14 μm;其次为勃氏甜龙竹,最薄的为香竹;叶鞘近轴面表皮厚度仅为5.08 μm。叶鞘远轴面表皮为单层细胞,细胞均为长圆形,排列整齐。细胞远轴面细胞壁角质化加厚。不同竹种叶鞘远轴面表皮厚度也存在差异,具体表现为云南箭竹和绵竹叶鞘的远轴面表皮细胞最厚,分别为10.54 μm和10.28 μm;最薄的为香竹,远轴面表皮细胞厚度仅为8.39 μm(表1)。
2)基本组织与维管束。竹子叶鞘维管束分布在远轴面,维管束之间没有梭形细胞。基本组织靠近叶鞘两面的薄壁细胞较小,维管束间的薄壁细胞较大。不同于叶柄,叶鞘维管束只有1排。不同竹种维管束的间距不同,勃氏甜龙竹与绵竹维管束间距相对较远,慈竹、香竹、云南箭竹和美竹相对较近。叶鞘维管束也分为大、小维管束,相间排列,所有竹种叶鞘维管束与叶片维管束解剖结构差异较大,而与叶柄维管束一致,也不具有叶片维管束所具有的薄壁鞘细胞。叶鞘维管束鞘均为单层细胞鞘,鞘细胞细胞壁发生显著的加厚及木质素沉积(番红染成红色),维管束内部维管分子之间薄壁细胞也显著加厚。与叶柄相似,叶鞘韧皮部纤维鞘细胞(远轴面方向)十分发达,向外与远轴面表皮连接。大维管束发育程度较高,具有完整的韧皮部、初生导管和后生导管及纤维鞘结构,小维管束横切面面积较小且发育程度较低,会出现只有1个木质部导管的情况,结构简单(图3 Ac—Fc)。
不同竹种叶鞘维管束面积也有较大的差异。勃氏甜龙竹叶鞘维管束面积最大,其木质部、韧皮部面积也最大。同叶片、叶柄维管束一样,叶鞘维管束的韧皮部面积及其维管束面积占比均大于木质部。在小维管束中,往往出现木质部发育不完善而韧皮部较为发达的情况(表1)。因此,叶鞘作为运输器官,与叶柄一致,其有机物运输组织更为发达。
2.1.5 叶枝
本研究选择叶鞘包裹着的小枝作为6种竹子的叶器官维管束解剖结构的对照,分析不同叶器官维管束与枝维管束解剖结构上的异同。不同于叶器官,叶枝呈圆柱形,其近轴面处于枝腔内壁,远轴面表皮为叶枝外表皮。叶枝近轴面相当于竹秆的竹黄处,由排列不规则的薄壁细胞组成,不同于叶鞘表皮细胞为整齐的长方形或长圆形,无法对其进行测量。远轴面表皮(外表皮)为单层细胞构成,细胞呈近圆形,细胞排列整齐,细胞外壁加厚,气孔器穿插于远轴面表皮中。
叶枝基本组织的薄壁细胞由内向外逐渐变小。散生在基本组织中的维管束由内而外逐渐变小呈放射状分布。慈竹与绵竹叶枝维管束横切面上由内而外分布为3层,勃氏甜龙竹、香竹、云南箭竹和美竹,维管束分布为4层。勃氏甜龙竹、慈竹、绵竹远轴面最小的维管束与远轴面表皮之间有1~2层细胞,而香竹、云南箭竹有3层,美竹有4~5层细胞(图4 Ad—Fd)。

d.叶枝twig.1.100×;2.400×.图4 6种竹子叶枝解剖形态特征Fig.4 Anatomical and morphological characteristics of six bamboo species leaf twigs
6个竹种叶枝的维管束都为单层维管束鞘,维管束周围无薄壁鞘细胞和明显的纤维鞘细胞,与竹秆维管束形态存在差异[8-11],也与叶片、叶柄、叶鞘维管束(图3)均有一定的差异。不同竹种叶枝维管束面积也有一定的差异,其中勃氏甜龙竹叶枝维管束面积显著大于其他竹种叶枝的维管束面积,其他竹种之间的叶枝维管束面积差异不显著(表1)。与叶片、叶柄及叶鞘一致,叶枝维管束也表现出韧皮部均朝向远轴面,木质部均朝向近轴面规律。相较叶片、叶柄与叶鞘而言,叶枝维管束导管发达,但各竹种叶枝维管束总体也表现出韧皮部面积占比大于木质部,远轴面小维管束几乎被韧皮部占据,说明叶枝有机物运输功能较强(图4)。在6种竹子叶枝中,维管束均没有厚壁组织的存在,而竹子叶枝在生长过程中叶鞘始终宿存,说明叶鞘对叶枝的机械支持功能起到了至关重要的作用。
2.2 6种竹子秆箨解剖结构比较
竹类植物的秆箨,交替着生在新竹茎秆的节部,主要由箨鞘与箨片两部分组成。基于野外观察发现,竹子新秆上、下部位的箨片形态差异较大。通常秆下部的箨片较短、质地较硬,而秆上部的箨片细长如叶片,因此对两种类型的秆箨分别进行比较与分析是必要的,6种竹子秆箨解剖结构见图5。

e.秆上部箨片upper culm blade;f.秆下部箨片lower culm blade;g.秆上部箨鞘upper culm sheath;h.秆下部箨鞘 lower culm sheath.1.100×;2.400×.Bar=100 μm.图5 6种竹子茎生叶(秆箨)解剖形态特征Fig.5 The anatomical and morphological characteristics of six bamboos species stem leaves (culm sheaths)
2.2.1 秆上部箨片
竹子的箨片与其他器官基本结构一致,均包括表皮、基本组织和维管束3个部分。箨片表皮与叶片形态一致,均包括近轴面与远轴面表皮组织,箨片中央具有明显的主脉。不同竹种间箨片厚度存在差异,具体表现为绵竹的箨片最厚,达262.15 μm;云南箭竹的箨片最薄,为129.32 μm(表1)。
1)秆上部箨片表皮。箨片近轴面表皮为单层细胞,厚度均大于远轴面(表1)。不同竹种间,香竹近轴面表皮厚度最厚(16.33 μm),最薄的为云南箭竹(9.86 μm),勃氏甜龙竹与绵竹比较接近,分别为12.15与12.62 μm。箨片近轴面表皮细胞近圆形,高度角质化有乳突,两个维管束之间分布着大小不一的泡状细胞组成扇形的泡状细胞复合体。近轴面表皮因泡状细胞下陷、维管束位置突出,使箨片近轴面表皮呈波浪状起伏(图5 Ae—Fe)。
箨片的泡状细胞复合体与叶片的泡状细胞复合体形态差异显著。如勃氏甜龙竹叶片泡状细胞复合体一般多数由3个泡状细胞组成,泡状细胞整体与近轴面表皮在同一平面,呈扇形,而箨片泡状细胞复合体一般由6个泡状细胞组成,且下陷呈马鞍状(图5 Ae)。泡状细胞复合体的面积,不同竹种也有差异,具体表现为勃氏甜龙竹远大于其他竹种,为4 628.46 μm2,最小的为绵竹(1 158.58 μm2),其次为香竹(1 247.57 μm2)(表1)。6个竹种箨片的远轴面表皮均为单层细胞,厚度小于近轴面表皮(表1)。表皮细胞近圆形,外壁加厚有角质化的针状或圆锥状乳突,外壁加厚程度低于近轴面表皮,气孔器穿插于远轴面表皮中(图5 Ae—Fe)。
2)基本组织。6个竹种秆上部箨片的基本组织主要由叶肉组织和梭形细胞收缩后的空穴构成(图5 Ae—Fe)。与叶片一致,箨片维管束的两侧均可见空穴,空穴将维管束间的叶肉组织分为近轴面与远轴面两部分。除慈竹以外,箨片近轴面与远轴面的叶肉细胞与叶片的叶肉细胞存在显著差异,箨片叶肉细胞均呈圆形或者椭圆形,并未分化出叶片那样整齐的指状臂细胞和梅花状细胞。慈竹箨片叶肉细胞形态接近叶片叶肉细胞形态。
3)维管束。所有竹种秆上部箨片维管束形态与叶片一致,具有大、小两种维管束,大维管束具有薄壁与厚壁2层鞘细胞,小维管束具有1层鞘细胞。维管束面积在种内、种间都存在较大差异(表1)。与营养叶器官不同,箨片维管束中的木质部更加发达,在维管束中的面积占比巨大,其中勃氏甜龙竹、美竹等都达到40%,而韧皮部仅占5%左右,其余4个竹种的木质部在维管束中的面积占比都大于韧皮部(表1)。秆上部箨片与叶片在叶肉组织与维管束解剖结构的显著差异,说明了箨片与叶片的功能侧重不同,叶片是以有机物运输为主,而箨片以水分运输为主。
2.2.2 秆下部箨片
新竹秆下部的秆箨通常为竹笋的笋壳。该部位的箨片与新竹梢部秆箨的箨片相比,形状短小而尖,质地较硬,与叶片形态差异显著。除香竹外,其他竹种秆下部箨片厚度均明显大于秆上部箨片。
1)表皮。与秆上部箨片类似,秆下部箨片近轴面表皮由单层细胞构成,细胞近圆形,细胞排列紧密,但排列不整齐,表皮上方有少量角质化的锥状乳突(图5 Af—Df)。不同竹种近轴面表皮厚度也具有显著差异,表现为绵竹近轴面表皮厚度最厚,为15.28 μm,最薄的为慈竹(9.45 μm)(表1)。勃氏甜龙竹、慈竹、绵竹与香竹的箨片近轴面表皮能看到少量呈扇形的泡状细胞复合体(图5 Af—Df)。云南箭竹与美竹无法辨别出泡状细胞(图5 Ef、5Ff)。竹类植物的秆下部箨片泡状细胞少,与其不容易发生卷曲相一致。不同竹种秆下部箨片的远轴面表皮均为单层细胞构成,厚度小于近轴面表皮(表1),细胞近圆形,排列更加整齐紧密。
2)基本组织。秆下部箨片基本组织主要由薄壁细胞构成,与叶片及秆上部箨片相比,无梭形细胞、指状壁细胞及梅花状薄壁细胞的分化,所有薄壁细胞呈圆形或长椭圆形,这也反映了秆下部箨片并不以光合作用为主要功能(图5Af—Ff)。箨片中的薄壁细胞自中央部位向远轴面及近轴面逐渐变小、变扁,尤其远轴面靠近表皮的1~2层薄壁细胞排列整齐、紧密,这与秆下部箨片往往比其他叶器官的硬度更高相一致。
3)维管束。所有竹种的秆下部箨片、主脉维管束系统均不及叶片及秆上部箨片发达(图5 Af—Ff)。箨片维管束形态与叶片及上部箨片一致,均具有大、小两种维管束,但在解剖结构有显著差异。慈竹和绵竹箨片表现为2个大维管束间分布有2个小维管束,其余竹种均为大、小维管束相间排列。箨片的维管束包括韧皮部、木质部以及一圈维管束鞘细胞和分别朝向两边的韧皮部、木质部纤维组织(纤维鞘),但不同竹种之间略有差异。勃氏甜龙竹下部箨片维管束的韧皮部纤维鞘更发达,而慈竹与绵竹下部箨片纤维鞘则在近轴面和远轴面都有分布,近轴面为独立的纤维鞘,并不与维管束直接相连(图5 Af—Cf)。香竹与云南箭竹下部箨片的维管束韧皮部与木质部纤维鞘均比较发达,且与维管束相连(图5 Df、5Ef),美竹的箨片维管束木质部和韧皮部纤维鞘则非常发达,木质部表现为独立的纤维鞘(图5Ff)。
与叶片和上部箨片显著不同,下部箨片的维管束鞘细胞细胞壁加厚均比较显著,没有薄壁鞘细胞(图5 Af—Ff)。勃氏甜龙竹、慈竹、绵竹和香竹下部箨片维管束鞘为双层鞘细胞,云南箭竹和美竹下部箨片维管束鞘为单层鞘细胞(图5 Af—Ff)。加厚的维管束鞘细胞以及发达的纤维鞘,均与各竹种下部箨片质地较硬有关。与秆上部箨片相似,秆下部箨片维管束也表现为木质部更发达,维管束经常出现多个后生导管的现象,所有竹种下部箨片维管束的木质部面积占比均高于韧皮部(表1),这一点与营养叶器官表现刚好相反,反映了两者功能的显著差异。
2.2.3 秆上部箨鞘
箨鞘是竹类植物分类与竹种鉴定的最典型形态依据之一。不同竹种箨鞘的形态特征在相关的竹类图志与植物志中有十分详细的描述,而有关其解剖特征却缺少关注。通过比较不同竹种秆上部箨鞘的厚度,发现勃氏甜龙竹箨鞘厚度最厚(459.59 μm),其次为绵竹(360.05 μm),最薄的为美竹,仅381.10 μm(表1)。
1)表皮。秆上部箨鞘近轴面表皮为单层细胞,呈长方形,整齐排列(图5Ag—Fg),不同竹种之间近轴面表皮厚度有一定差异,具体表现为慈竹与美竹近轴面表皮厚度最厚,分别为7.87和7.02 μm,勃氏甜龙竹最薄,为4.93 μm(表1)。远轴面表皮也为单层细胞,细胞近圆形,与近轴面表皮相比,细胞壁加厚明显,细胞排列更加紧密(图5 Ag—Fg)。勃氏甜龙竹、慈竹、绵竹和香竹远轴面表皮层与表皮层下的1层细胞的细胞壁均加厚显著(图5 Ag—Dg),而云南箭竹和美竹只有表皮层细胞的细胞壁加厚(图5 Eg—Fg),不同于叶鞘等其他器官仅是外壁加厚。
2)基本组织。箨鞘的基本组织主要由薄壁细胞构成,无梭形细胞。中央的薄壁细胞巨大,近圆形,远、近轴面的薄壁细胞逐渐变小(图5Ag—Fg)。同叶鞘解剖结构一致,缺乏梭形细胞的分化,尤其缺乏类似叶片中的指状臂细胞和梅花状细胞的分化,说明箨鞘不是一个以光合功能为主的器官。
3)维管束。不同竹种秆上部箨鞘的维管束面积大小差异显著,表现为绵竹箨鞘维管束面积最大,香竹的面积最小(表1)。6个竹种的秆上部箨鞘在横切面上均没有主脉维管束。与其他叶器官一致,秆上部箨鞘维管束也分为大、小两种维管束,且不同竹种箨鞘维管束之间排列规律有一定的差异。勃氏甜龙竹、云南箭竹和美竹大、小维管束相间排列(图5Ag、5Eg、5Fg),而慈竹、绵竹和香竹两个大维管束之间隔着2个以上的小维管束(图5Bg、5Cg、5Dg)。在分布位置上,6个竹种箨鞘维管束的分布位置基本一致,均靠近远轴面一端,相应的远轴面一端的纤维鞘组织也比较发达(图5Ag—Fg),这也与其保护功能相关。与叶片显著差异的是6个竹种箨鞘的维管束鞘细胞加厚显著,不存在薄壁鞘细胞,与叶鞘维管束鞘细胞一致(图5Ag—Fg)。与箨片相同,所有竹种箨鞘维管束的木质部面积占比均高于韧皮部(表1)。在小维管束中,维管分子主要由木质部构成,这也反映了竹子箨鞘主要以水分运输为主(图5Ag—Fg)。
2.2.3 秆下部箨鞘
秆下部箨鞘与秆上部箨鞘在形态上类似。但秆下部的箨鞘厚度往往比上部的箨鞘更厚、更硬。从横切面上,也可将其分为表皮、基本组织与维管组织。
1)表皮。秆下部箨鞘近轴面表皮为单层细胞,细胞呈长方形,整齐紧密排列(图5)。在表皮厚度方面,表现为勃氏甜龙竹、慈竹与绵竹箨鞘近轴面表皮较厚,香竹、云南箭竹与美竹相对较薄(表1)。远轴面表皮也为单层细胞,细胞呈正方形,排列紧密,外向细胞壁加厚显著(图5 Ah—Fh)。不同竹种箨鞘在远轴面表皮的厚度也表现出差异,其中勃氏甜龙竹与香竹相对较厚,其他竹种相对较薄。
2)基本组织。秆下部箨鞘基本组织与秆上部箨鞘类似,箨鞘中央的薄壁细胞巨大,向远轴面与近轴面逐渐变小,无梭形细胞的分化(图5 Ah—Fh)。虽然幼竹秆的下部箨片也没有梭形细胞的分布,但总体而言,梭形细胞是叶片(箨片)器官与叶鞘(箨鞘)器官的关键区别之一。此外,与上部箨鞘一致,秆下部箨鞘缺乏类似叶片的指状臂细胞与梅花状细胞的分化,这也反映了光合并不是箨鞘的主要生理功能。
3)维管束。秆下部箨鞘维管束与秆上部箨鞘形态类似(表1)。与箨片相比,慈竹与美竹秆下部箨鞘的维管束面积大于秆下部箨片的面积,其余竹种维管束面积小于秆下部箨片的面积(表1)。秆下部箨鞘维管束鞘细胞与上部箨鞘维管束一致,维管束鞘细胞均发生显著加厚,不存在薄壁鞘细胞,这与叶片及箨片的维管束存在显著的区别(图3Aa—Fa、图5 Ae—Fe和图5 Af—Ff)。此外,所有竹种秆下部箨鞘维管束中的木质部维管束面积占比均大于韧皮部,但小于秆下部箨片维管束中的木质部面积占比(表1),推测秆下部箨鞘在水分运输上起着至关重要的作用。
3 讨 论
营养器官外部形态特征一直作为竹类植物分类的重要依据[12-14]。早在1907年Brandis[15]就对竹叶片的解剖构造进行了系统的研究。近年来,Vega等[16]提出梭形细胞随着生长会逐渐收缩形成“I”形的梭形细胞并留下1个空穴,为梭形细胞的研究提供了一定的理论信息。丁雨龙等[17]从多学科的角度总结、分析了竹类植物不同器官的发育生物学研究。苏佳露等[18]对菲白竹(Pleioblastusfortunei)等6个竹种叶片进行三维结构的构建,为了解竹叶内部结构及竹子系统演化与分类提供一定的借鉴。李璟等[19]通过对云南箭竹3个变异类型的叶片进行解剖,认为其叶片的形态和解剖结构特征在讨论种下分类水平上具有一定的分类价值。但部分学者认为叶片的解剖构造在分类上没有实质性的意义,因为环境、种源、生长状态、取样位置等都会造成叶片解剖构造的差异[20-25]。黄玲等[26]通过对不同栽培条件下的勃氏甜龙竹苗叶片解剖结构进行观察,证实了组培苗抗逆境能力弱,必须经过炼苗驯化才能移栽,炼苗驯化的过程其实就是组织解剖结构逐渐适应环境的过程。张月高等[27]对慈竹扦插苗移栽后不同时期叶片进行组织解剖观察,发现叶片与角质层及上下表皮细胞逐渐增厚,叶片增厚的过程也是慈竹扦插苗对环境适应能力增强的过程。这些研究均反映了竹子叶片对环境的生理响应。
在竹类植物中,叶器官主要包括两大类,营养叶和茎生叶/秆箨,这两类叶器官除了着生的位置不同,组成基本一致,均包括叶片/箨片和叶鞘/箨鞘两部分。两者之间的区别在于营养叶在叶片与叶鞘之间具有叶柄这一组织,而秆箨往往缺乏这一组织。从器官演化的角度上来看,营养叶和茎生叶均属于同源器官的演化,叶片与箨片为同源器官,箨鞘与叶鞘为同源器官。然而,研究者们忽略了箨鞘/叶鞘/是不是叶柄或者叶片的同源演化,即箨鞘与叶鞘本身到底是变态叶、变态叶柄还是变态枝的问题。
丁雨龙等[8]认为竹类植物的叶鞘与叶片之间维管束密集的部分为叶柄,但未对箨鞘的形成进行说明。结合传统植物学观点,叶柄是连接叶片和茎秆的部分[28],因此竹类植物的叶鞘或者箨鞘是叶柄向下延伸扩展包裹枝的器官。兰科植物中也报道了类似的观点[9],在其他植物中,叶柄与枝之间存在着腋芽,而竹类植物的箨鞘或叶鞘着生在茎秆与叶枝的节上,也包裹着茎秆或叶枝的节芽。由于箨鞘/叶鞘与叶柄在空间位置上的联系及其相似的解剖结构,因此竹类植物的鞘器官更可能与叶柄同源,而不是枝同源,Wang[3]认为箨鞘是枝变态并不精确。此外,本研究发现6个竹种的箨鞘/叶鞘与箨片/叶片在解剖上存在显著的区别,箨鞘与叶鞘不存在主脉维管束,也不存在叶片及幼竹上部箨片常见的梭形细胞。在维管束结构上,箨鞘与叶鞘不具有叶片及箨片维管束常见的发达薄壁鞘细胞,反而均为厚壁鞘细胞,并且维管束具有发达的韧皮部纤维鞘。由于叶片及箨片与叶鞘及箨鞘在解剖结构上差异巨大,不支持传统观点认为的叶鞘或箨鞘是叶片的变态,这也与Wang[3]的观点一致。
此外,箨鞘与叶鞘及叶柄的基本组织中,均无叶肉细胞所具有的指状臂细胞与梅花状细胞的分化,反而均具有发达的纤维鞘组织,说明竹类植物的鞘器官并不以光合作用为主要的生理功能,而以机械支持功能为主。本研究对叶鞘所包括的叶枝进行解剖观察发现,叶枝缺乏叶鞘及箨鞘维管束所具有的发达纤维鞘组织,仅具呈放射状的维管束,说明这些小叶枝并不具有很强的机械支持功能,而围绕在叶枝的周围宿存的叶鞘起到关键的机械支持功能。
箨片及箨鞘常作为分类上的依据[12-14],虽然不同竹种在形态上存在较大差异,但在显微解剖结构上相近。秆上部箨片与秆下部箨片是不同位置的同一器官,但结构上却表现出极大差异。秆上部的箨片结构上更接近叶片,泡状细胞和梭形细胞收缩后的空穴都很明显,而在秆下部箨片中没有观察到空穴的分布,这一发现或许有助于未来为梭形细胞功能研究提供新的思路。同时,通过对6种竹子的叶器官以及叶枝进行解剖比较,发现梭形细胞、主脉维管及薄壁维管束鞘细胞的有无,以及基本组织中薄壁细胞的分化程度(指状臂细胞和梅花状的分化),成为叶片、箨片与箨鞘和叶鞘在解剖结构上的关键区分标准。另外,竹类植物中所有器官的维管束,均表现出木质部朝向近轴面,而韧皮部均朝向远轴面这一稳定的解剖规律与特征。
通过对不同器官的韧皮部与木质部的维管束面积占比进行比较,本研究发现营养叶包括叶片、叶柄、叶鞘以及叶枝的维管束均表现出韧皮部的维管束面积占比显著高于木质部,而在秆箨/笋箨中则表现为木质部的维管束面积占比明显占优势的情况。如勃氏甜龙竹和美竹箨鞘中的木质部维管束面积占比高达40%以上,而韧皮部仅有5%左右。因此,在竹类植物中,营养叶器官以及叶枝主要以光合同化物运输为主,而秆箨在生理功能上除了机械支持作用,更多的是以水分运输为主。因此,竹类植物的茎生叶(秆箨)作为运输器官是一个以水分运输为主、有机物运输为辅的器官,这也说明了发笋期水分在幼竹生长中的巨大作用。
