炭疽病对油橄榄果实真菌群落结构的影响
2023-03-13郭海霞黄雯澜李谨宵杨育林向成华张建华
郭海霞,黄雯澜,,王 谢,李谨宵,王 锐,杨育林,向成华,张建华
(1.特色园艺生物资源开发与利用四川省高校重点实验室/成都师范学院,成都 613000;2.四川省农业科学院农业资源与环境研究所,成都 610066;3. 四川省林业科学研究院,成都 712100)
【研究意义】油橄榄(Oleaeuropaea)是木犀科木本果用兼油料作物,因其环境适应能力强、果实含油量高、油品质好、经济效益高,越来越受种植者和消费者欢迎[1-2]。全球油橄榄的生产地主要集中在地中海沿岸,主要生产国家为西班牙和意大利。2011年全球油橄榄种植面积为1.01×107hm2,橄榄油消费量达3.09×107t,呈逐年上升趋势[3]。油橄榄最早于1907年由法国传教士引入我国,1956—1962年由中国科学院等正式引入试种[4]。据统计,2021年我国油橄榄产量为7.29×104t,油橄榄主要种植区在甘肃和四川[5]。两省总产量占全国总产量的94%左右。其中,甘肃产量约4.1×104t,四川约2.6×104t。炭疽病是制约我国南方地区油橄榄产业发展的主要病害。该病主要危害果实、嫩芽、嫩叶及嫩梢,引起大量落果、落叶、枯梢,果实品质下降。病斑初为褐色小点,圆形,后扩大,中心略凹陷,可造成油橄榄大面积减产。Juan等[6]在测试了来自科尔多瓦世界橄榄种质库384个橄榄品种的炭疽病易感性后指出,几乎所有西班牙油橄榄品种都可归类为极易感染、容易感染或较易感染三类。而我国大多数推广栽培品种来源于西班牙。因此,掌握感染炭疽病油橄榄果的真菌区系变化特征对我国油橄榄产业的发展具有重要的现实意义。【前人研究进展】炭疽病是一种危害性较强的真菌性病害,主要危害叶片和果实,常引起大量落叶、落果和枯梢,以及果实品质和产量降低[7]。炭疽病的病原菌主要为刺盘孢属(Colletotrichum)真菌,可侵染草莓、茶树、辣椒、油橄榄等30多种植物,严重损害植物品质,造成较大的经济损失[8-11]。时全昌[12]1979年研究指出油橄榄炭疽病是由刺盘孢菌属(Colletotrichumsp.)侵染所致。2018年,张知晓等[11]对云南永仁油橄榄炭疽病病原菌进行了分离鉴定,进一步确定了我国油橄榄炭疽病的主要病原菌为盘长孢状刺盘孢(C.gloeosporioides)。同时,王敬文[13]研究发现,盘长孢状刺盘孢主要以孢子侵染果实,侵染后能分泌对油橄榄组织发生毒害的化合物,该毒素对油橄榄原生质体具有毒害作用,能使最敏感的愈伤组织在短时间内坏死,从而毒害油橄榄。【本研究切入点】虽然当前研究已经分离鉴定出油橄榄炭疽病的病原菌,并形成一定的防治技术[14],但对炭疽病病原菌与健康果实原有微生物之间的互作关系尚不知晓。可从微生物生态学角度,进一步探明感染炭疽病后油橄榄果实内微生物结构和多样性变化,为全面解析感染炭疽病后油橄榄果实各部位微环境和生理变化提供依据[15]。另外,日益成熟的高通量测序技术[16]为微生物多样性研究提供了更加可靠的技术支撑。【拟解决的关键问题】本研究采用高通量测序技术,探究炭疽病油橄榄果实和健康果实的真菌群落结构及多样性的差异,为油橄榄炭疽病的综合防治提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于四川省成都市金堂县淮口产业科技综合示范园油橄榄种植区(104°08′ E,30°11′ N,海拔432 m),属于亚热带季风气候。种植区于2009年开始种植油橄榄,栽培密度4 m×4 m,主要栽培品种有阿贝基娜、皮瓜尔和莱星,种植面积约5153.33 hm2。区内气候条件、栽培管理措施和水肥用量基本一致,年平均气温16.9 ℃,年均降水量760 mm,湿度大,年平均日照1295.5 h,土壤类型为黄壤。研究区内油橄榄炭疽病在各品种上均有发生。油橄榄炭疽病主要影响果实,对树木生长无影响。
1.2 样品采集
2020年12月10日,于研究区内选择3块空间位置相对独立、种植面积不小于300 m2且有油橄榄炭疽病发生的橄榄种植田块进行调查。每个田块上不分品种随机采集健康果实和发病果实,按照发病程度分为健康果实(CK)、发病前期果实(MP)和发病后期果实(SP)。果实未变质为健康果实,果实变质部分少于1/2为发病初期,果实整个部分全部变质为染病发病后期(图1)。每个园区CK、MP和SP的样品采集量不低于100 g。采集后当日带回实验室,按照无菌棉花富集法[17]采集果皮(Peel)上的真菌群落,然后果实表面消毒后在无菌条件下将果皮、果肉、种子完全分离开来。将无菌分离的果皮放入棉花富集法获得的样品中合为1个果皮真菌检测的总样品,将无菌分离的果肉(Flesh)作为1个果肉真菌检测的总样品。采集后的样品按照发病程度、果实部位、来自第i个试验区分别编号为SP_peel_i、SP_flesh_i、MP_peel_i、MP_flesh_i、CK_peel_i和CK_flesh_i,编号后立即放入-80 ℃冰箱,用于DNA提取。

图1 健康油橄榄果实、发病初期油橄榄果实和发病后期油橄榄果实对比Fig.1 Comparison of healthy fruits,mildly anthracnose diseased fruits and severe anthracnose diseased fruits
1.3 高通量测序分析
所有样品送至生工生物工程(上海)股份有限公司进行DNA提取和ITS1-2高通量测序。试剂盒扩增引物为Read1 3′端和Read2 3′端。测序引物接头分别为AGATCGGAAGAGCACACGTCTGAACTCC AGTCA和AGATCGGAAGAGCGTCGTGTAGGGAAA GAGTGT,扩增片段长度为300 bp[18]。
1.4 统计与分析
为揭示真菌分类单元(Operational taxonomic units,OTU),本研究将序列进行归类操作(Cluster)。通过归类操作,将序列按照彼此的相似性分为许多小组,1个小组就是1个OTU。通常按97%的相似度,对所有序列进行OTU划分并进行生物信息统计分析。
为揭示处理间真菌物种组成的差异性,本研究基于ImageGP平台(http://www.ehbio.com/ImageGP/)开展韦恩(Venn)分析和群落结构分析。为比较不同组间标志性物种差异,本研究基于微科盟生科云平台(https://bioincloud.tech/)平台,开展了线性判别分析(Linear discriminant analysis Effect Size,LEfSe)。真菌功能类群利用第三方工具FUNGuild v1.0 完成。为揭示处理间物种的多样性,本研究采用ACE指数[19](Abundance-based coverage estimators)和Chao指数[20](Chao’s index)表征物种的丰富度,用Shannon 指数、Simpson指数、Shannon evenness指数代表群落内物种的多样性。Shannon指数是物种丰富度和分布均匀度的综合度量,Shannon指数越大则群落多样性越高。Simpson指数反映了群落中个体数量分布的集中度,Simpson指数越高表明群落中个体越集中在少量优势物种上,群落优势度越大,群落分布不均匀,多样性低。Shannon evenness指数描述群落均匀性,Shannon evenness指数越高,代表群落分布越均匀,多样性越高。本研究基于SPSS 23.0软件,采用单因素(One-way ANOVA)和Duncan法多重比较(α=0.05)分析了发病程度和果实部位对群落多样性指数、相对丰富度的影响(P<0.05)。图表中数据为平均值±标准差。
2 结果与分析
2.1 真菌分类单元结构特征
测序结果显示,6个处理测出真菌覆盖率均大于99%,检测结果有效(表1)。由表2可知,油橄榄果实的18个样本中共获得有效真菌序列80 182条。碱基数在10 396 294~28 667 930 bp,平均碱基数为19 637 830 bp,碱基长度在42~490 bp,真菌序列长度高度集中于200~300 bp,平均有效长度245 bp。将有效序列在97%的相似度水平下进行聚类,分别获得有效真菌分类单元(OTU)1596个。其中果皮有真菌OTU 939个,果肉有真菌OTU 921个。

表1 采集样品真菌检测覆盖率
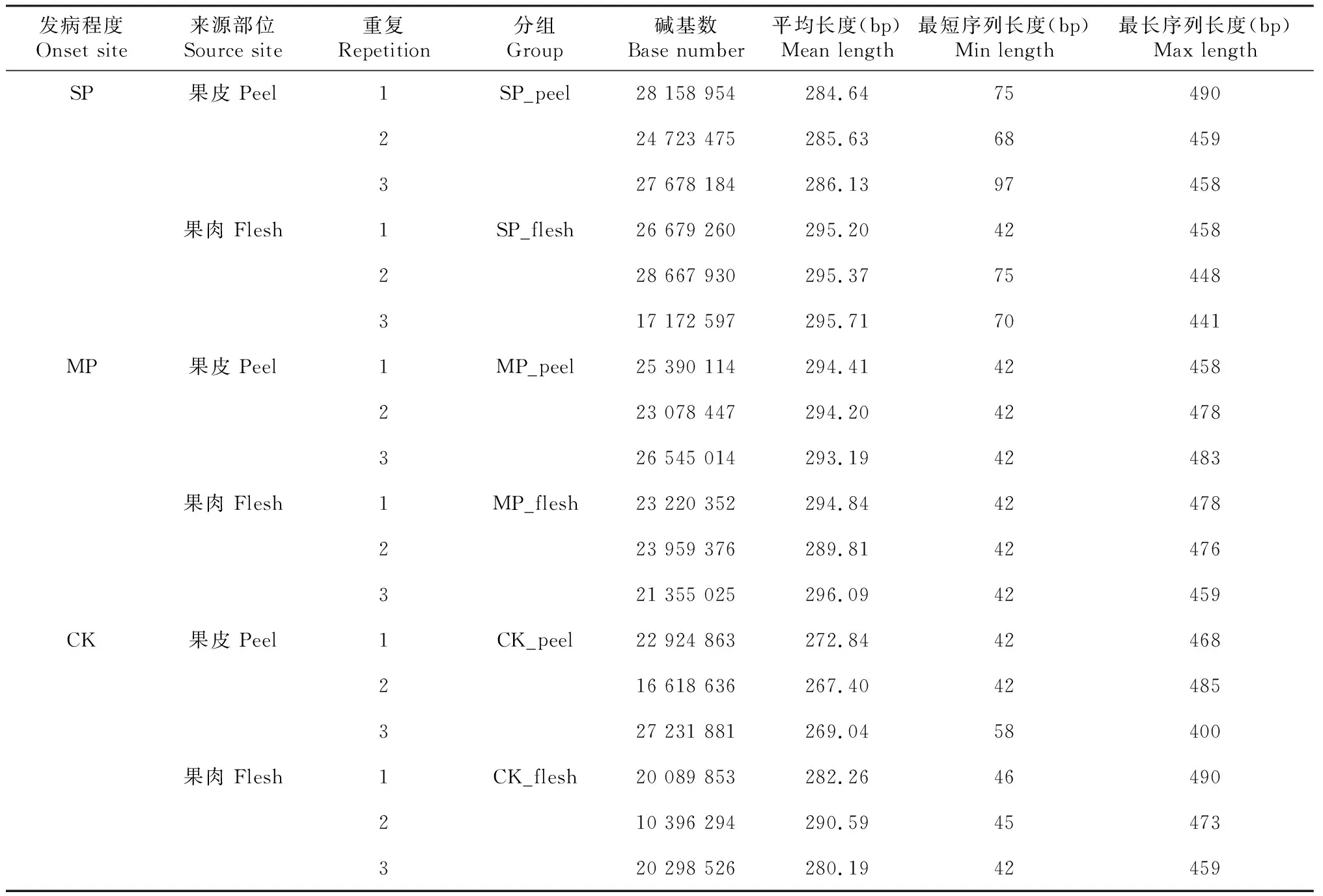
表2 各样本有效序列数据统计
由图2可知,6个分组的全部OTU为1022个。其中,共有OTU为317个,占全部OTU的34.64%。SP_peel组特有的OTU数为46,占全部OTU的4.50%;SP_flesh组特有的OTU数为18,占全部OTU的1.76%;MP_peel组特有的OTU数为31,占全部OTU的3.03%;MP_flesh组特有的OTU数为48,占全部OTU的4.70%;CK_peel组特有的OTU数为68,占全部OTU的6.65%;CK_flesh组特有的OTU数为46,占全部OTU的4.50%。综上可知,6个分组中真菌的共有菌较多,各组中特有菌占比均不高,其中CK_peel组特有菌数最多,说明健康果皮的特有真菌物种最丰富。
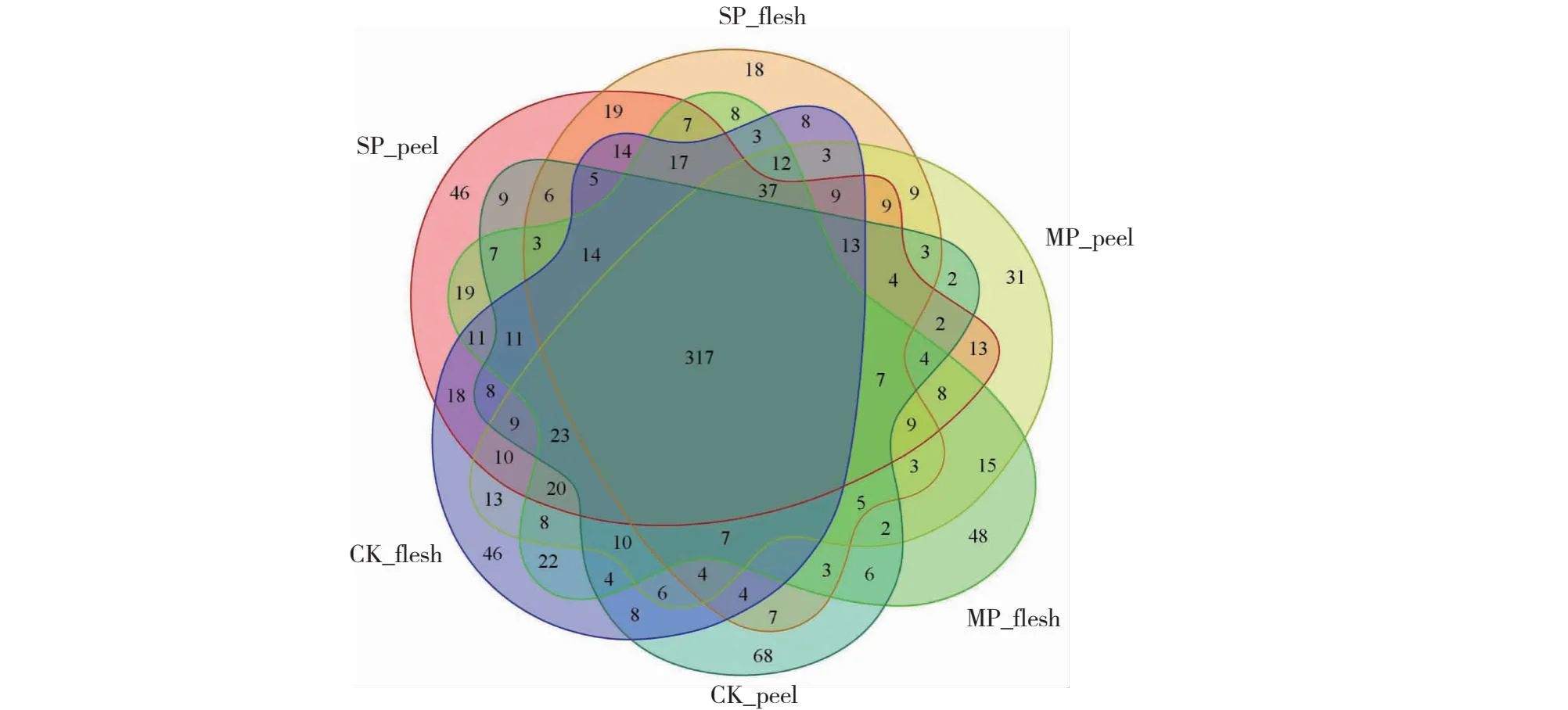
图2 六组样品中真菌韦恩图Fig.2 Venn diagram on fungi identified in six sample groups
2.2 真菌类群多样性变化分析
由表3可知,Chao指数对发病程度和果实部位均无明显响应;Ace指数对发病程度无显著响应,差异主要是由果实部位所决定,结合表4可知,果肉的Ace指数显著高于果皮。由此推断,果肉真菌的种类数高于果皮。
由表3可知,发病程度显著影响了果皮和果肉中真菌群落的多样性。由表4可知,染病后,果皮和果肉上真菌群落的Shannon和Shannon evenness指数均显著降低。随着染病程度的加重,果皮真菌的Shannon和Shannon evenness指数呈先降低后升高趋势,果肉部位则随着病情的加重呈持续降低趋势。染病后,果皮和果肉上真菌群落的Simpson指数显著升高,果肉部位随着病情的加重而持续升高,果皮部位呈先升高后降低趋势。
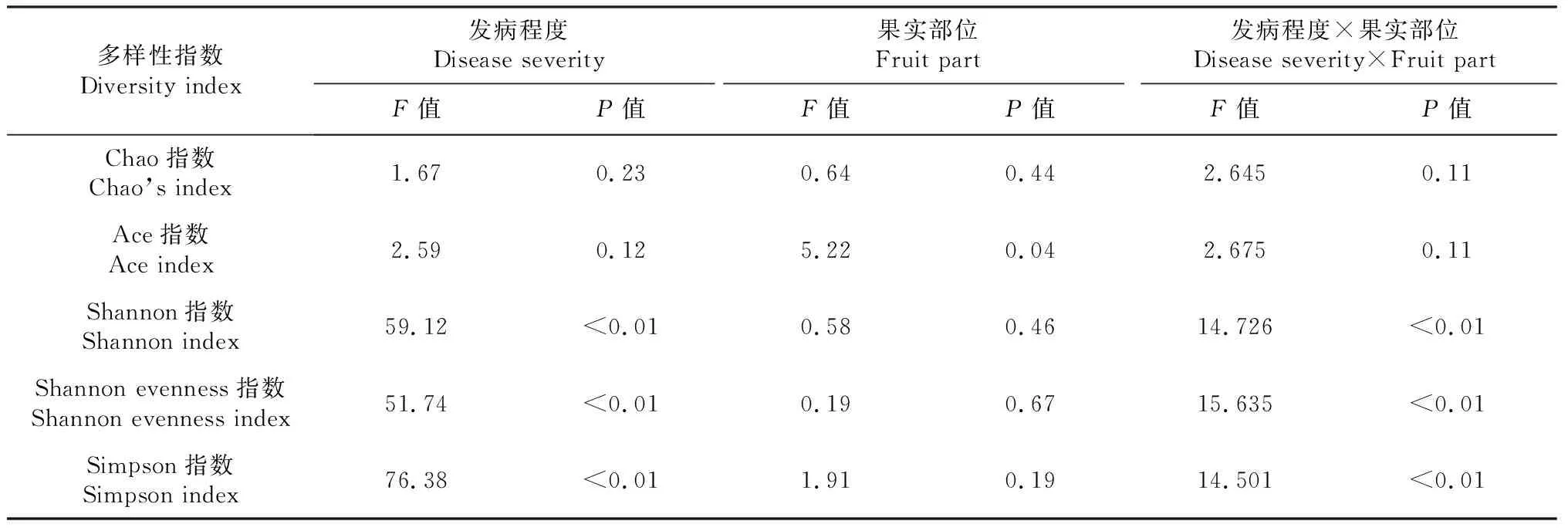
表3 发病程度和果实部位对油橄榄果实真菌群落多样性指数的影响
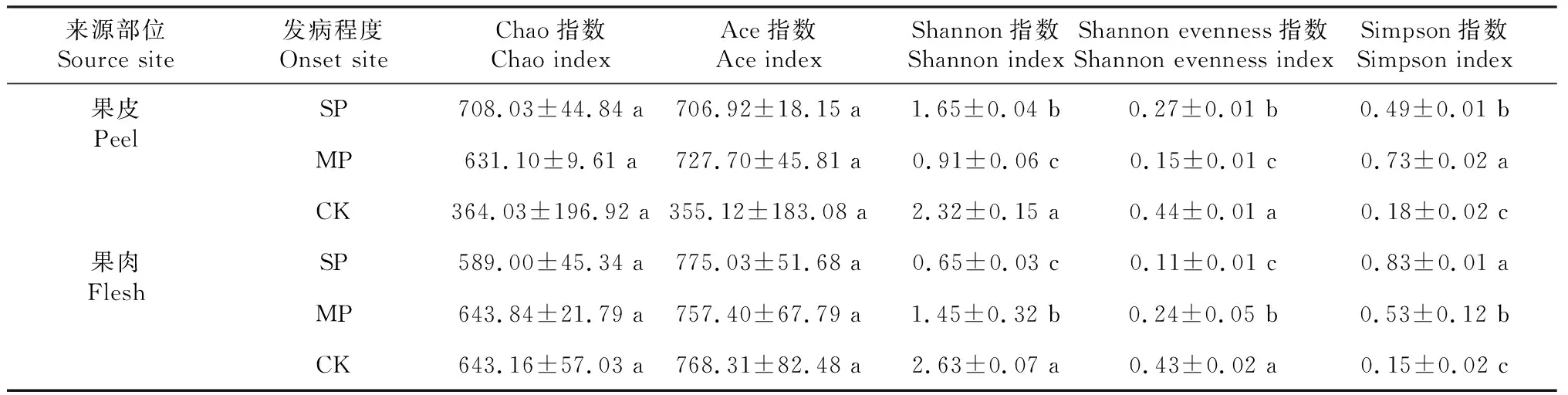
表4 油橄榄果实真菌群落多样性指数
以上结果表明,炭疽病感染显著降低了油橄榄果真菌群落的多样性,且其主要通过改变群落中个体数量在种间的分布来影响群落多样性。感染炭疽病后,真菌群落种类丰富度指数变化不明显,但优势物种数量增加显著,导致群落优势度增加,均匀度下降,多样性降低。另外,真菌群落多样性在果皮和果肉的变化趋势表现出差异,染病后期果皮真菌多样性呈回升趋势。
2.3 不同分类水平上真菌相对丰度的响应
样品检测出的真菌分属于2门9纲17目21科25属。其中,健康的果皮中有2门6纲12目11科12属13种,患病果皮中有2门5纲7目7科8属11种;健康果肉中有2门5纲10目12科13属16种,患病果肉中有2门5纲10目7科4属7种。各时期和各部位的优势菌门均为子囊菌门(Ascomycota)。
由表5可知,排名前5的真菌类群均对发病程度有显著响应。在不同分类水平上,对果实部位有显著响应的包括门水平下的子囊菌门(Ascomycota)和担子菌门(Basidiomycota),纲水平下的银耳纲(Tremellomycetes),目水平下的座囊菌目(Dothideales)和煤炱目(Capnodiales),科水平下的短梗霉科(Aureobasidiaceae)、锥梗菌科(Dissoconiaceae),属水平下的短梗霉属(Aureobasidium)、乌氏霉属(Uwebraunia),以及种水平下的黑酵母菌(AureobasidiumpullulansSH1515141.08FU)和UwebrauniaaustraliensisSH1930250.08FU。此外,除掷孢酵母属(Sporobolomyces)及其下的Sporobolomycessymmetricus外,在门、纲、目、科、属、种水平上排名前5的真菌类群均对发病程度与果实部位的交互作用具有显著响应。表明炭疽病的发生显著改变了真菌群落结构,且不同菌群在果皮、果肉的变化存在一定差异。
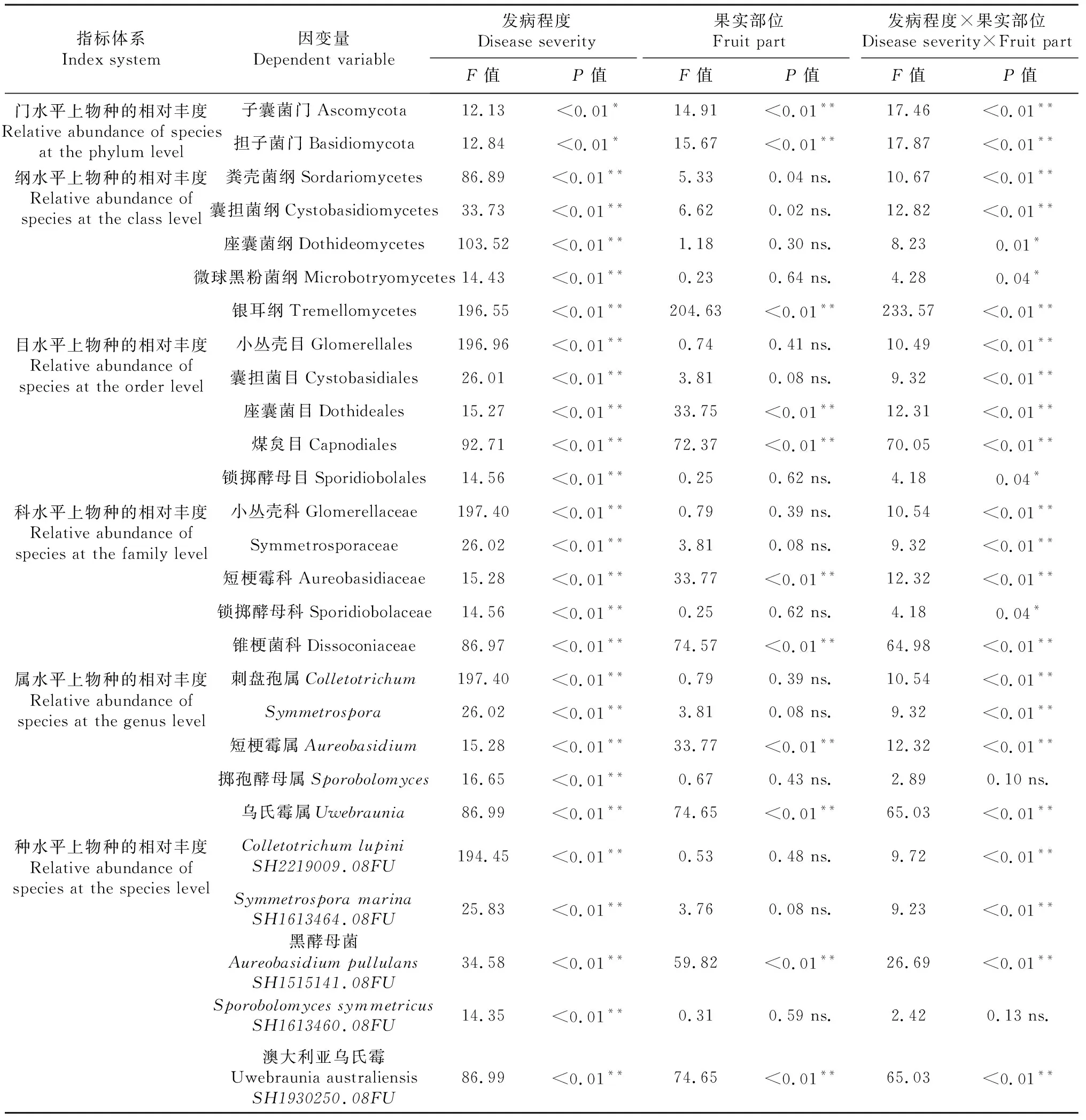
表5 不同分类水平上真菌相对丰度对发病程度和果实部位的响应
2.3.1 果皮真菌相对丰度对发病程度的响应 从门水平上看,在健康果实果皮上,子囊菌门占比84.71%,染病初期上升至94.93%,染病后期又下降至86.03%。与之相反,健康果皮中担子菌门占比15.03%,染病初期下降为4.63%,染病后期增加至13.47%(图3-A)。
从属水平上看,健康果皮中排名前5的优势菌属分别是假尾孢属(Pseudocercospora)、核盘菌属(Sclerotinia)、乌氏霉属、Symmetrospora、刺盘孢属,占比分别为33.97%、16.74%、10.67%、10.62%、9.39%。发病后假尾孢属和核盘菌属占比均小于0.01%,说明炭疽病对其具有明显的抑制作用。病原菌刺盘孢属在健康果皮中占比仅为9.39%,在发病后相对丰度迅速升高,达到86.97%,但到发病后期,下降到70.02%。此外,在健康和初病油橄榄果果皮中占比均较少的Bisifusarium属、汉纳酵母属(Hannaella)、镰刀菌属(Fusarium)真菌在染病后期数量大量增加,其相对丰度增长超过10倍(图3-B)。
2.3.2 果肉真菌相对丰度对发病程度的响应 在果肉中,健康组子囊菌门占比89.85%,发病初期为91.09%,发病后期为97.41%,呈随炭疽病加重而占比增加趋势。而担子菌门在健康果实果皮中占比9.67%,染病初期为8.46%,染病后期为2.14%,呈随炭疽病加重而占比降低趋势。此外,子囊菌门在各个时期均为占比约90%的优势菌门(图4-A)。
在属水平上,健康果肉中排名前5的优势菌属分别是黑孢霉属(Nigrospora)、短梗霉属、刺盘孢属、unclassified_Pleosporales属、蓝盘伯菌属(Lambertella),占比分别为30.28%、21.13%、12.28%、7.10%、6.91%。发病后,黑孢霉属占比小于0.50%,说明炭疽病对其有明显的抑制作用。短梗霉属占比也明显降低,发病初期下降至10.6%,染病后期占比仅1.59%,下降92.47%。而刺盘孢属在发病中期占比上升至72.31%,在发病后期占比增加至91.38%。说明炭疽病病原菌刺盘孢属在发病后迅速成为绝对的优势菌群。此外,在健康果肉中占比较少的假丝酵母菌属(Candida)在发病初期增加最迅速,相对丰度增长超过30倍(图4-B)。
2.4 真菌群落的LEfSe差异分析
采用LEfSe[21]分析具有统计学差异的生物标志物(Biomarker),不同样本中的生物标志物通过线性判别分析效应大小,LDA score大于3.0的为显著差异物种。分析结果(图5)显示,炭疽病感染降低了油橄榄果的标志真菌种类的数量,改变了标志真菌的种类。健康果皮的差异真菌有2纲5目7科9属,染病初期、后期均仅有1纲1目1科1属;健康果肉的差异真菌有3纲8目12科14属,CK和SP处理也仅有1纲1目1科1属。果皮上,健康果皮的差异指示菌属为假尾孢属和核盘菌属在内的9个属;MP处理的差异指示菌属为刺盘孢属,染病后期的差异指示菌属则为汉纳酵母属;果肉上,健康果肉的差异指示菌属为短梗霉属和黑孢霉属在内的14个属,染病中期的差异指示菌属为假丝酵母菌属,染病后期的的差异指示菌属为刺盘孢属。
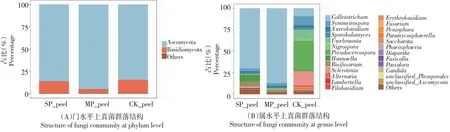
图3 果皮真菌群落结构Fig.3 Structure of fungi community in peel
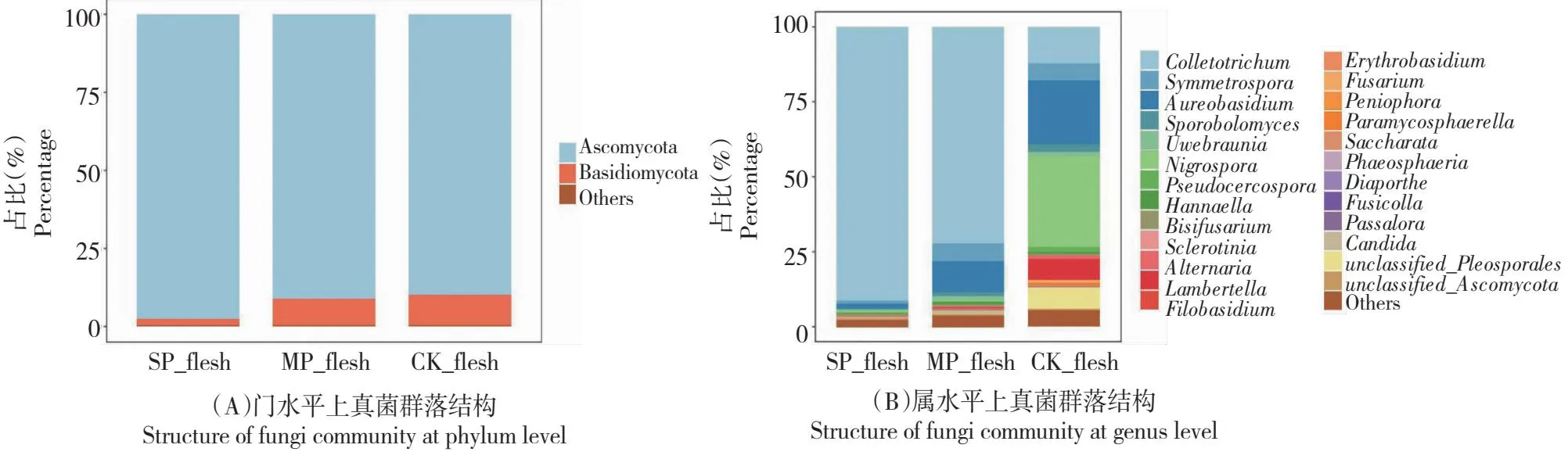
图4 果肉真菌群落结构Fig.4 Structure of fungi community in flesh
2.5 真菌的营养结构分析
从表6可以看出,在果皮部位,各营养结构的真菌丰富度在不同患病期的数量呈4种变化趋势。首先,除了兼共营养和腐生病理型、腐生共生型外,其余几类真菌在健康果皮上的丰富度均与患病时存在显著差异。其中,病理营养型、腐生病理型、腐生营养型CK均显著高于SP和MP,共生病理型CK显著低于SP和MP,而健康果皮上(CK)共生营养型真菌显著低于患病后期(SP)但显著高于患病初期(MP)。
在果肉部位,仅共生病理型真菌在患病后相对丰度显著升高,CK显著低于SP和MP。其余6类真菌的相对丰度在患病后均表现为显著降低,CK显著高于SP和MP。
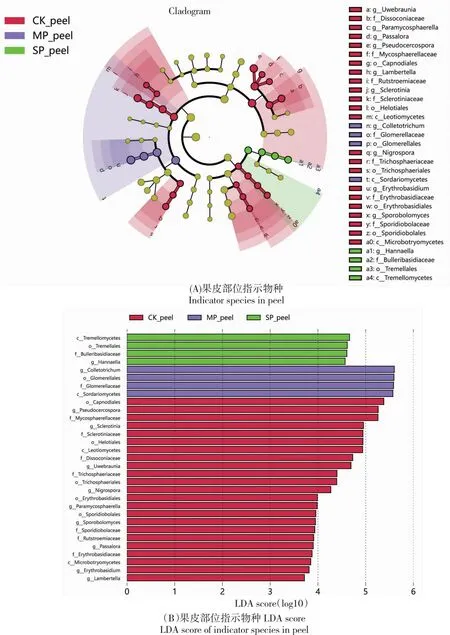
图5 基于LEfSe分析的各处理的指数物种Fig.5 Indicator species for each treatment based on LEfSe analysis
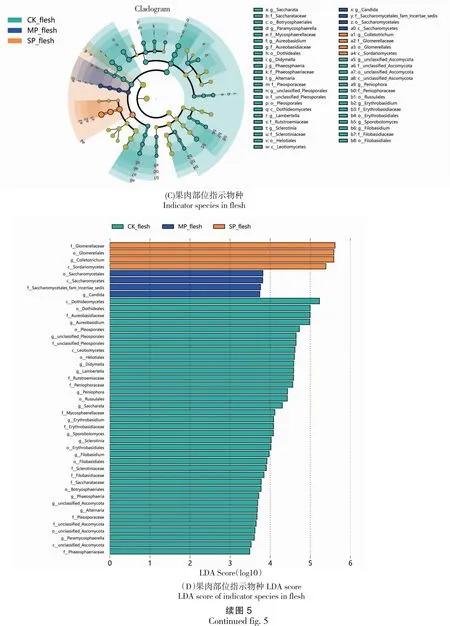
从以上结果可知,共生病理型是油橄榄果实感染炭疽病菌后的优势菌群;除共生营养型真菌外,其他营养型的真菌在感染炭疽病后,果皮和果实部位的变化都较一致,患病后共生病理型真菌数量均呈上升趋势,病理营养型、腐生营养型、兼共营养和腐生病理型、腐生营养型真菌均呈下降趋势。
3 讨 论
内生菌是植物与微生物长期协同进化形成的结构复杂并处于动态变化的功能体,广泛存在于植物的根、茎、叶、花、果实、种子等各个器官中,对植物的生长、繁殖和健康发挥着重要作用[22-24]。本研究通过高通量测序手段研究了炭疽病感染对油橄榄果实内生真菌群落的影响。结果表明,油橄榄果感染炭疽病后,真菌群落多样性显著降低。这与预期及已有的诸多研究结果相似,已有的研究表明,植物染病后,病原菌的侵染往往会打破植物体内微生物群原有的稳态结构[22]。彭芳芳等[25]对桑果内生菌的研究发现,感染菌核病的桑果,其果实内生真菌多样性远小于健康果实。与之相似,刘晓菲等[26]对黄龙病罹病柑橘叶片的研究,以及谢宪等[27]对感染枯梢病赤松针叶的研究,也证实病原菌侵染会降低植物内生真菌群落的多样性。这可能是因为病原菌的大量繁殖,一方面改变了内生真菌的生态环境,使之不利于原有菌群的生长,另一方面大量的病原菌通过竞争空间、资源或分泌有害物质,抑制原生真菌的生长。金勤[28]研究指出,炭疽病主要通过产生侵入钉侵入寄主细胞,侵入后从表皮细胞逐渐扩展到栅栏组织,然后进入到海绵组织中,逐渐杀死周围细胞。本研究结果显示,炭疽病病原菌的扩张是导致油橄榄果真菌群落多样性降低的重要因素。多样性和丰富度指数的变化也显示,染病后油橄榄果中真菌种类数量相对稳定,但是优势菌群数量扩张,显著抑制了其他菌群的生长,改变了群落内部结构,导致真菌群落的多样性显著降低,而最大的优势菌群则是炭疽病病原菌。各分类水平上各菌群相对丰度的分析结果进一步证实了这一观点。在门水平上,果皮和果肉的优势菌门在各个时期都为子囊菌门,在属水平上,染病后刺盘孢属迅速成为绝对的优势菌群,占比始终高于70%。

表6 各处理真菌的营养结构差异
植物抗病性与植物内生真菌多样性、群落结构息息相关。生态系统微生物群落多样性越低,植株抵抗病原菌侵害的能力越低[29-31]。感染炭疽病后油橄榄果真菌多样性降低,暗示其抗病能力可能降低。而研究结果显示,油橄榄果感染炭疽病后期,除了炭疽病病原菌,果皮中其它病原菌菌属,如Bisifusarium属、汉纳酵母属、镰刀菌属在重病时期也明显增加。这一结果说明,炭疽病发病后期,油橄榄果可能会并发多种病害。姜莉莉等[32]对染病“香玲”核桃果的研究也发现类似现象。感染炭疽病后,核桃果中其他病原菌,如赤霉菌属、葡萄座腔菌属(干腐病菌)等病原菌的相对丰度也明显提高。由此可见,炭疽病防治不仅要防治炭疽病,还要全面考虑其他病原菌的发生情况,进行综合防治。
本研究还发现,在炭疽病侵染过程中,油橄榄果皮、果实真菌群落多样性变化趋势呈现差异性。果皮真菌群落的多样性在染病后期有所回升,这可能是由于果皮启动了对病菌的防御机制。已有的研究发现[7,33],感染炭疽病后,植物果皮中抗真菌二烯、酚类物质等浓度升高,可有效抑制炭疽病病原菌的生长。此外,植物内生菌中有一些是天然的生防菌,能抑制病原菌的生长,并且在与病原菌共处的过程中种群及其抗菌能力得到强化[30-32]。本研究中,染病后期果皮上共生营养型真菌数量显著回升,且显著高于健康果实,双胞双镰孢(Bisifusariumdimerum)、Peniophorarufomarginata、Fusicollaviolacea这些共生营养型真菌有可能就是油橄榄果潜在的抗炭疽病生防菌。其中Fusicollaviolacea被研究证明对引发猕猴桃软腐病的交链孢霉属具有拮抗作用,有一定生防效果[34]。
本研究中,部分真菌对炭疽病感染较敏感。果皮中假尾孢属和核盘菌属对炭疽病病原菌的响应最明显,为炭疽病病原菌敏感菌。LEFse分析结果显示,假尾孢属和核盘菌属为健康果皮差异的指示菌群。在果肉中,发病后黑孢霉属和短梗霉属两种优势菌属均明显降低,LEFse分析结果显示它们为健康果肉中的指示菌群之一。由此可见,假尾孢属和核盘菌属可作为油橄榄果炭疽病感染早期的指示菌,黑孢霉属和短梗霉属则可作为炭疽病病原菌入侵果肉的指示菌群。
4 结 论
本研究采用高通量测序技术,分析了感染炭疽病对油橄榄果真菌群落结构的影响。研究得出:①炭疽病感染显著改变了油橄榄果果皮和果实中真菌群落的结构。感染炭疽病后,油橄榄果真菌丰富度指数变化不显著,但各分类水平上各类群间的相对丰富度明显改变,群落的多样性显著降低,表明感染炭疽病后,由于病原菌数量的增长,改变了菌群间个体数量的分布,降低了真菌群落多样性;②果皮中的假尾孢属和核盘菌属,果肉中的黑孢霉属和短梗霉属为炭疽病病原菌敏感菌,可作为油橄榄果炭疽病感染早期的指示菌;③染病后期果皮上共生营养型真菌数量显著回升,可能是抗炭疽病生防菌菌群来源,可作进一步筛选;④油橄榄果感染炭疽病后期,除了炭疽病病原菌,果皮中其它病原菌菌属,如Bisifusarium属、汉纳酵母属、镰刀菌属在重病时期也明显增加,提示炭疽病防治需要针对这些菌群进行综合防治。
