短花针茅荒漠草原物种间关系的多度不对称性对载畜率的响应
2023-03-05郑佳华李雅男乔荠瑢赵天启赵萌莉
张 彬, 张 峰, 郑佳华, 王 琪, 李雅男, 乔荠瑢, 赵天启, 于 跃, 赵萌莉*
(1.内蒙古农业大学草原与资源环境学院, 内蒙古 呼和浩特 010019; 2.内蒙古农业大学草地资源教育部重点实验室, 内蒙古 呼和浩特 010019; 3. 巴林右旗森林草原保护发现中心, 内蒙古 赤峰 025150)
中图分类号:S812文献标识码:A
文章编号:1007-0435(2023)02-0510-08
Responses of Abundance-asymmetric of Interspecific Interaction to Stocking Rates in theStipabrevifloraDesert Steppe
ZHANG Bin1,2, ZHANG Feng1,2, ZHENG Jia-hua1,2, WANG Qi1,2, LI Ya-nan1,2, QIAO Ji-rong1,2, ZHAO Tian-qi1,2, YU Yue3, ZHAO Meng-li1,2*
(1.College of Grassland, Resource and Environment, Inner Mongolia Agricultural University, Hohhot, Inner Mongolia Autonomous Region 010019, China; 2. Key Laboratory of Grassland Resources, Ministry of Education of China, Inner Mongolia Agricultural University, Hohhot, Inner Mongolia Autonomous Region 010019, China; 3. Forest Grassland Protection and Discovery Center in Bairin Right Banner, Chifeng, Inner Mongolia Autonomous Region 025150, China)
Abstract:The relationship between the relative abundance of species and its interspecific relationship affecting the coexistence of species is unclear. We conducted a grazing experiment to test abundance-asymmetric of interspecific interaction in the plant communities and its responses to different stocking rates including the control,light grazing,moderate grazing and heavy grazing in theStipabrevifloradesert steppe in Inner Mongolia. Taking the plant communities ofS.brevifloradesert steppe as the research objects,the abundance and frequency of all species in the communities under control,light grazing,moderate grazing and heavy grazing were analyzed. The results showed the numbers of species in the steppe had a decreasing trend with increasing stocking rates. The codistribution index of species pairs had a significant dependence on its abundance under all different stocking rates,and common species (e.g.,S.breviflora,C.songoricaandC.aristatum) had a generally stronger effect on the abundance of other species. The interaction intensity ofS.brevifloraonC.songoricaandC.aristatumwas higher than that ofC.songoricaandC.aristatumonS.brevifloraunder all different stocking rates,showing the existence of abundance-asymmetric of interspecific interaction. Besides,grazing increased the interaction intensity ofS.brevifloraonC.songorica,while decreased the interaction intensity ofC.songoricaonS.breviflora. As a summary,there was an abundance-asymmetric of interspecific interaction among species,and the livestock grazing further imposed a change on the intensity of interspecific interaction.
Keywords:Desert steppe;Grazing;Abundance-asymmetric;Interspecific interaction
植物群落中的各物种之间会竞争营养、光线和空间等资源,然而其利用资源的方式却存在很大差异[1]。植物群落通常物种组成丰富,但大多数物种个体较少,仅有少数物种个体较多[2-3],造成群落出现多度分布不均匀的格局。群落构建机制提出以来,较多生态学家从不同理论角度阐述、归纳总结了多样性的维持机制[4-6],试图揭示个体数较少的稀有种如何与个体数较多的常见种实现共存,如生态位和中性理论[7-9]。其中,群落构建过程中相关理论机制对于了解常见种与稀有种间的长期共存至关重要,如同种负密度制约效应[10-12]、种间关系的“随机稀释”假说[13]。“多度不对称假说”认为群落中植物个体与个体之间发生相互的作用均等,则会造成种间关系与物种多度相关[14];个体数较多的常见种间更容易检测到物种间的相互作用[15]。种间关系与物种多度显著正相关[16]。虽然较多研究从不同理论、角度支持该假说,但较少有研究对其进行相关验证。
放牧是荒漠草原主要利用方式之一[17]。荒漠草原作为我国北方天然生态屏障,但是过度放牧造成其不同程度的退化[18]。以往的研究中,较多研究从群落结构[19]、种群空间分布[20]及稳定性[21]等角度对放牧干扰下植被群落进行相关研究,也有较多研究试图通过种间关联[19]、物种间空间分布关系[22]、灌木-草本关系[23]等角度研究群落物种间关系,但并未能揭示物种间关系的不对称性。与稀有种相比,常见种种群个体数较多,更易与其他常见种或稀有种相遇[3],从而造成常见种对其他物种的影响强度偏大;而稀有种由于种群个体数相对较少而难以与其他物种相遇,而造成稀有种对常见种或稀有种均影响较小,由此造成物种间关系强度不对称。植物群落内物种的相对多度与其种间关系存在一定的内部联系。然而,这种关系如何进一步影响群落内物种共存仍不清晰;故本文试图以短花针茅(Stipabreviflora)荒漠草原植被群落为研究对象,通过分析其群落出现物种的种群多度变化,试图检测物种间关系的多度不对称性,并试图揭示其物种关系的多度不对称性对不同载畜率的响应,为草地合理利用提供一定的理论基础。
1 材料与方法
1.1 研究区概况
本试验区位于内蒙古农牧业科学院综合试验示范中心四子王基地(41°46′44″ N,111°53′42″ E),海拔约1 456 m,典型中温带大陆性季风气候。年均温3.4℃,年均降水量约为250 mm。植被主要由多年生丛生禾草、杂类草和灌木、半灌木等组成,建群种为短花针茅(S.breviflora),主要优势种为无芒隐子草(Cleistogenessongorica)和冷蒿(Artemisiafrigida),其他常见植物还有银灰旋花(Convolvulusammanni)、刺穗藜(Chenopodumaristatum)和冷蒿(Artimisiafrigida)等。植被稀疏低矮,群落平均高度为8 cm左右;土壤较贫瘠,主要为淡栗钙土;土壤有机质含量较低,具有缺磷、少氮、钾有余的特点。试验区于2004年进行围建,在设立之前,该地区主要为放牧场,全年放牧绵羊且放牧强度相对较高;植被覆盖率为17%~20%,平均植物高度为8 cm。在长期放牧过程中(2004—2020年间),随着放牧强度增大,群落中物种数逐渐降低,主要表现为银灰旋花、冷蒿等多年生杂类草,及篦齿蒿(Neopallasiapetinata)、灰绿藜(Chenopodiumglaucum)等一、二年生植物物种的减少;主要原因在于放牧家畜的选择性采食性;相比于多年生丛生禾草,多年生杂类草及一、二年生植物适口性整体较高。
1.2 试验设计及取样
试验区于2004年进行围建,设置对照区(Control,CK)、轻度放牧(Light grazing,LG)、中度放牧(Moderate grazing,MG)和重度放牧(Heavy grazing,HG) 4个放牧强度,各3次重复,共12个小区(4.4 公顷·小区-1)。放牧家畜为当地成年蒙古羯绵羊,载畜率分别为0(CK),1.80(LG),3.60(MG)和5.40(HG)羊单位·hm-2·a-1;对照区、轻度、中度及重度放牧区放牧绵羊数分别为0,4,8和12只。自2004年起进行连续放牧,每年放牧期于6月初开始并于11月底结束。
本试验于2020年8月生物量高峰期时,在各小区均设置10条样带,每条样带各设置10个0.5 m×0.5 m样方,共计1 200个样方。记录各样方中所有出现物种的多度(密度),即单位面积内出现的同种个体的数量;其次,通过调查的样方物种数据计算各物种的出现频率,即物种出现的样方数与总样方数之比的百分率。
1.3 物种的同分布指数
CO1,2=O1,2/O1
(1)
其中,O1,2代表两个物种同时出现的样方数,O1代表只有物种1出现的样方数。如果CO1,2值很大,说明物种2在物种1出现的大多数样方都出现,物种1受到物种2影响的可能性比较大;如果CO1,2值很小,说明在物种1分布的大多数样方内,都没有物种2个体,二者发生作用的可能性很小。
1.4 数据处理
基于所有1 200个0.5 m×0.5 m样方对植被群落各种群多度(密度)及频度原始数据进行初步整理后统计其物种丰富度,筛选出不同载畜率下的有效物种,即各处理下筛选出种群个体数大于10株的物种。其次,筛选出各载畜率水平下共有物种,根据种群多度进行降序排列;并通过计算各物种的同分布指数结合种群多度进行气泡图制作。所有图表在SigmaPlot 14.0制作。
2 结果与分析
2.1 物种多度及出现频率
从图1可以看出,对试验区1 200个样方调查发现,共出现48种植物,各物种多度差异较大,在1~45 743株之间,合计85 226株。其中,刺穗藜种群多度最高,共45 743株;个体数在1 000株以上的物种有8种,分别为刺穗藜、无芒隐子草和短花针茅等;而个体数低于10株的物种有12种,分别为细叶鸢尾(Iristenuifolia)、扁蓿豆(Melilotoidesruthenica)和点地梅(Androsaceumbellata)等。对群落调查发现,个体数较多的物种相对较少,而个体数较少的物种相对较多。
从图2可以看出,样方调查出现的48种植物中,各物种出现频率差异较大,在0.08%~96.92%之间。其中,短花针茅种群出现频率最高,为96.92%,无芒隐子草次之,为92.72%。仅有7种植物出现频率大于10%,分别为短花针茅、无芒隐子草和刺穗藜等;而出现频率低于1%的物种有16种,分别为达乌里芯芭(Cymbariadahurica)、砂珍棘豆(Oxytropisgracilima)和细叶鸢尾等。对群落调查发现,出现频率高的物种相对较少,而出现频率低的物种相对较多。
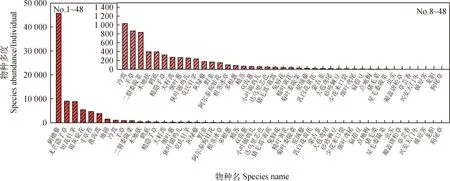
图1 试验区出现的所有物种及其多度Fig.1 The appearance and abundance of the all species in the experimental sites
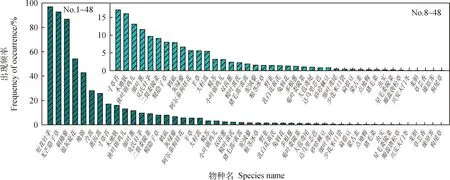
图2 试验区各物种的频率Fig.2 The occurrence frequency of the species in the experimental sites
2.2 不同载畜率下有效物种的物种多度
从图3可以看出,不同载畜率下有效物种(个体数大于10株)存在一定差异,CK,LG,MG及HG的有效物种数分别为30,29,18和12种;随着载畜率增大,有效物种数呈现出降低的趋势。此外,不同载畜率下物种多度排序发生变化,CK和LG下主要以刺穗藜、银灰旋花和短花针茅为主,MG下主要以刺穗藜、无芒隐子草和短花针茅为主,而HG主要物种与MG相同,但排序发生变化。再者,不同载畜率下刺穗藜种群多度差异较大,CK,LG,MG及HG下刺穗藜种群多度分别为3 738,9 037,19 522和13 446株。

图3 不同载畜率下有效物种的物种多度Fig.3 The abundance of effective species under the different stocking rates
2.3 不同载畜率下群落物种同分布指数
从图4可以看出,对不同载畜率下共同出现的11种植物进行两两物种对的同分布指数分析发现,不同载畜率下同分布指数CO均存在明显的多度依赖性,总体呈现出“左小右大”的结构,表明CO值的大小主要受物种2(X轴物种)多度的影响。此外,当物种2(X轴物种)较小时,无论物种1(Y轴物种)个体数多或少,CO值大多较小,表示稀有种对其他物种的作用可能性普遍很小;表明稀有种之间很难相遇,而常见种与常见种之间相遇的概率比较大。再者,不同载畜率下物种对的同分布指数存在一定差异,但各载畜率下主要表现为刺穗藜、短花针茅和无芒隐子草(X轴物种)对其他物种表现出较强的种间关系。
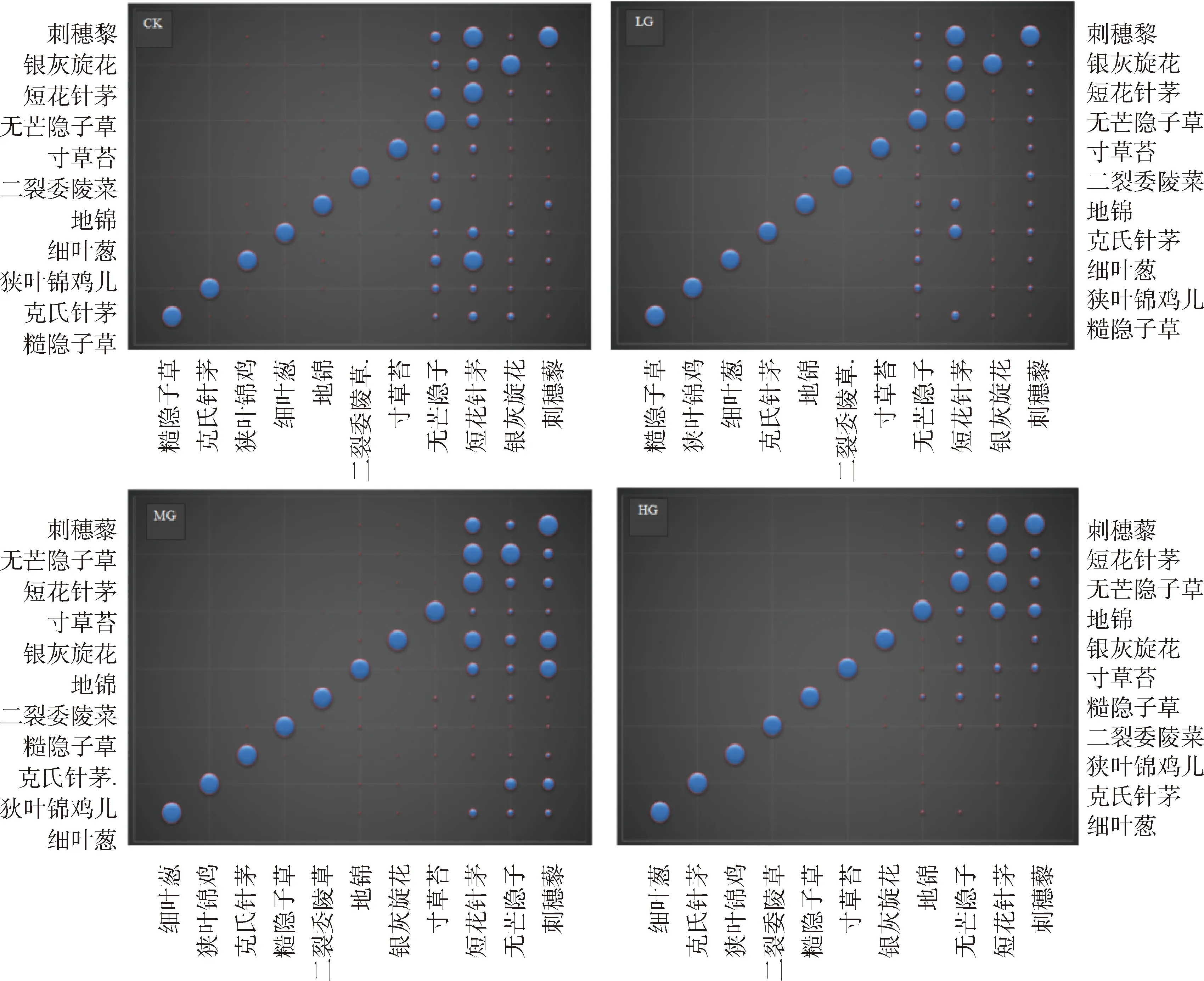
图4 不同载畜率下物种同分布指数与多度的关系Fig. 4 The relationship between codistribution index and species abundance under the different stocking rates注:横、纵坐标物种多度均从坐标原点向两轴逐渐增加;图中气泡表示物种之间的同分布指数,其气泡越大表明其作用关系越强Note:The species abundance in horizontal or vertical coordinate increases gradually from the origin along the coordinate. The individual bubbles represent the co-distribution index between two species. The larger the bubble is,the stronger the interaction relationship between two species exists
2.4 不同载畜率下主要物种对的同分布指数
从图5可以看出,对不同载畜率下主要物种对的同分布指数进行分析表明,CK和HG下短花针茅对刺穗藜表现出很强的作用强度,其次则是对无芒隐子草表现出较强的作用强度;而LG和MG下短花针茅对无芒隐子草表现出很强的作用强度,其次则是对刺穗藜表现出较强的作用强度。此外,CK和LG下短花针茅还对银灰旋花表现出较强的作用强度,而HG下短花针茅则是对地锦表现出较强的作用强度。主要物种对的同分布指数表明,优势种对其他物种的作用强度普遍较强(如短花针茅、无芒隐子草及刺穗藜)。
从图6可以看出,不同载畜率下短花针茅对无芒隐子草及刺穗藜的作用强度要高于无芒隐子草和刺穗藜对短花针茅的作用强度,表现出物种间作用强度存在不对称性。此外,放牧使得短花针茅对无芒隐子草的作用强度呈现出增大的趋势,使得无芒隐子草对短花针茅的作用强度呈现出降低的趋势;而MG使得刺穗藜对短花针茅的作用强度呈现出增大的趋势。

图5 不同载畜率下主要物种对的同分布指数Fig.5 Codistribution index of major species pairs under the different stocking rates
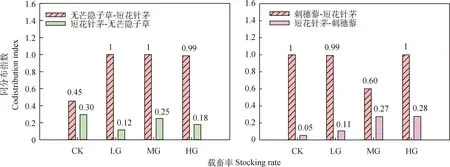
图6 不同载畜率下短花针茅-无芒隐子草及短花针茅-刺穗藜物种对的同分布指数Fig.6 Codistribution index of species pairs between Stipa breviflora-Cleistogenes songorica and Stipa breviflora-Chenopodium aristatum under the different stocking rates
3 讨论
3.1 不同载畜率对物种出现频率及多度的影响
在长期放牧干扰下,草地群落结构发生改变,竞争力较弱的优良牧草易被竞争力较强的杂类草替代,从而造成群落格局发生变化[24]。此外,由于营养、光线和空间等资源的限制,植物群落中各物种之间会存在一定的竞争[1]。由于植物群落中的各物种种群的个体数不同,物种关系之间可能存在一定的多度依赖性[3]。本研究表明,植物群落中个体数较多的物种相对较少,而个体数较少的物种相对较多;出现频率高的物种相对较少,而出现频率低的物种相对较多。有研究表明,由于家畜的选择性采食作用,放牧会造成适口性高的物种的种群多度降低[25]。也有研究表明,放牧过程中,家畜选择性采食、践踏及游走等牧食行为易造成种群出现破碎化,从而造成丛生禾草植物多度增加[26]。此外,放牧通过采食植株地上部分降低其顶端优势,从而促进丛生禾草分蘖,也可造成其种群多度增加[27]。个体数较多的常见种与其他物种的相遇较大,容易受到其他物种较强的排斥作用,从而在一定程度上限制其种群增长,故造成个体数较多的物种偏少[3]。稀有种与其他物种的相遇概率较小,虽然缺乏竞争优势,但可以通过种群聚集分布而在一定程度上达到局部优势,从而达到与个体数较多的常见种共存;但局部共存的种群聚集分布方式与物种繁殖方式、种子扩散方式、传播距离或环境异质性有关[3]。本研究中,有些物种虽然种群个体数较多,但并未有较多的种群出现频率,可能是由于物种个体数较少而自身生存策略上的聚集分布已达到与其他物种的局部共存,主要是由于种群分布格局会造成其种内及种间竞争能力[28]。有研究表明,物种若具有较强的聚集性,则会具有较强种内竞争力,而聚集性较弱的物种则具有较强的种间竞争力[29];而稀有种更倾向于聚集分布[30-31]。
3.2 不同载畜率对物种间相互作用强度的影响
种间关系是植物群落重要的数量与结构特征,是形成植物群落结构和空间格局的主要动力[26]。本研究表明,物种间相互作用的强度具有多度依赖性;当某个物种的个体数较少时,无论其它物种的个体数多或少,此物种(个体数较少的物种)与其它物种的同分布指数均较小(即物种间的相互作用强度较小),这说明稀有种对其他物种的作用可能性普遍较小;常见种(如短花针茅、无芒隐子草及刺穗藜)对其他物种的作用强度普遍较强;这与毛子昆等[3]对长白山阔叶红松林进行的研究一致;其研究认为物种的聚集分布可能造成种间关系的多度不对称性,主要是由于稀有种的聚集造成其种群分布较小,而与其他物种形成空间隔离[32-33];而常见种由于分布范围广而更容易与其他物种作用而造成种间关系的多度不对称性。在本研究中,短花针茅、无芒隐子草及刺穗藜等常见种种群个体数较多,其出现频率也相对较高,也表明其种群分布较广,容易与其他常见种或稀有种相遇,故对其他物种的作用强度偏高。
放牧会改变物种的种群多度及物种间的关系[22]。有研究表明,放牧可以通过改变植物冠层高度造成植物截获光照的能力发生变化,从而改变种间关系[34];家畜的选择性采食作用通过降低适口性高的物种的种群多度也可以改变种间关系[25]。还有研究表明,物种间相互作用的能力取决于物种的生活习性、生活型和生态幅度等,放牧可以通过改变物种的空间异质性来影响种间关系[35-36]。此外,放牧可能造成物种间的关系强度发生变化。有研究表明,围封和轻度放牧处理下植物群落整体表现为竞争关系,而中度和重度放牧处理下群落整体呈亲和关系[19]。也有研究表明,放牧干扰导致植物种群种内竞争强度增加,种间竞争强度减弱;不同放牧处理下物种间亲和关系与竞争关系均存在不对称性[37]。本研究表明,随着载畜率增大,物种数呈现出降低的趋势且物种多度排序发生变化,CK和LG下主要以刺穗藜、银灰旋花和短花针茅为主,MG和HG下则主要以刺穗藜、无芒隐子草和短花针茅为主。这可能与家畜的选择性采食有关[38-40];相比于短花针茅和无芒隐子草等多年生丛生禾草,家畜更喜采食银灰旋花等多年生杂类草。此外,本研究还表明,不同载畜率下,短花针茅对无芒隐子草及刺穗藜的作用强度要高于无芒隐子草和刺穗藜对短花针茅的作用强度,表现出物种间作用强度存在不对称性。这主要是由于短花针茅种群出现频率很高,当无芒隐子草或刺穗藜出现时,总会与短花针茅相遇,而受到短花针茅较强的作用;而短花针茅出现时,并非总是与无芒隐子草或刺穗藜相遇,而受到无芒隐子草或刺穗藜的作用相对较小。此外,放牧使得短花针茅对无芒隐子草的作用强度呈现出增大的趋势,使得无芒隐子草对短花针茅的作用强度呈现出降低的趋势;这主要是因为放牧造成短花针茅优势度增大,而无芒隐子草优势度降低;随着载畜率增大,短花针茅与无芒隐子草相遇的概率减小,而无芒隐子草与短花针茅相遇的增大。
4 结论
植物群落中个体数较多的物种相对较少,而个体数较少的物种相对较多;出现频率高的物种相对较少,而出现频率低的物种相对较多。不同载畜率下,同分布指数CO均存在明显的多度依赖性,常见种(如短花针茅、无芒隐子草及刺穗藜)对其他物种的作用强度普遍较强。此外,不同载畜率下均表现出物种间作用强度的不对称性,短花针茅对无芒隐子草及刺穗藜的作用强度要高于无芒隐子草和刺穗藜对短花针茅的作用强度。放牧使得短花针茅对无芒隐子草的作用强度呈现出增大的趋势,使得无芒隐子草对短花针茅的作用强度呈现出降低的趋势;而中度放牧使得刺穗藜对短花针茅的作用强度呈现出增大的趋势。
