包被胍基乙酸对奶牛泌乳性能和瘤胃发酵的影响
2023-02-23刘永嘉霍文婕裴彩霞
■刘永嘉 郭 刚 霍文婕 张 静 裴彩霞 刘 强
(山西农业大学动物科学学院,山西太谷 030801)
胍基乙酸(GA)作为肌酸合成的前体物质,由甘氨酸和精氨酸转化而成[1]。肌酸是组织与细胞的能量缓冲剂,ADP接受磷酸肌酸的磷酸基团转化为ATP[1]。动物自身合成的肌酸只能满足需求量的50%[2],外源补充肌酸能改善动物体的能量状态,促进细胞生长和蛋白质合成[3]。与肌酸相比,GA性质稳定、价格低、易吸收,对提高组织和细胞中肌酸的含量与能量的供应更有效[1],因此,被作为营养性添加剂补充动物内源肌酸合成的不足。研究证实,猪或鸡的日粮中添加GA,生长性能提高,饲料利用率改善,肝脏、肌肉和血液中肌酸浓度增加[4-5]。日粮中添加GA,滩羊生长性能和屠宰性能改善[6];安格斯公牛平均日增重与血液肌酸浓度提高[7]。目前,GA对奶牛泌乳性能影响的研究还未见报道。另外,补充GA,公牛瘤胃总挥发性脂肪酸浓度、总细菌、真菌和纤维分解菌数量增加[7]。但是,Speer[8]发现,补充的GA约50%在瘤胃中被降解,与瘤胃灌注相比,皱胃灌注GA 对提高血液肌酸浓度更有效。包被GA(CGA)添加剂在瘤胃中的降解率低,能使添加的GA最大限度到达小肠被吸收。基于现有报道,试验探究补充CGA 对奶牛产奶性能、养分消化和瘤胃菌群的影响,为GA 用于反刍动物生产提供理论支撑。
1 材料与方法
1.1 试验动物与试验设计
泌乳天数(111.2±9.6)d、产奶量(29.9±1.8)kg/d和胎次(2.2±0.8)的48 头荷斯坦奶牛,依据随机区组试验设计分为:对照组、LCGA、MCGA 和HCGA 组,分别以CGA的形式在基础日粮中添加0、0.3、0.6 g GA/kg和0.9 g GA/kg 干物质(DM)。试验期为80 d,其中预试期20 d,正试期60 d。
1.2 试验日粮与饲养管理
试验日粮依据NRC(2001)[9]配制(见表1),试验用GA 由河北盘恒科技有限公司提供,饲料级,纯度98.3%,CGA 依据Wang 等[10]的方法制备,过瘤胃率85.3%。试验牛单栏饲养、自由饮水和采食,每天挤奶两次(05:00 和17:00),饲喂两次(08:30 和20:30)。晨饲时,先将CGA 与少量日粮混合,待牛全部采食完毕后,再饲喂其余日粮。
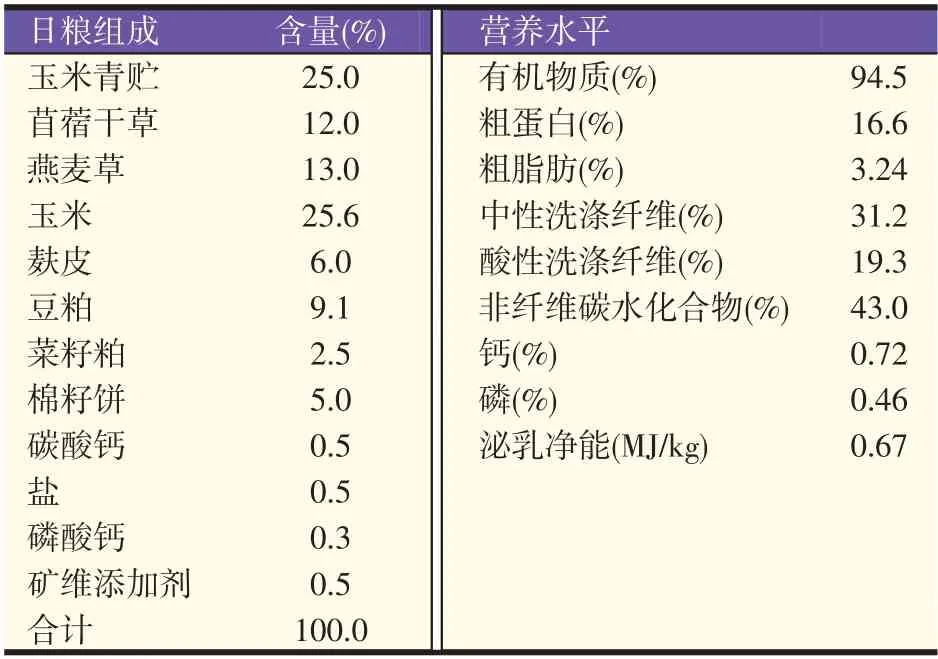
表1 日粮组成和营养水平(干物质基础)
1.3 样品采集与分析
1.3.1 饲料样和粪样
正试期间,计算每头试验动物每天的干物质采食量(DMI)(喂料量与剩料量之差)。第51~58 天,采集每头牛每天所喂的饲料和剩料样品,-20 ℃保存。每天06:00、12:00、18:00 和24:00,通过直肠采集每头牛粪便样品约200 g,-20 ℃保存。试验结束后,粪样和饲料样按比例以牛为单位混匀,65 ℃烘至恒重,粉碎(1 mm),依据实验室常规分析法[11]测定样品中DM、粗灰分、蛋白质(CP)、脂肪、酸性洗涤纤维(ADF)、中性洗涤纤维(NDF)和酸不溶灰分含量,同时计算非纤维碳水化合物(NFC)的含量(100-CP-脂肪-NDF-灰分)。
1.3.2 产奶量和奶样
正试期间,记录每头试验牛每天产奶量。每隔7 d,采集当天两次奶样(100 mL),依据产奶量以牛为单位混合均匀,装入加重铬酸钾防腐剂的奶样瓶中,测定样品中脂肪、乳糖和蛋白质[11]。
1.3.3 瘤胃液
正式试验期的第59天和第60天,早晨饲喂后3 h,通过口腔,用胃管采集每头牛瘤胃液样品(150 mL),立即用PHS-3CPH数显酸度计测定样品pH。然后,用四层医用纱布过滤样品,分两份(10 mL),-20 ℃保存,用于挥发性脂肪酸(VFA)含量和菌群数量的测定[11]。
1.4 数据处理与统计分析
试验数据通过统计分析软件SAS 9.0 的one-way ANOVA 模型进行方差分析和Duncan’s 法进行多重比较[12]。
2 结果与分析
2.1 采食量、泌乳性能和饲料效率
由表2 可知,各试验组奶牛的DMI 差异不显著(P>0.05);实际产奶量和乳脂矫正奶产量,MCGA 组和HCGA 组最高,其次是LCGA 组,对照组显著低于CGA 添加组(P<0.05);乳糖产量,MCGA 组和HCGA组显著高于对照组(P<0.05);乳脂肪和乳蛋白质产量,CGA添加组显著高于对照组(P<0.05);饲料效率,HCGA组最高,其次是MCGA组和LCGA组,对照组显著低于CGA添加组(P<0.05)。

表2 包被胍基乙酸对奶牛产奶性能的影响
2.2 日粮养分表观消化率
由表3可知,日粮DM表观消化率,MCGA和HCGA组最高,LCGA 组次之,对照组最低(P<0.05);CGA 添加组有机物(OM)表观消化率显著高于对照组(P<0.05);MCGA组CP表观消化率显著高于其余3组(P<0.05);MCGA 组和HCGA 组NDF 和ADF 表观消化率显著高于对照组和LCGA 组(P<0.05);MCGA 组和HCGA组NFC表观消化率显著高于对照组(P<0.05)。

表3 包被胍基乙酸对奶牛日粮养分表观消化率的影响(%)
2.3 瘤胃发酵
由表4可知,HCGA组和MCGA组,瘤胃液pH、乙酸摩尔比和乙酸/丙酸显著低于对照组和LCGA 组(P<0.05),瘤胃总VFA浓度和丙酸摩尔比显著高于对照组和LCGA 组(P<0.05);各试验组之间,瘤胃中丁酸、异丁酸、戊酸和异戊酸摩尔比与氨态氮浓度差异不显著(P>0.05)。

表4 包被胍基乙酸对奶牛瘤胃发酵的影响
2.4 瘤胃微生物数量
由表5 可知,HCGA 组和MCGA 组,瘤胃液总细菌、总原虫和溶纤维丁酸弧菌数量显著高于对照组和LCGA 组(P<0.05);总厌氧真菌、黄色瘤胃球菌、白色瘤胃球菌、栖瘤胃普雷沃氏菌、嗜淀粉瘤胃杆菌和产琥珀酸丝状杆菌数量,MCGA 组和HCGA 组最高,LCGA组次之,对照组最低(P<0.05)。
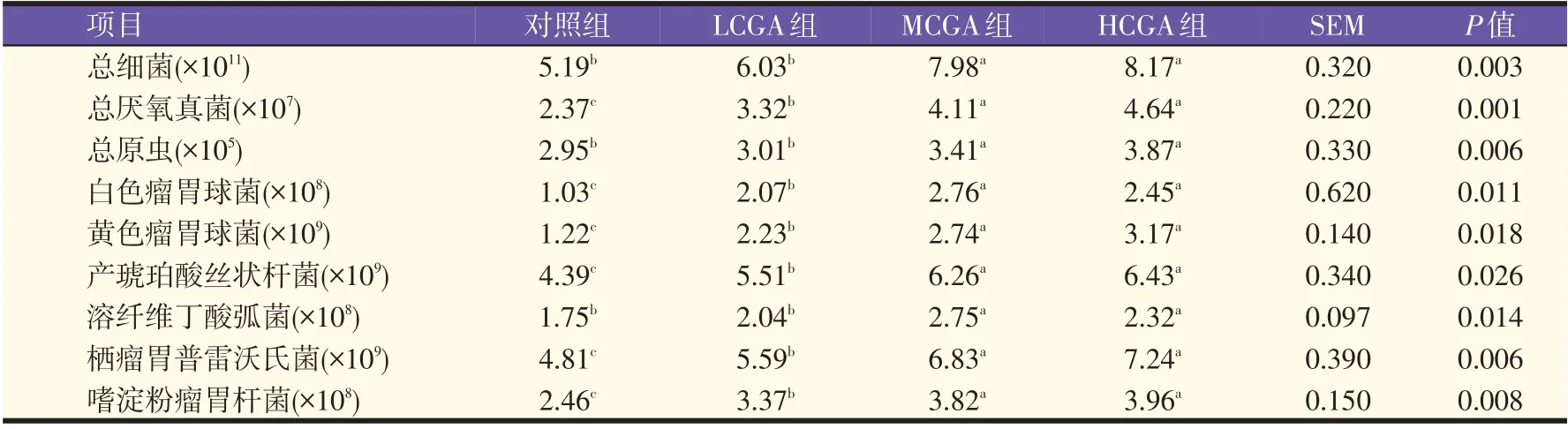
表5 包被胍基乙酸对奶牛瘤胃微生物数量的影响(拷贝数/mL)
3 讨论
奶牛日粮补充CGA 对DMI 的影响不显著。同样,Speer[8]报道,肉牛瘤胃灌注GA 10 g/d 或20 g/d 对DMI 的影响不显著。但是,Li 等[7]报道,添加GA 0.3、0.6 g/kg和0.9 g/kg DM,安格斯公牛的DMI线性提高。血液肌酸浓度对动物采食量有重要的调控作用[13],因此,试验结果的差异可能与试验动物肌酸状态的不同有关。泌乳牛乳脂矫正奶产量和实际产奶量的增加,与瘤胃总VFA浓度和日粮养分消化率的提高有关,说明补充CGA 改善了奶牛对养分的利用效率,表现为增加的饲料效率。其他研究同样发现,添加GA 提高了滩羊或安格斯公牛的平均日增重和饲料效率[6-7]。日粮补充CGA后,泌乳牛乳脂肪产量的提高与瘤胃乙酸浓度提高的结果一致。瘤胃乙酸浓度为总VFA浓度和乙酸摩尔比的乘积,对照组、LCGA、MCGA和HCGA分别为:77.9、80.6、83.3 mol/mL 和85.9 mol/mL。乙酸是奶牛乳脂合成的主要前体物质,添加乙酸,牛乳腺上皮细胞参与乳脂合成的关键基因表达上调,乳脂产量提高[14-15]。乳蛋白产量的提高与添加CGA改善了奶牛肌酸的供应有关。肌酸是组织与细胞的能量缓冲剂[1],能促进细胞生长和蛋白质合成[3]。研究发现,日粮添加GA,安格斯公牛血液和肝脏中肌酸浓度提高[16]。添加CGA后,奶牛乳糖产量的提高,与瘤胃总VFA和丙酸摩尔比的提高有关。血液中的葡萄糖是乳糖的主要来源,而糖异生的主要前体物质是丙酸,奶牛的葡萄糖供应约80%来源于丙酸的糖异生[17]。
全消化道DM 和OM 消化率增加,与瘤胃总VFA浓度增加的结果相一致,表明添加CGA 提高了养分在瘤胃的降解。另外,CGA的添加对养分在小肠的消化也有促进作用。研究发现,肌酸是肠道消化液分泌、细胞更新和养分吸收的必需营养因子,补充GA能改善动物肠黏膜形态[18]。Li 等[7]在公牛日粮中添加GA,同样发现,日粮养分消化率增加。日粮NDF 和ADF 消化率的提高与瘤胃乙酸浓度提高的结果相一致,归因于瘤胃真菌、主要纤维分解菌和原虫数量的增加。反刍动物瘤胃中,黄色瘤胃球菌、产琥珀酸丝状杆菌、溶纤维丁酸弧菌和白色瘤胃球菌是最重要的纤维分解细菌,这4类细菌与真菌和原虫通过分泌纤维分解酶,降解饲料纤维类物质为乙酸[17]。添加CGA后,全消化道NFC消化率的增加与瘤胃丙酸摩尔比的增加相一致,与栖瘤胃普雷沃氏菌、嗜淀粉瘤胃杆菌和溶纤维丁酸弧菌数量的增加有关。这3 种细菌通过分泌淀粉酶,降解饲料中NFC为丙酸[17]。
瘤胃液pH 与总VFA 浓度呈负相关[17]。因此,瘤胃pH的降低归因于补充CGA后总VFA浓度的提高,与Li 等[7]的报道一致。试验结果表明,泌乳牛日粮补充CGA 刺激了瘤胃微生物的增殖,表现为总真菌、细菌和原虫数量的提高。肌酸是微生物细胞能量代谢的必需成分,而且,细菌氧化GA 的过程能产生ATP[19]。瘤胃乙酸摩尔比的降低和丙酸摩尔比的增加导致乙酸/丙酸值降低,表明添加CGA 使瘤胃发酵类型转化为产生更多的丙酸。瘤胃氨态氮浓度无显著变化的结果与全消化道CP 消化率和瘤胃蛋白质分解菌(栖瘤胃普雷沃氏菌、嗜淀粉瘤胃杆菌和溶纤维丁酸弧菌)数量的提高不一致,可能是CGA 的添加促进了微生物蛋白质合成。瘤胃中的氨态氮主要为微生物降解蛋白质的产物,细菌(尤其是纤维分解菌)利用氨态氮作为氮源,VFA作为碳架合成微生物蛋白质[17]。瘤胃总VFA 浓度和纤维分解菌数量的提高,表明添加CGA可能促进了微生物蛋白质的合成。
4 结论
日粮添加CGA改善了奶牛产奶性能、养分消化和瘤胃发酵;CGA的最佳添加水平为0.6 g GA/kg DM。
