植物生长发育和逆境响应中SR-like蛋白的研究进展
2023-02-21张源珊陈冠群郭祥鑫唐倩雯李清韵申晓辉
张源珊,陈冠群,郭祥鑫,唐倩雯,李清韵,申晓辉
(上海交通大学 设计学院,上海 200240)
可变剪接又称选择性剪接(alternative splicing,AS),是指基因的初始转录产物前体mRNA(pre-mRNA)通过连接不同的剪接位点产生两种或多种成熟mRNA的过程,普遍存在于生命活动中,参与生物的生长发育过程。
植物中有42%~83%的断裂基因通过可变剪接产生多种剪接变异体[1-3]。根据可变剪接引起的变化结果可将剪接类型分为8种:1)外显子跳跃(exon skipping,ES);2)内含子保留(retained intron,RI);3)5′ 位点可变剪接(alternative 5′ splice sites,A5SS);4)3′ 位点可变剪接(alternative 3′ splice sites,A3SS);5)外显子互斥(mutually exclusive exons,WXE);6)第一个外显子可变(alternative first exon,AFE);7)最后一个外显子可变(alternative last exon,ALE);8)串联3′-端非编码区(tandem 3′-UTRs)[4]。在植物中,内含子保留是较为常见的可变剪接方式。
Pre-mRNA的剪接是剪接体以一种循序渐进的方式进行组装并作用到pre-mRNA上。剪接体的主要类型包括5个富含尿苷的小核核糖核蛋白(small nuclear ribonucleoproteins,U1、U2、U4、U5和U6 snRNPs)和多种非小核核糖核蛋白(non-small nuclear ribonucleoprotein,non-snSNPs)[5-6],其中,富含丝氨酸-精氨酸蛋白(serine/arginine-rich proteins,SR proteins)是决定可变剪接效率和准确性的一个重要剪接因子(splicing factors,SFs)家族,可控制剪接位点的选择、引导拼接体组装,在剪接的执行和调控中都是必需的[7]。
1 SR-like蛋白亚家族特点与定位
1.1 SR-like蛋白亚家族成员的鉴定
SR蛋白是一类富含丝氨酸/精氨酸的剪接因子,属于non-snRNP剪接体蛋白家族[8-10],在高等生物中高度保守[11]。SR蛋白氨基末端(N端)具有1或2个RNA识别基序(RNA recognition motif,RRM),羧基末端(C端)含有高度磷酸化的丝氨酸/精氨酸结构域(serine/arginine-rich domain,SR domain)或精氨酸/丝氨酸结构域(arginine/serine-rich domain,RS domain)或丝氨酸/脯氨酸重复序列(serine/proline repeat,SP);此外,有一些SR蛋白还包含一些特殊的结构如锌指节/结构域(Zinc Knuckle,ZnK/ Zinc binding domain)。根据结构域的类型和数量,植物SR蛋白可分为7个亚家族:SC、SCL、RSZ、RS2Z、SR、RS和SR-like,其中SR、SC和RSZ亚家族分别是人类SRSF1、SRSF2和SRSF7的直接同源基因,而RS、SCL、RS2Z和SR-like亚家族的成员由于获得了新的特征而被认为是植物特有的。
SR-like亚家族的结构特殊(单个RRM两侧各有一个RS结构域),之前由于SR-like亚家族成员SR45这种非典型的结构域被排除在SR家族外[12]。但近年有研究基于RRM的高度保守性和广泛性,从古生物、细菌、真核生物和病毒中挖掘了约700个完整的蛋白质组,发现所有的典型SR蛋白都有一个古老的起源,拟南芥(Arabidopsisthaliana)SR45和SR45a分别是人类RNPS1和Tra2的同源物[13],SR45及哺乳动物同源RNPS1在功能和进化方面与SR蛋白密切相关;Tra2蛋白控制着双性生殖的可变剪接,在果蝇性别决定中起着关键作用[14]。现已将它们定义为SR蛋白的亚家族成员[10,15-19](图1,表1)。目前,在十字花科、豆科、禾本科、大戟科、山茶科和菊科等124种植物中均挖掘到了SR-like蛋白家族(表2,数据来自NCBI,https://www.ncbi.nlm.nih.gov/)。有花植物中的SR45蛋白普遍存在同源物,但藻类中不存在[20];另外,SR45和SR45a基因自身经过选择性剪接能产生转录副本,如拟南芥AtSR45AtSR45a、水稻(Oryzasativa)OsSR45OsSR45a等。

表1 拟南芥中7个SR蛋白亚家族成员Table 1 Seven subfamily members of SR protein in Arabidopsis thaliana
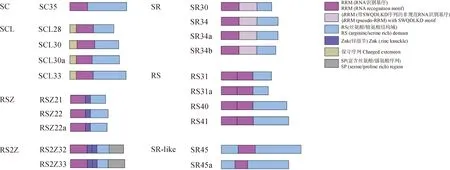
图1 拟南芥7个SR蛋白亚家族成员的结构示意图Fig.1 Structure of seven SR protein subfamilies in Arabidopsis thaliana
1.2 SR-like蛋白的结构特点
SR-like亚家族作为一类植物特异的SR蛋白,已被证明在体外异源互补分析中作为一个重要的剪接因子发挥作用[36]。SR-like亚家族包括SR45和SR45a成员。在拟南芥中,Sr45由3 379 个碱基组成,SR45蛋白由425个氨基酸组成;Sr45a由2 804个碱基组成,SR45a蛋白由382个氨基酸组成。第一个植物SR45蛋白是Golovkin等[9]使用酵母双杂技术钓取拟南芥U1-70K蛋白的互作蛋白时发现的。
如前文所述,SR-like的C端较其他SR蛋白多一个RS结构域,即单个RRM两侧有2个RS结构域(图1)。RRM结构域可以特异地识别并结合pre-mRNA来确定剪接位点,且偏向于结合外显子剪接增强子(exon splicing enhancers,ESE)序列,从而参与可变剪接[42-43];而RS结构域主要通过蛋白-蛋白之间的相互作用,募集其他剪接因子到RNA上完成剪接体的组装并进行剪接[44-46]。此外,RS域的磷酸化/脱磷酸化对蛋白的定位和移动起决定性作用,使得SR-like蛋白成为近10年SR蛋白研究中的热点。
1.3 SR-like蛋白的亚细胞定位
近几年,对SR-like蛋白的研究如火如荼,但对SR-like蛋白亚细胞定位研究涉猎的植物种类还不够广泛,有限的研究结果表明,SR-like蛋白一般定位在细胞核中。在烟草BY-2细胞中瞬时表达了绿色荧光蛋白(green fluorescent protein,GFP)和拟南芥SR45a蛋白融合载体[47],发现在DAPI染色的细胞核中检测到atSR45a-1a和atSR45a-2蛋白,表明拟南芥SR45a蛋白定位于细胞核。Zheng等[48]在分析低温胁迫下铁皮石斛(Dendrobiumcatenatum)的可变剪接事件转录组数据时,通过构建GFP载体进行瞬时表达检测帽结合复合物亚基帽结合蛋白 20(cap-binding protein 20,CBP20)和SR45a蛋白亚细胞定位发现,GFP荧光仅在细胞核中检测到,与在拟南芥中的亚细胞定位是一致的。用CELLO v2.5进行甘蓝型油菜(Brassicanapus)SR蛋白家族的亚细胞定位预测,6个SR45蛋白(BnaA06g11140D、BnaA08g23570D、BnaA09g56240D、BnaC05g12680D、BnaC08g16960D和BnaC08g38300D)均定位在细胞核内[17]。
SR蛋白是一种动态蛋白质,其定位与其分子功能直接相关。大多数SR蛋白在细胞核中都显示出类似的斑点定位[22,49],但SR-like蛋白的亚细胞定位与RS结构域有关。对拟南芥缺失突变体中的AtSR45a蛋白研究发现,RS1和 RS2域是核定位必需的信号区[50]。将烟草NtSR45蛋白的RS1域、RS2域及RRM域分别与GFP标签连接,发现含RS域(RS1或RS2域)的融合蛋白定位于细胞核,而RRM编码的蛋白则分散在细胞核和细胞质中,表明RRM域与SR45蛋白的定位无关,而RSl和RS2域都包含核定位信号,推测两个RS域可能存在功能冗余现象。RS1缺失蛋白定位于细胞核,而RS2缺失蛋白则分布于细胞核和细胞质中,进一步证明了RS2域对SR45定位于细胞核起决定性作用[51]。
SR45的核斑点分布受磷酸化和转录调控[20,37],并与5′剪接位点识别的关键蛋白U1-70K相互作用。为了研究SR45的定位和动态分布,在培养细胞和转基因拟南芥的间期核中发现,GFP-SR45存在于斑点状核和核质中,散斑表现为核内移动和形态变化;抑制转录和蛋白磷酸化导致SR45向更大的斑点移动,由转录抑制引起的斑点数目和形态的变化可被磷酸酶抑制剂阻断,表明细胞的转录活性和蛋白(去磷酸化)可以调节SR45的核内分布[20]。目前SR45a蛋白C端RS结构域中的磷酸化位点已被鉴定出来,并通过促分裂原活化蛋白激酶3(mitogen-activated protein kinase 3,MPK3)磷酸化[52]。
2 SR-like蛋白亚家族的调控方式和作用机制
早期的研究认为,可变剪接是介于转录和翻译之间的独立步骤,而后续的一系列研究则表明它与基因转录过程是偶联进行的,转录速率通过影响可变外显子剪接位点的选择调控可变剪接[53]。SR蛋白在剪接调控过程中的每一步都发挥着重要的作用,包括从剪切位点识别到最后阶段的剪接体组装[54]。然而,关于SR-like蛋白如何在这些过程中发挥作用的信息知之甚少。
目前研究表明SR-like蛋白参与了植物生长发育及逆境诱导反应的调控。在拟南芥、水稻、铁皮石斛、蔓菁(Brassicarapa)、木薯(Manihotesculenta)、烟草(Nicotianatabacum)、柳枝稷(Panicumvirgatum)、海枣(Phoenixdactylifera)、欧洲李(Prunusdulcis)、葡萄(Vitisvinifera)和玉米(Zeamays)等中发现,SR-like蛋白可通过自身或上、下游靶基因可变剪接转录后调控机制应答生长、发育和环境胁迫信号,进而影响植株大小、开花时间、器官形态学建成和生物/非生物胁迫[8,55-56](表2)。
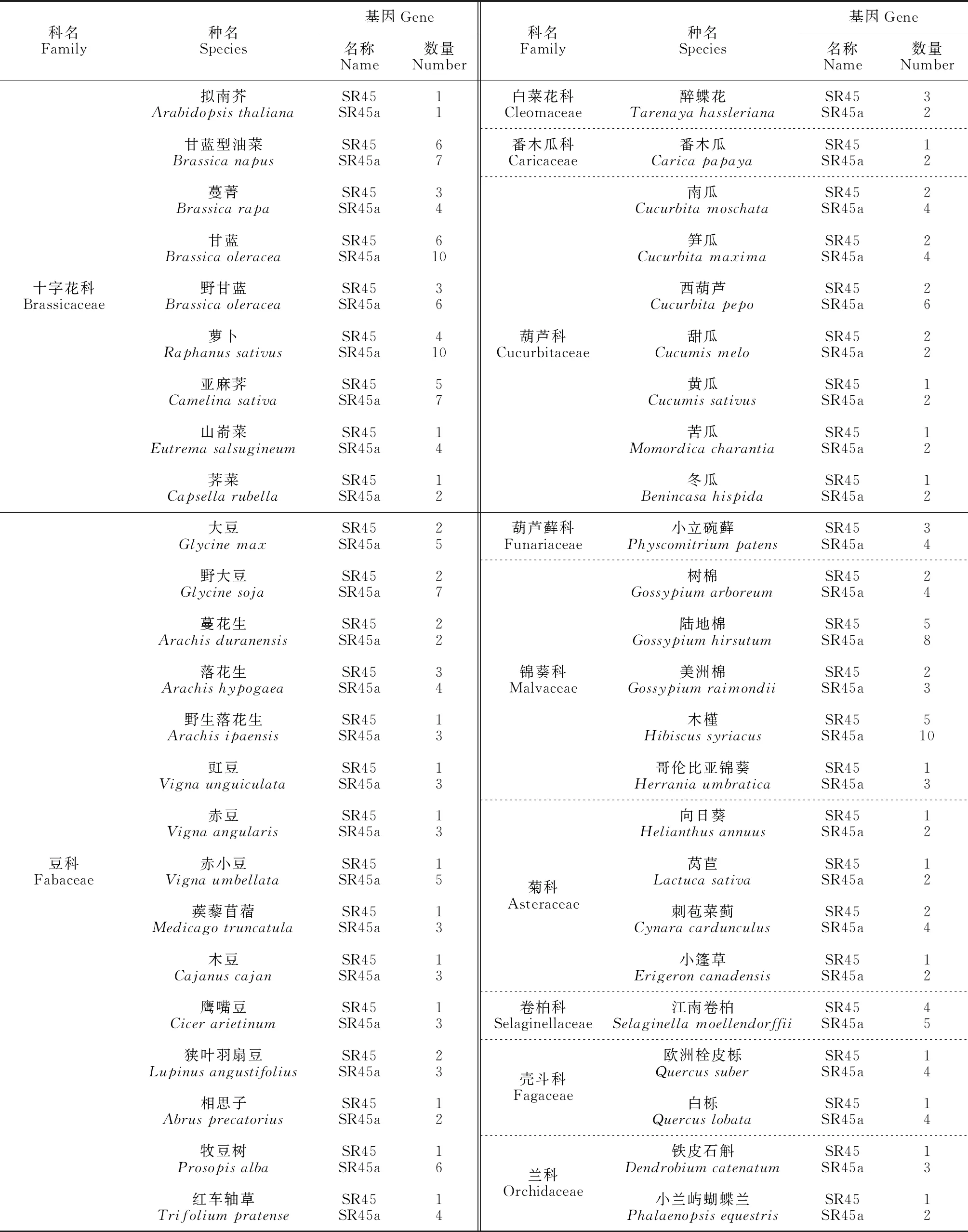
表2 从124种植物中鉴定出的SR-like蛋白亚家族成员及数量Table 2 Members and numbers of SR-like protein subfamily identified from 124 species of plants

续表2 Continued Table 2
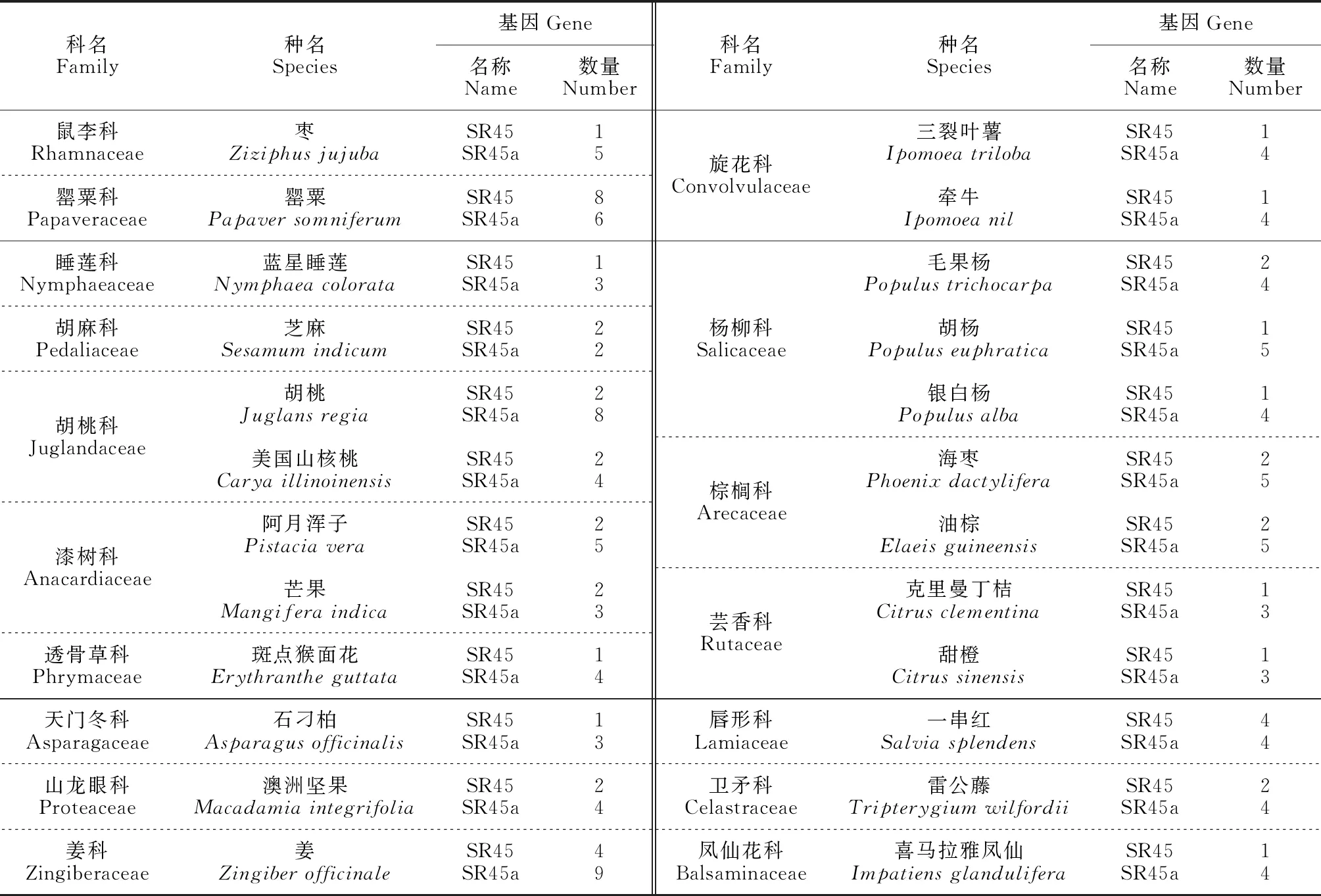
续表2 Continued Table 2
2.1 通过自身可变剪接调控下游靶基因转录
到目前为止,有关SR-like 蛋白可变剪接功能研究仅局限于模式植物拟南芥上。AtSR45可变剪接产生SR45.1(长亚型)和SR45.2(短亚型)2个转录本,在不同发育阶段(种子、幼苗和成苗)对热胁迫和盐胁迫有很高的敏感性。只有SR45.1是耐热性和耐盐性的正调节因子[57],并在花瓣发育中起主要作用;而SR45.2主要调控根的生长[56]。拟南芥SR45a自身也存在可变剪接,SR45a-1a编码的全长蛋白RRM结构域两侧为2个RS基序,SR45a-1b则编码了C端缺少RS结构域的截短蛋白;过表达SR45a-1a和SR45a-1b的拟南芥株系表现出盐敏感的表型[10]。由此可见,SR-like通过自身可变剪接调控下游靶基因转录。
2.2 通过蛋白互作及后修饰的调控
SR-like蛋白不仅可以和SR蛋白的其他亚家族成员相互作用(图2,A),还可以和剪接体蛋白或蛋白激酶之间互作,但目前已有研究仅局限于SR45(图2,B)。Stankovic等[30]通过酵母双杂分析表明,SR45的两个RS结构域和SR亚家族成员SR30、SR34和SR34a之间存在强烈的相互作用。Yan等[21]在分析拟南芥SR蛋白亚家族的突变体表型时发现,SR45与SCL33通过相互作用调节转录和剪接以维持基因的正确表达和发育功能。U1-70K是U1 snRNP特异的3种蛋白质之一,与pre-mRNAs的组成性剪接(constitutive splicing,CS)和可变剪接有关[58]。Golovkin等[9]发现了第一个SR45蛋白,并通过免疫共沉淀实验证实U1-70K和SCL33、SR45蛋白以及SR45和SCL33蛋白间发生相互作用。U2AF35是U2AF复合物的小亚基,参与识别3′剪接位点[59]。Day等[60]筛选了带有SR45的酵母双杂文库,通过体内和体外实验发现SR45可以与pre-mRNA、U1-70K和U2AF35相互作用,将U1 snRNP和U2AF分别招募到5′和3′剪接位点调节可变剪接。另外还发现SR45的RS1和RS2结构域可以独立与U2AF35蛋白结合。
甲基化、磷酸化、糖基化及乙酰化等蛋白修饰在可变剪接中发挥重要作用,但目前仅有关于SR-like甲基化和磷酸化修饰的研究报道。DNA胞嘧啶甲基化是一种表观遗传标记,常与基因和转座子的沉默有关。Ausin等[61]在研究预测的RNA相互作用域基因筛选DNA甲基化的突变时发现,SR45和Dicer-like3(DCL3)在RNA介导的DNA甲基化(RNA-directed DNA methylation,RdDM)途径中存在协同作用,表明SR45能够与DCL3相互作用,进而影响由RNA介导的DNA甲基化。AFC2是一种类似CDC2-2-like(LAMMOL/LER)的clk/Sty蛋白激酶,能够调控特定的SR蛋白的磷酸化状态,从而参与mRNA的可变剪切调控。Reddy[62]总结出SR45和AFC2、SCL33可以两两相互作用,在植物剪接位点的选择和剪接体组装的早期阶段发挥重要功能。
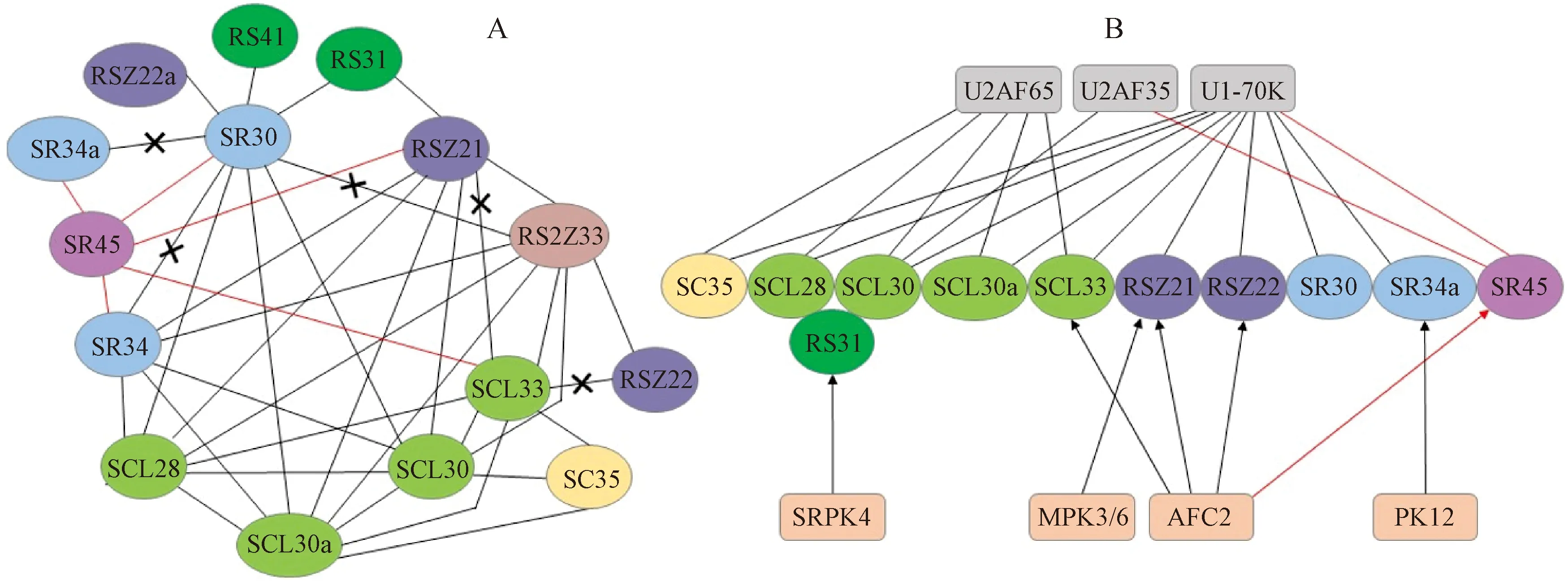
A.SR蛋白家族之间的相互作用;B.SR蛋白家族与剪接体蛋白或蛋白激酶之间的相互作用。椭圆形表示SR蛋白,同一亚家族的成员用相同的颜色标记。圆角矩形表示剪接体蛋白和激酶。红线和黑线表示蛋白之间的相互作用,带有“×”的黑线表示没有相互作用。黑色和红色箭头表示相应的激酶使SR蛋白磷酸化图2 拟南芥SR蛋白家族成员与剪接体蛋白/蛋白激酶间相互作用的分子网络[9,21- 22,30,58,60,63-68]A.Interactions between SR proteins;B.Interactions between SR proteins and spliceosomal proteins or protein kinases.SR proteins are shown in ovals,and the members of the same subfamily are marked with the same color.Spliceosomal proteins and kinases are shown in rounded rectangles.Red and black lines indicate protein interactions.Black lines with “×”indicate no interaction.Red and black arrows indicate phosphorylation of the SR proteins by corresponding kinasesFig.2 Protein interaction network of Arabidopsis SR proteins[9,21- 22,30,58,60,63-68]
2.3 参与调控植物生长发育和环境胁迫信号应答
植物的调控应答反应是一个涉及多基因、多信号途径及多基因产物参与的复杂过程,其中pre-mRNA可变剪接能够产生更多的转录本,并参与到植物生长发育、生物/非生物胁迫应答的调控过程中,这也是真核生物基因表达调控中的重要环节之一。SR-like蛋白不仅可以直接参与调控其他基因的剪接方式,还作为功能基因参与调控植株的生长发育。
在拟南芥种子萌发过程中,SR45负调控葡萄糖和脱落酸(abscisic acid,ABA)信号的转导。SR45通过影响ABA的积累和生物合成及信号转导相关基因(ABI3和ABI5)的表达,负调控葡萄糖响应应答;sr45-1突变体证明在幼苗发育早期对葡萄糖和ABA具有较高的敏感性[8]。SR45介导的葡萄糖负调控机制也存在于水稻种子萌发中,且ABA的代谢较ABA的生物合成更显著地抑制种子的萌发[69]。AtSR45在拟南芥根和花中的表达量很高[26],sr45-1突变株的生命周期缩短,但营养生长至生殖生长的过渡期延迟,晚花;叶片细长卷曲,根生长缓慢,花瓣与雄蕊数量不同,但控制花器官相关的基因表达却没有明显变化[36]。
在应答环境胁迫及植物激素信号时,植物SR-like蛋白显著地影响着一些基因的选择性剪接,包括形成一些新的剪接产物以响应胁迫[56,70-72]。如从表3可知,目前已从11种经生物/非生物胁迫的植物中鉴定到SR-like。
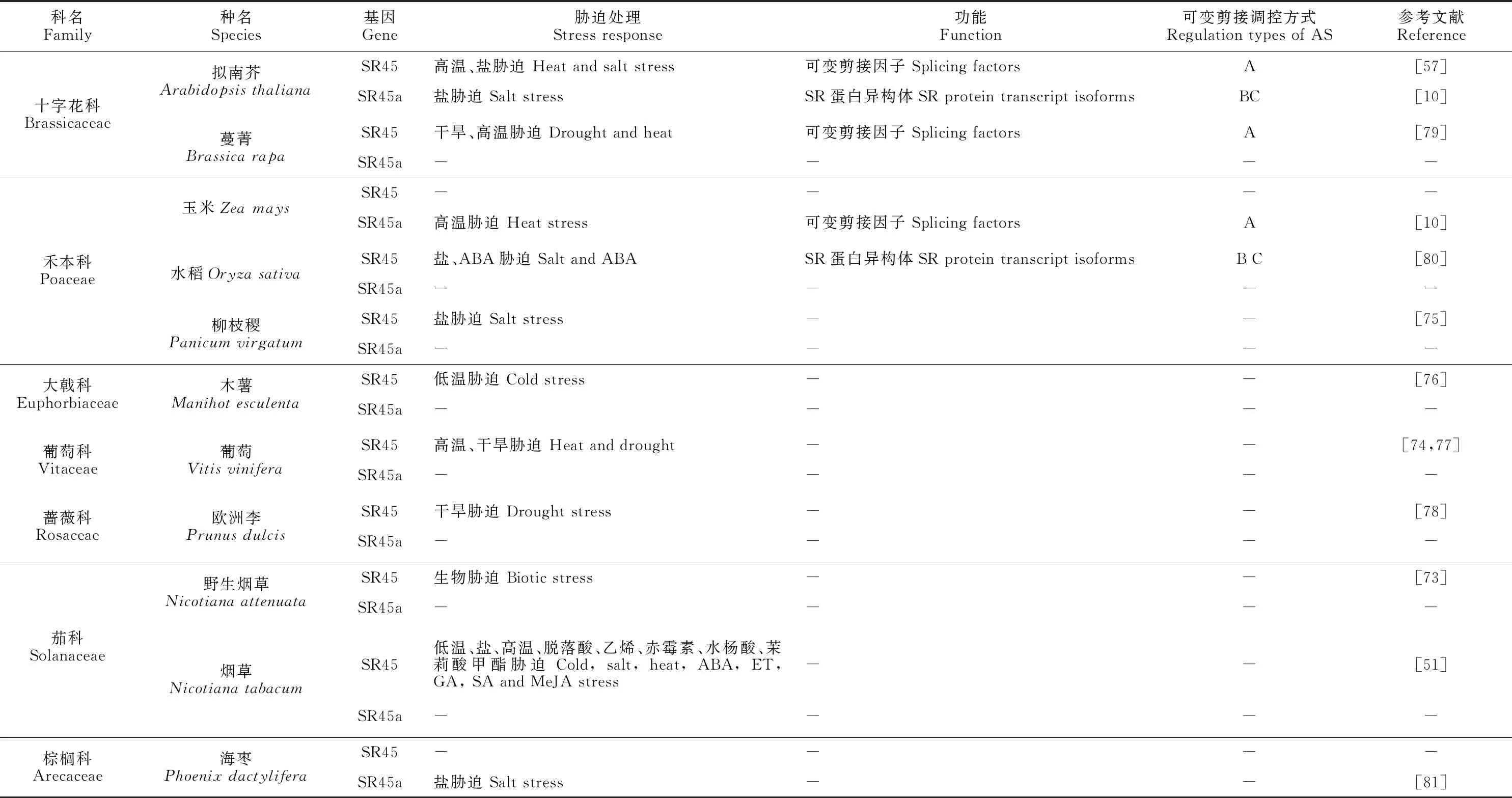
表3 生物/非生物胁迫下植物SR-like的功能和参与可变剪接的作用方式Table 3 Regulation types of AS and function about SR-like proteins in response to environmental stresses
关于SR45a生物胁迫的研究仅有1篇论文发表。Ling等[73]对烟草天蛾 (Manducasexta)摄食的野生烟草(Nicotianaattenuata)叶片和根进行了全基因组分析,发现烟草天蛾摄食野生烟草5 h后,叶片中总的可变剪接事件减少了7.3%,但根中增加了8.0%,这种生物胁迫显著改变了叶片和根中的可变剪接模式,并导致参与可变剪接的SR45a转录水平增加(在根中)和降低(在叶中);叶片中SR45a基因的表达不依赖茉莉酸(jasmonic acid,JA)途径,但在根中却依赖茉莉酸JA途径,表明生物胁迫引起的可变剪接和SR45a基因的差异表达在两种组织中都是独立调节的。为应对食草动物产卵和啃食等行为对植物造成的生物胁迫,激活防御系统至关重要,了解这一生物胁迫防御系统有助于今后培育食草耐受品种。
在SR-like蛋白参与非生物胁迫的相关研究中发现,非生物胁迫诱导剪接因子SR-like基因自身转录和表达模式发生改变,进而发挥功能。杨晓娜[51]发现NtSR45基因的表达受低温、干旱、高盐和高温等胁迫因素调控,其中低温、干旱和高盐胁迫下NtSR45基因下调表达;而高温诱导其表达量呈先下调、后上调的模式。高温胁迫下,葡萄SR45、SR30和SR34与核糖核蛋白组分U1 snRNP丰度显著上调[74]。在柳枝稷耐盐性相关的上游和下游的候选基因中发现,染色体6B上的QTL和SR45a具有85%的相似性[75]。李智博等[76]利用生物信息学重新构建了木薯SR45基因家族进化树,其转录组数据分析表明MeSR45亚家族均响应低温胁迫,且MeSR45-2显著上调表达。另外,SR-like蛋白还可以通过蛋白修饰改变剪接因子的亚细胞定位和移动性,从而改变其调控功能。Zhang等[77]对葡萄叶片进行磷酸蛋白质组学的比较分析发现,参与RNA剪接的SR45、RS2Z33、SCL33和SCL30的磷酸化水平显著上调,表明干旱会诱导剪接因子的磷酸化,从而增强耐热性。通过qRT-PCR 发现干旱胁迫伴随着‘Sefied’欧洲李叶片中AFC2 蛋白激酶的显著上调和随后RNA 剪接体蛋白(包括 SR45、SR33、SRZ22 和 RSZP21)的下调,验证了干旱胁迫对欧洲李AFC2蛋白激酶、SR45、SR33、SRZ22 和 RSZP21 控制的子网络激活的影响[78]。
依据目前已报道的5种植物SR-like在环境胁迫下行使可变剪接调控方式的不同,本文将SR-like应对非生物胁迫的调控机制总结归纳为以下3类(图3,表3):
(1)SR-like作为剪接因子通过自身可变剪接调节可变剪接的模式 在非生物胁迫下,SR-like可以发挥剪接因子功能进行可变剪接。Albaqami[57]利用atsr45突变体和转化全长或截短可变剪接异构体基因AtSR45的株系,研究了SR45及其剪接变异体在高温和盐胁迫中的作用。与野生型相比,SR45在不同发育阶段对高温和盐胁迫均表现出较高的敏感性,并且只有SR45-L长异构体在这些胁迫中起作用,因为长异构体可以缓解SR45株系对高温和盐胁迫的敏感性。通过进一步体外结合分析发现,SR45和热休克转录因子A2(heat shock transcription factorA2,HsfA2)的选择性剪接内含子结合,调控高温胁迫下HsfA2的可变剪接和表达;SR45在盐胁迫下会产生新的剪接异构体微调基因表达,表明SR45通过调节几个胁迫反应基因的表达和剪接,对这两种非生物胁迫的耐受性起到积极的调节作用。Muthusamy等[79]研究了正常条件和干旱条件下蔓菁野生型、过表达和突变体中与BrSR45a互作的关键耐旱性基因的可变剪接模式,表明BrSR45a通过浓度依赖性方式调节靶基因的选择性剪接应对干旱胁迫反应。干旱胁迫不仅使正常条件下的DCP5、RD29A、GOLS1、AKR、U2AF和SDR的剪接模式与过表达和突变体之间存在差异,还改变了 OCED2、SQE、UPF1、U4/U6-U5 tri-snRNP 相关蛋白和 UPF1 在过表达和突变体之间的剪接模式,表明过表达和突变体的功能丧失对靶基因的剪接模式有不同的影响。Li等[10]模拟了夏季日常温度变化,发现随着温度的升高,玉米SR45a发生可变剪接事件的频率升高,并使得SR45a的RNA异构体增加,表明SR45a的RNA异构体在应对高温时能够产生更有效的剪接调节。在每日的较低温度下,SR45a的mRNA会跳过保守结构域的外显子产生RNA异构体,翻译成SR45a蛋白质后与snRNPs组成剪接体后行使剪接功能;在每日的较高温度时,SR45a会保留保守结构域的外显子,编码功能有效的剪接调节因子,有可能增强其他基因的可变剪接(图3,A)。
(2)环境诱导逆境胁迫相关基因可变剪接产生异构体,再与下游靶标结合 环境诱导的逆境胁迫经常引入终止密码子,使得相关基因产生截短的异构体后再与下游靶标结合,在转录水平上对环境胁迫作出快速反应(图3,B)。这些截短的异构体通常保留了与全长蛋白质相同的互作靶点,但也有一些能力因为缺少关键结构域而丧失。例如,在盐胁迫下拟南芥SR45a发生可变剪接事件,产生两种剪接异构体——全长SR45a-1a和截短的SR45a-1b;全长 SR45a-1a 可用作剪接因子,而截短的 SR45a-1b 则不能。CBP20可与 SR45a-1a和 SR45a1b 发生物理相互作用;SR45a-1b 通过促进 SR45a-1a与CBP20 的结合介导盐胁迫信号转导通路[10]。
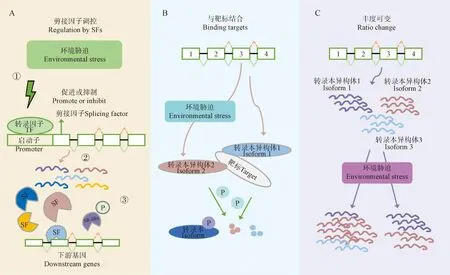
A.逆境胁迫通过剪接因子诱导可变剪接事件。SFs在多个水平上受到逆境胁迫调控:①环境胁迫促进或抑制剪接因子的转录;②许多剪接因子自身发生可变剪接事件,进一步调节下游基因的剪接;③逆境胁迫下SR-like蛋白发生磷酸化(标记为P);B.环境胁迫诱导产生不同的剪接异构体;C.可变剪接产生丰度可变的转录本异构体。环境胁迫导致剪接变异体的比例发生变化。图中外显子基因显示为框,内含子显示为线条图3 环境胁迫下剪接因子SR-like参与调控植物可变剪接的作用类型A.Stresses can induce alternative splicing through splicing factors (SF).SFs are regulated at multiple levels:① Environmental stress promotes or inhibits transcription of splicing factors;② Many splicing factors themselves generate alternative splicing events which further regulate splicing of downstream genes;③ SR-like proteins can be phosphorylated (labeled P) under stress;B.Environmental stress induces different splicing isoforms;C.Alternative splicing generates transcript isoforms with variable abundance.Environmental stresses lead to the altered ratios of splice variants.Exons are displayed as boxes and introns as lines in gene diagramsFig.3 Regulation types of alternative splicing about SR-like splicing factors in response to environmental stresses
(3)环境胁迫引起胁迫响应基因剪接异构体的比例发生变化 环境胁迫诱导逆境响应基因发生可变剪接,导致剪接异构体的比例发生变化,即通过可变剪接产生丰度可变的转录本异构体(图3,C)。例如,在非生物胁迫条件下,水稻免疫亲素FK506结合蛋白(Immunophilins FK506-binding proteins,OsFKBP20-1b)正向影响胁迫响应基因的转录和pre-mRNA 剪接。在osfkbp20-1b功能丧失的突变体中,应激反应基因表达下调,而其剪接异构体的表达量增加;过表达OsFKBP20-1b的植物中,相同的胁迫响应基因的表达显著上调。体内实验表明,OsFKBP20-1b 与处于核斑点和细胞质中的剪接因子 OsSR45 相互作用,维持剪接因子OsSR45蛋白的稳定性,并在非生物应激反应的转录后调控中通过介导RNA加工发挥重要作用[80]。
综上所述,环境信号能够调节植物自身SR-like基因pre-mRNA的剪接,产生不同亚型的SR-like蛋白及其他胁迫相关的蛋白,以适应生长发育和生物/非生物胁迫。
3 展 望
SR-like蛋白在转录和可变剪接两方面都具有重要的调控功能,并且与植物的生长发育、环境信号应答密切相关。由于SR-like独特的结构域使其成为SR蛋白研究领域的热点且取得了一系列突破性进展。但是关于SR-like仍然存在许多有待探索的内容,主要集中于以下几个方面。
一是关于研究的物种范围。目前多集中在模式植物(尤其是局限于单一的拟南芥)上,需进一步挖掘SR-like在非模式植物(如重要的经济作物、园林园艺植物)可变剪接及其转录调控上的功能。
二是关于SR-like精细的调控作用机制。随着高通量测序技术的普及,在植物的不同发育阶段、组织器官和生理生化过程中,越来越多的受SR-like调控的可变剪接事件被发掘出来,但大部分的研究仅从序列结构和表达水平对调控机制进行分析,与SR-like相关的可变剪接机制如何诱发、调节,是否可控和是否参与所有植物的生长发育过程仍是一片空白,其具体的调控机制也尚未形成共识。
三是关于SR-like响应生物与非生物胁迫的研究。目前仅有11种植物SR-like经过生物/非生物胁迫处理后的研究报道,对SR-like进行功能分析与可变剪接方式研究的只有5种植物,其中响应生物胁迫的研究仅有1篇文献,因此,需扩大物种且深入研究SR-like响应生物与非生物胁迫的调节机制。
SR-like参与的可变剪接是调控真核生物基因表达以及蛋白质多样性的重要机制,贯穿植物的整个且重要生命过程,目前还有很多未解之谜。利用关键SR-like基因的遗传转化技术才有可能实现SR-like蛋白时空表达水平,阐明植物可变剪接的分子机理,为提高植物抗逆能力和未来定向改良植物提供新线索。
