茶树CsbHLH71基因克隆及转录活性分析
2023-02-21吴应奇宋小凤邓见田烨白思蕾王坤波
吴应奇,宋小凤,邓见田烨,白思蕾,3,李 娟,3,王坤波,3*
(1 湖南农业大学 教育部茶学重点实验室,长沙 410128;2 湖南农业大学 国家植物功能成分利用工程技术研究中心,植物功能成分利用省部共建协同创新中心,长沙 410128;3 湖南农业大学 农业农村部园艺作物基因资源评价利用重点实验室,长沙 410128)
茶树[Camelliasinensis(L.)O.Kuntze]起源于中国西南地区,是世界广泛种植的一种叶用经济作物[1]。茶树中积累的次生代谢产物对茶叶的品质具有十分重要的作用[2]。儿茶素是茶树中一种主要的类黄酮次生代谢产物,是茶叶中造成苦、涩味的主要物质[3]。研究发现,向茶树叶面喷施微生物肥,有利于提高茶叶中氨基酸含量,降低酚氨比,提升茶叶的感官品质[4]和产量[5]。解淀粉芽孢杆菌(Bacillusamyloliquefaciens)是一种根际促生菌和有益内生菌,常被制成微生物菌肥用于农业生产[6]。然而,目前关于解淀粉芽孢杆菌微生物肥影响茶树中儿茶素生物合成的分子调控机制尚不明晰。
转录因子(transcription factors),也称为反式作用因子,是一组能够与靶基因启动子区域中顺式作用元件发生特异性相互作用从而调控靶基因表达的蛋白质[7]。碱性螺旋环螺旋(basic helix-loop-helix,bHLH)是植物体内仅次于MYB的第二大类真核转录因子家族[8],广泛参与植物的生长发育[9]、次生代谢生物合成[10]、非生物胁迫响应[11]和新陈代谢[12]等过程。研究发现,bHLH蛋白在茶树类黄酮次生代谢过程中扮演重要角色[13],能调控茶树类黄酮次生代谢产物如儿茶素的合成[14],从而影响茶叶的感官品质。茶树是一个遗传性十分复杂的植物,转录因子在茶树中的研究起步较晚,bHLH转录因子在茶树体内转录调控的分子机理还不清晰。因此明确茶树类黄酮代谢途径中的分子机理、挖掘茶树转录调控相关基因,对于阐明儿茶素生物合成的分子机制,进一步推动茶树转录水平的代谢调控机制和推动特异性茶树种质资源的开发利用具有重要意义[15]。
本研究以叶面喷施不同浓度微生物肥的茶树鲜叶为原料,结合茶树中儿茶素含量与转录组(RNA-seq)数据,筛选出了一个与儿茶素含量高度负相关的bHLH家族转录因子CsbHLH71。通过PCR克隆得到茶树CsbHLH71转录因子的全长CDS序列,并利用生物信息学分析该基因的结构、理化特征、进化关系及转录活性等,为进一步探究bHLH转录因子调控茶树中儿茶素生物合成的分子机制奠定基础。
1 材料和方法
1.1 试验材料
‘碧香早’茶树种植于湖南农业大学长安基地,微生物肥生态液(甘肃尚农生物科技有限公司,白银,甘肃,中国)有效成分为解淀粉芽孢杆菌EZ99,有效活菌数≥250亿/mL。将微生物肥分别稀释1 000(C1)和2 000(C2)倍,于夏季(6月)对统一修剪后的茶树进行叶面喷施,共喷施3次,每次间隔7 d,对照组(CK)进行喷水处理。待新梢萌发后,采摘茶树的一芽二叶叶片,迅速使用液氮冷冻处理,于-80 ℃储藏备用。
1.2 方 法
1.2.1 茶树CsbHLH71基因CDS序列克隆利用Primer Premier 5.0设计CDS克隆引物(表1),委托北京擎科生物科技有限公司进行引物合成。采用植物总RNA提取试剂盒提取‘碧香早’茶树鲜叶总RNA,逆转录得到cDNA,以cDNA为模板克隆目的基因,送北京擎科生物有限公司测序。

表1 本研究所用引物Table 1 Primers used in this study
1.2.2 茶树CsbHLH71基因生物信息学分析利用NCBI在线工具对茶树CsbHLH71蛋白氨基酸序列进行分析,在线软件SOPMA和SWISS-MODEL分别预测蛋白序列的二、三级结构。利用在线软件ExPASy中的ProtParam tool工具分析蛋白的理化性质,TMHMM server V.2.0在线工具分析CsbHLH71蛋白的跨膜结构。通过在线网站NCBI中Blast查找与茶树CsbHLH71蛋白同源序列,多序列比对采用DNAMAN软件,并利用MEGA 7.0软件构建系统进化树。Cell-PLoc 2.0在线预测CsbHLH71蛋白定位情况。
1.2.3 茶树CsbHLH71基因表达分析以‘碧香早’茶树鲜叶总RNA逆转录合成的第一链cDNA为模板进行实时荧光定量PCR(引物见表1),以β-actin为内参基因,3次生物学重复,采用2-ΔΔCT法[16]分析CsbHLH71基因在不同处理中的相对表达量。
1.2.4 茶树CsbHLH71蛋白的亚细胞定位为了验证CsbHLH71蛋白的亚细胞定位情况,将其通过同源重组法构建到pEAQ-GFP载体上并导入农杆菌中,注射到本氏烟草叶片中,适宜条件下培养48~72 h后,通过荧光显微镜观察GFP信号并拍照记录。具体参照程美女[17]实验方法。
1.2.5 茶树CsbHLH71蛋白的转录活性分析采用酵母分析系统及DLR烟草瞬时表达试验分析CsbHLH71蛋白的转录活性。1)酵母体内转录活性分析:将CsbHLH71基因构建到pGBKT载体上,将其转入到酵母感受态中,涂布至SD/-Trp和SD/-Trp/-Ade/-His缺陷筛选培养基上,28 ℃培养2~3 d。2)烟草体内的转录活性分析:将CsbHLH71基因构建到35S驱动的pBD载体上,作为Effector质粒,Reporter质粒为35S驱动的REN。将构建好的pBD-CsbHLH71质粒转入农杆菌中,将含有Reporter和Effector的农杆菌按1∶9的比例充分混匀,注射到本氏烟草,培养2~3 d,检测两种荧光素酶的比值(LUC/REN)。具体实验操作参照罗勇[18]的方法。
2 结果与分析
2.1 茶树CsbHLH71基因的CDS克隆与生物信息学分析
对喷施浓度梯度微生物肥的‘碧香早’茶树一芽二叶进行RNA-seq测序,对RNA-seq数据(未公开)进行差异基因筛选,发现CsbHLH71基因(TEA030725.1)与CK组相比,基因的表达量明显上升,且与叶片中儿茶素含量高度负相关。因此,我们设计了CsbHLH71的PCR特异性引物,从‘碧香早’茶树中克隆了该转录因子,扩增后得到约912 bp大小的片段。
ProtParam tool在线工具分析显示,CsbHLH71基因编码303个氨基酸,化学式为C1461H2371N419O455S15,蛋白分子量为33.6 kD,理论等电点为8.12,脂肪系数87.33,总平均亲水指数(GRAVY)为-0.280,不稳定系数59.00,是一种不稳定的亲水性蛋白。分析该基因编码蛋白的氨基酸组成,发现Leu(10.3%)和Ser(8.9%)含量较多,Trp(0.3%)和Cys(2.3%)等含量相对较少,带正电荷的残基总数(Arg+Lys)有34个,带负电荷的残基总数(Asp+Glu)有32个。
CsbHLH71蛋白跨膜结构分析表明,该蛋白的303个氨基酸都位于细胞膜,跨膜螺旋氨基酸残基数量的期望值是0.538 08,跨膜螺旋数量为0,CsbHLH71蛋白不是跨膜蛋白。通过NCBI对氨基酸序列进行分析,发现CsbHLH71蛋白包含一个从101-160位的碱性螺旋-环-螺旋(bHLH)保守结构域,属于bHLH家族蛋白。
CsbHLH71蛋白的二级结构预测表明,此蛋白303个氨基酸包含47.52%无规则卷曲,40.59%α-螺旋,9.24%的延长链,2.64%β转角(图1,A)。使用SWISS-MODEL预测CsbHLH71蛋白的三维结构与二级结构预测的结果一致,该蛋白的三维空间结构主要是螺旋和卷曲(图1,B)。

A(蓝色.α螺旋;红色.延伸带;绿色.β转角;紫色.无规则卷曲)图1 CsbHLH71蛋白的二级结构(A)和三级结构(B)A(Alpha helix,extended strand,beta turn and random coil are represented by blue,red,green and purple,respectively)Fig.1 The prediction of secondary structure (A) and tertiary structure (B)
将CsbHLH71蛋白氨基酸序列上传到NCBI数据库进行Blast比对,发现CsbHLH71蛋白具有典型的bHLH结构域,与其他植物bHLH氨基酸序列相似,这表明不同植物bHLH氨基酸序列保守性较高,只有少部分的结构域发生了细微变异(图2)。系统发育树结果(图3)显示,CsbHLH71蛋白与中国猕猴桃变种、杜鹃花的亲缘关系最近,而与栎木、核桃的进化距离较远。

CsbHLH71.茶树;AcbHLH71-like.中华猕猴桃变种;DzbHLH71-like.榴莲;JrbHLH71-like.核桃;PtbHLH71.毛果杨;PvbHLH71-like.黄连木;QlbHLH71-like.栎木;RgRHGRI-protein.杜鹃花;VrbHLH71-like.河滨葡萄;VvbHLH71.葡萄图2 CsbHLH71蛋白与其他物种氨基酸序列同源比对CsbHLH71.Camellia sinensis;AcbHLH71-like.Actinidia chinensis var.chinensis;DzbHLH71-like.Durio zibethinus;JrbHLH71-like.Juglans regia;PtbHLH71.Populus trichocarpa;PvbHLH71-like.Pistacia vera;QlbHLH71-like.Quercus lobata;RgRHGRI-protein.Rhododendron griersonianum;VrbHLH71-like.Vitis riparia;VvbHLH71.Vitis viniferaFig.2 Multiple alignment of CsbHLH71 protein with other bHLH71 proteins of other species

图3 CsbHLH71蛋白与不同植物进化关系Fig.3 Phylogenetic relationships between CsbHLH71 protein and other species bHLH71 proteins
2.2 茶树CsbHLH71基因表达分析
为验证转录组中CsbHLH71基因的表达结果,我们利用qRT-PCR进行了验证。结果(图4)表明,在不同浓度微生物肥处理后,CsbHLH71基因表达水平显著上调,但在1 000倍中上调趋势更为显著,说明微生物稀释1 000倍处理对CsbHLH71基因表达更加明显,这与转录组数据一致。

CK.对照,喷施蒸馏水;C1和C2分别指叶面喷施稀释1 000倍和2 000倍的微生物肥;*与**分别表示与对照组(CK)相比差异显著(P<0.05)或极显著(P<0.01)图4 不同菌肥处理茶树鲜叶中CsbHLH71基因的表达CK.Spraying with distilled water;C1 and C2.1 000 fold and 2 000 fold dilution;* and ** mean significant (P<0.05)and extremely significant (P<0.01) differences compared with CK,respectivelyFig.4 Expression of CsbHLH71 gene in leaves of Camellia sinensis under different microbial fertilizer treatments
2.3 茶树CsbHLH71蛋白的亚细胞定位
Cell-PLoc 2.0在线预测表明CsbHLH71蛋白可能定位于细胞核。对浸染农杆菌的烟草叶片拍照记录,发现阳性对照GFP整个细胞都能观察到绿色荧光,而侵染了CsbHLH71-GFP的烟草只在细胞核中观察到绿色荧光(图5),说明CsbHLH71蛋白定位于细胞核,属于核蛋白。
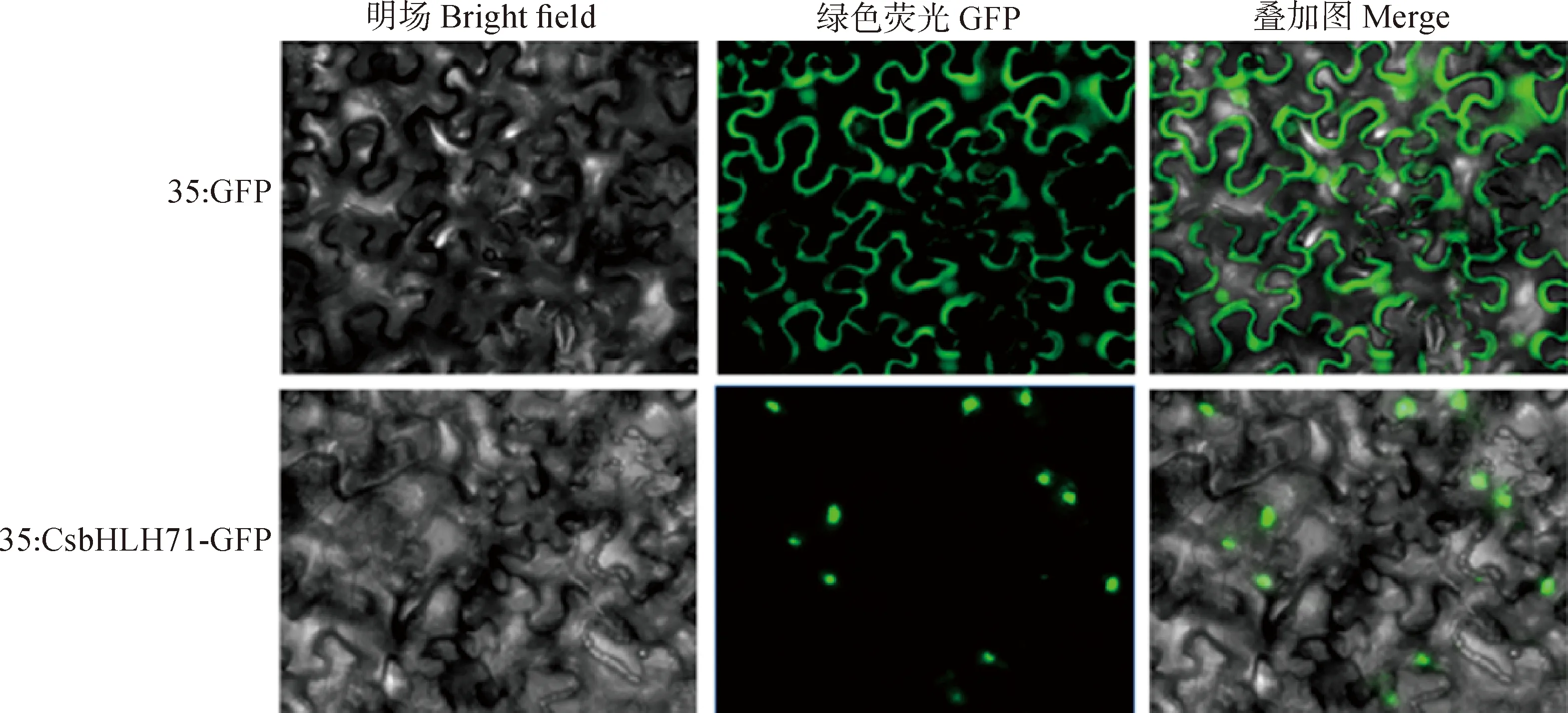
图5 CsbHLH71蛋白亚细胞定位Fig.5 Subcellular localization of CsbHLH71 protein
2.4 茶树CsbHLH71蛋白的转录活性分析
在酵母体内(图6,A),转化菌株在SD/-Trp缺素培养基上均正常生长,在SD/-Trp/-Ade/-His上只有阳性对照P-53的酵母菌株可以正常生长,且X-α-Gal活性检测变蓝,而阴性对照DBD-Empty和DBD-CsbHLH71的酵母菌株均不能正常生长,说明CsbHLH71蛋白在酵母内为抑制活性;在烟草体内(图6,B、C),pBD-CsbHLH71 LUC/REN数值比阴性对照pBD-Empty低。说明茶树CsbHLH71蛋白在烟草体内表现出转录抑制活性,属于转录抑制子。这些结果表明CsbHLH71蛋白在茶树中可能通过抑制下游基因的表达行使功能。
3 讨 论
茶树次生代谢过程中转录因子起着重要调控作用[19]。例如,Gong等[20]发现日照强度通过影响bHLH、MYB、NAC等转录因子影响黄酮类化合物合成途径中重要基因,如CHS、F3′5′H、SCPL和DFR的表达,导致茶叶中儿茶素在三个季节的含量不同。Liu等[21]发现遮阴降低了茶树中儿茶素和黄酮醇的含量,与其生物合成相关的功能基因F3′H、FLS、ANS、ANR、LAR、DFR和CHS及转录因子MYB4、MYB12、MYB14和MYB111表达水平同时下调。
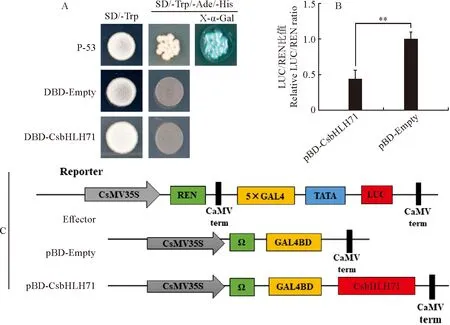
**表示与阴性对照比差异显著(P<0.05)图6 CsbHLH71蛋白在酵母(A)和烟草(B、C)的转录活性分析** means significant difference compared with contrast (P<0.05)Fig.6 Transcriptional activity analysis of CsbHLH71 protein in yeast cell assays (A) and tobacco (B,C)
本研究通过RNA-seq测序并从中筛选获得一个茶树CsbHLH71基因,生物信息学分析表明,CsbHLH71蛋白长度为303个氨基酸,含有一个59位的碱性螺旋-环-螺旋(bHLH)保守结构域,属于bHLH蛋白家族。氨基酸序列比对发现,在不同植物中bHLH家族氨基酸序列相似性较高,只有少数有细微变异,说明CsbHLH71蛋白在进化中相对保守,但不同植物的bHLH蛋白在进化过程中呈现出了多样性。qRT-PCR分析表明,茶树在微生物肥稀释喷施处理下,CsbHLH71基因表达水平明显高于未施肥的CK组,但在稀释2 000倍处理中CsbHLH71基因的表达水平虽然较CK组有所上升,但不如1 000倍处理下效果显著,说明高浓度的微生物肥可能促进茶树CsbHLH71基因的表达。
亚细胞定位显示CsbHLH71蛋白和大多数转录因子蛋白一样定位于细胞核,是一个核蛋白,在细胞核内行使转录调控功能。研究发现,转录因子并非都是转录激活因子,也会阻遏目的基因的表达,起负调控作用。Xu等[22]在银杏中发现了GbMYBF2负调控类黄酮合成,并通过遗传转化证实了它的功能。Luo等[23-24]在茶树中发现CsbHLH62、CsWRKY31和CsWRKY48能够抑制CsLAR、CsDFR和CCoAOMT,从而负调控EGCG3″Me的积累。在本研究中,我们发现CsbHLH71蛋白在酵母细胞中具有转录抑制活性,并通过在烟草的转录活性分析验证了这一结果,证实了CsbHLH71蛋白具有转录抑制剂的功能,可能负调控茶树类黄酮代谢合成产物儿茶素积累。CsbHLH71基因的表达水平与儿茶素含量呈负相关,猜测CsbHLH71基因可能参与了茶树类黄酮代谢调控儿茶素的生物合成。通过分析本课题组前期运用高效液相色谱法检测不同浓度微生物肥喷施处理下茶树类黄酮代谢产物儿茶素含量发现,部分儿茶素合成相关功能基因的表达水平受到了明显抑制。因此推测,CsbHLH71基因通过参与茶树类黄酮次生代谢,影响儿茶素合成相关基因的合成通路,但其与茶树儿茶素合成功能基因之间的调控关系还需要进一步研究。
研究发现,bHLH转录因子还能与MYB、WD40合作形成三元MBW复合体来参与调控次生代谢产物的合成。Zhao等[25]发现MBW三元复合体可控制黄酮类化合物合成途径中的多个酶促步骤。Chen等[26]发现R2R3-MYB蛋白可以直接结合靶基因的启动子或作用于MBW复合物以抑制花青素的生物合成。Liu等[27]发现茶树中MBW复合体能调节花青素和原花青素的积累。通过分析儿茶素合成的一些关键功能基因启动子发现,序列中包含大量可以与bHLH、MYB、WRKY等转录因子结合的元素,如G-Box、E-Box、W-Box等,推测CsbHLH71蛋白可能存在与其他转录因子蛋白之间相互作用,形成复合体共同调控儿茶素的合成。未来将继续通过凝胶阻滞迁移实验(EMSA)、染色质免疫共沉淀(CHIP-qPCR)等方法,对CsbHLH71基因进行深入研究,以揭示CsbHLH71基因调控儿茶素合成的分子机制。
