多营养层次养殖的海水池塘浮游植物群落特征
2023-02-14方民杰
方民杰
(福建省水产研究所,福建厦门 361013)
0 引言
池塘养殖是中国水产养殖的主要方式,历史悠久,规模庞大,养殖模式和品种多样。近年来,标准化池塘养殖、多营养层次池塘综合养殖、工程化池塘循环水养殖等多种模式持续创新,极大推动池塘养殖业的快速发展,为渔业高质量绿色发展注入新的动能[1]。多营养层次养殖是由不同营养级生物组成的综合养殖系统,可充分利用输入其中的营养物质和能量,提高池塘养殖容纳量,具有综合效益高、养殖环境稳定、生态健康的特点[2-3],该养殖模式的开发与应用已达到产业化水平,并于2014年被农业农村部遴选列入农业主推技术目录,已在中国沿海普遍应用[4]。在福建省漳浦县佛昙湾垦区,绝大多数海水池塘采用“鱼-虾-贝”多营养层次养殖模式。秋季至次年春夏季的生产季节中,一般以菊黄东方鲀(Takifugu flavidus)或双斑东方鲀(T.bimaculatus)、日本囊对虾(Marsupenaeus japonicus)、菲律宾蛤仔(Ruditapes philippinarum)为主要养殖对象。在这种养殖模式中,鱼虾需要人工投饵,鱼类摄食病虾阻断疾病传播,浮游植物利用水体中营养盐进行生长繁殖,贝类滤食水体中的有机碎屑和浮游植物[3]。
池塘水体中浮游植物群落对保障养殖贝类的饵料供应、维持水质稳定和养殖成功[5]具有重要意义,但某些种群数量急剧增加时,也存在着爆发水华的潜在危险[6]。因此,有必要对养殖水体的浮游植物群落结构进行研究分析,为养殖生产提供科学指导。骆云慧等[7]在凡纳滨对虾-缢蛏生态循环养殖研究中发现,硅藻种群变化主要与温度、盐度、pH相关,绿藻种群优势受氮含量的较大影响,而甲藻、蓝藻与磷含量相关。高欣等[8]研究表明凡纳滨对虾高位养殖池塘浮游植物优势种的主要驱动因子为盐度和氮磷。王旭娜等[9]研究认为凡纳滨对虾养殖池塘浮游植物结构主要受到温度、pH以及氮磷含量的影响。孙耀等[10]认为在对虾养殖水环境中浮游植物量与无机氮之间呈较好的幂函数衰减曲线关系,而与无机磷相关性较差。许永健等[11]研究发现海水池塘中海洋原甲藻水华爆发与硝酸氮浓度升高有关。在对池塘浮游植物与理化因子相关性进行分析的基础上,部分研究也提出池塘生态调控、养殖模式优化等措施建议。彭聪聪等[12]认为由优良微藻优势种类形成的藻相能对凡纳滨对虾半集约化养殖环境的稳定起到积极作用,郭永坚等[13]提出在凡纳滨对虾-鲻网围分隔混养池塘中应调整鲻的放养密度。
目前关于池塘水体浮游植物群落特征及其与氮磷等理化因子的相关性已有较多研究,但对菊黄东方鲀、日本囊对虾、菲律宾蛤仔为主要养殖对象的多营养层次养殖模式的浮游植物群落相关研究尚未见报道。为了掌握“鱼-虾-贝”海水池塘浮游植物群落变动情况,并为养殖生态调控、多营养层次养殖模式的优化提供数据支撑和科学依据,于养殖初、中、后期采集池塘水样和浮游植物样品,进行检测和分析。
1 材料和方法
1.1 试验池塘基本情况
试验池塘位于福建省漳浦县佛昙湾垦区,共有3口池塘 ,编号为 1、2、3 号 ,面积分别为 3.0、1.3、2.0 hm2。池塘开挖环沟,中间形成平整的埕地,其中用于底播养殖菲律宾蛤仔的面积约占池塘面积的15%。每口池塘各有1个进排水闸门,利用潮水涨落进行换水,池内均配备水车式增氧机。
1.2 养殖过程
试验池塘经过清淤、晒池、消毒、进水并肥水等前期准备之后,于2020年10月22—23日投放菊黄东方鲀大规格苗种和日本囊对虾苗,12月31日投放菲律宾蛤仔大规格苗种。鱼虾放苗后的30天内只进水,以后根据天气、水色、水温、投饵等情况适当换水。冬季低温期,一般每3~5天换水10%,春季水温回升后,每2~3天换水20%。养殖全程机械增氧。菲律宾蛤仔放苗后,根据水色、天气等情况,适量施放发酵鱼浆,以调节藻相。
饲料投喂量、投喂次数根据养殖鱼虾大小、密度、摄食、活动、水温、水质、天气等情况而定。日本囊对虾、菊黄东方鲀、菲律宾蛤仔分别于2021年1—4月、4—5月、5—7月捕捞上市。
1.3 样品的采集与分析
2020年10月28日、2021年1月25日、2021年4月23日,3口池塘各采样1次,以闸门外进水为对照点(编号为4号),代表自然海域。于上午10时进行,浮游植物样品采集1000 mL表层水样,并用鲁哥氏液固定;营养盐样品采集500 mL表层水样。按照GB 17378—2007海洋监测规范进行样品的分析,分析项目为无机氮、活性磷酸盐和浮游植物的种类、数量。
1.4 数据处理
用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)表征浮游植物群落特征。见公式(1)~(2)。


采用Excel软件进行数据处理,并绘制图表;浮游植物主要种类的细胞密度、营养盐(氮磷)含量数据均进行lg(x+1)转换[14],然后使用SPSS软件分析相关性。
2 结果与分析
2.1 种类组成
由图1可知,养殖期间3口池塘共鉴定浮游植物3门34属60种(包括变种、变型),其中硅藻门25属48种,占80.00%;甲藻门8属11种,占18.33%;金藻门1属1种,占1.67%。对照点共鉴定浮游植物3门25属35种,其中硅藻门20属29种,占82.86%;甲藻门4属5种,占14.29%;金藻门1属1种,占2.86%。

图1 浮游植物种类数变化
3次采样各池塘浮游植物种类数在17~25种之间,对照点在13~17种之间,池塘浮游植物种类数均大于对照点。
2.2 细胞密度
由图2可知,养殖期间10月、1月、4月池塘浮游植物的细胞密度的均值分别为 21.67×104、22.23×104、10.84×104cells/L,以1月最高,4月最低。对照点10月、1月、4月浮游植物的细胞密度分别为20.30×104、10.77×104、7.57×104cells/L,以10月最高,4月最低。养殖初期部分池塘的浮游植物细胞密度稍低于对照点,但在养殖中后期均高于对照点。
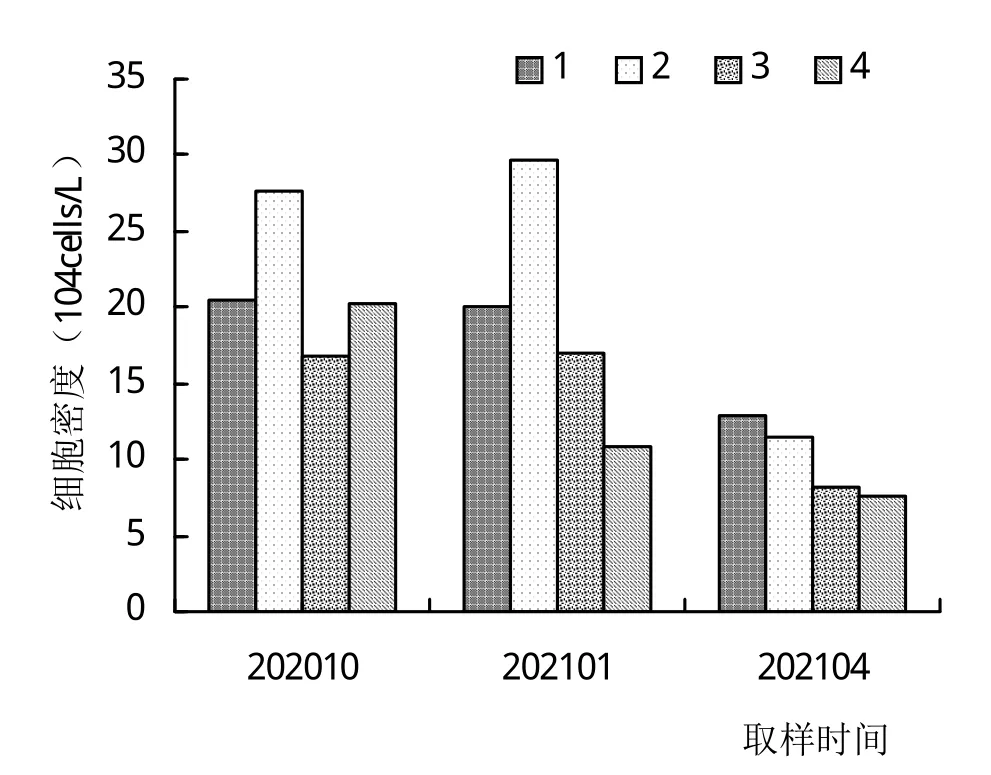
图2 浮游植物细胞密度变化
2.3 优势种和主要种
10月养殖池塘的浮游植物优势种为中肋骨条藻(Skeletonema costatum),细胞密度占总细胞密度的41.69%,其次为笔尖形根管藻(Rhizosolenia styliformis)、劳氏角毛藻(Chaetoceros lorenzianus),分别占6.15%和6.10%。对照点优势种为中肋骨条藻,细胞密度占总细胞密度的47.13%,其次为丹麦细柱藻(Leptocylindrus danicus)、窄隙角毛藻(Chaetoceros affinis),分别占14.29%和6.73%。中肋骨条藻在池塘和对照点均占优势。
1月养殖池塘的浮游植物优势种为锥状斯克里普藻(Scippsiella trochoidea),细胞密度占总细胞密度的77.31%,其次为中肋骨条藻,占4.37%。对照点优势种为中肋骨条藻,细胞密度占总细胞密度的47.52%,其次为锥状斯克里普藻、奇异棍形藻(Bacillaria paradoxa),分别占11.15%和7.43%。锥状斯克里普藻在池塘占绝对优势,而中肋骨条藻在对照点占优势。
4月养殖池塘的浮游植物主要种为中肋骨条藻、米氏凯伦藻(Karenia mikimotoi)、锥状斯克里普藻,细胞密度分别占总细胞密度的22.39%、14.46%和7.95%。对照点主要种为中肋骨条藻、具槽直链藻(Melosira sulcata)、锥状斯克里普藻,细胞密度分别占总细胞密度的22.99%、16.78%和15.33%。米氏凯伦藻在其中2口池塘成为主要种,而中肋骨条藻在池塘和对照点均为主要种(见表1)。
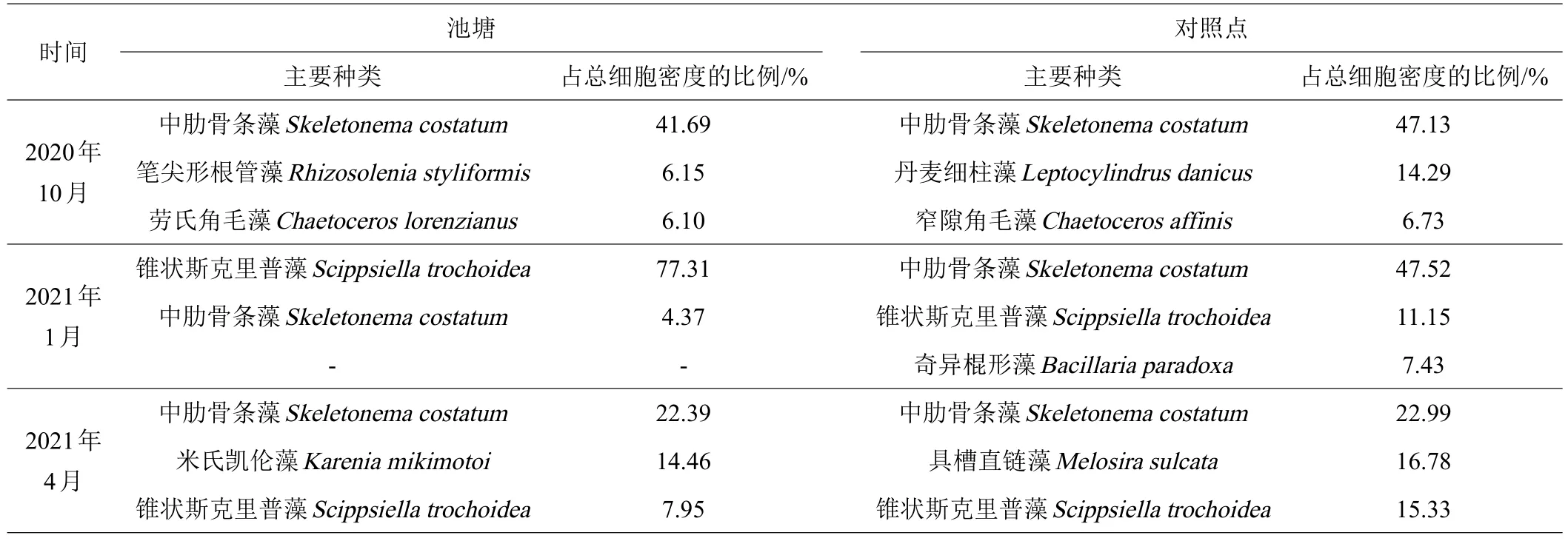
表1 养殖池塘与对照点的浮游植物优势种及主要种
2.4 多样性特征
群落多样性是反映群落稳定性的重要参数,养殖池塘受到投饵、施肥及天气等因素的影响,容易形成浮游植物大量繁殖和消亡现象,导致群落多样性的激烈波动。1月池塘浮游植物种类多样性指数(H′)、均匀度指数(J)明显低于10月和4月,这与1月锥状斯克里普藻占总细胞密度的比例高达77.31%有关。对照点的多样性指数、均匀度指数波动较小(见表2)。
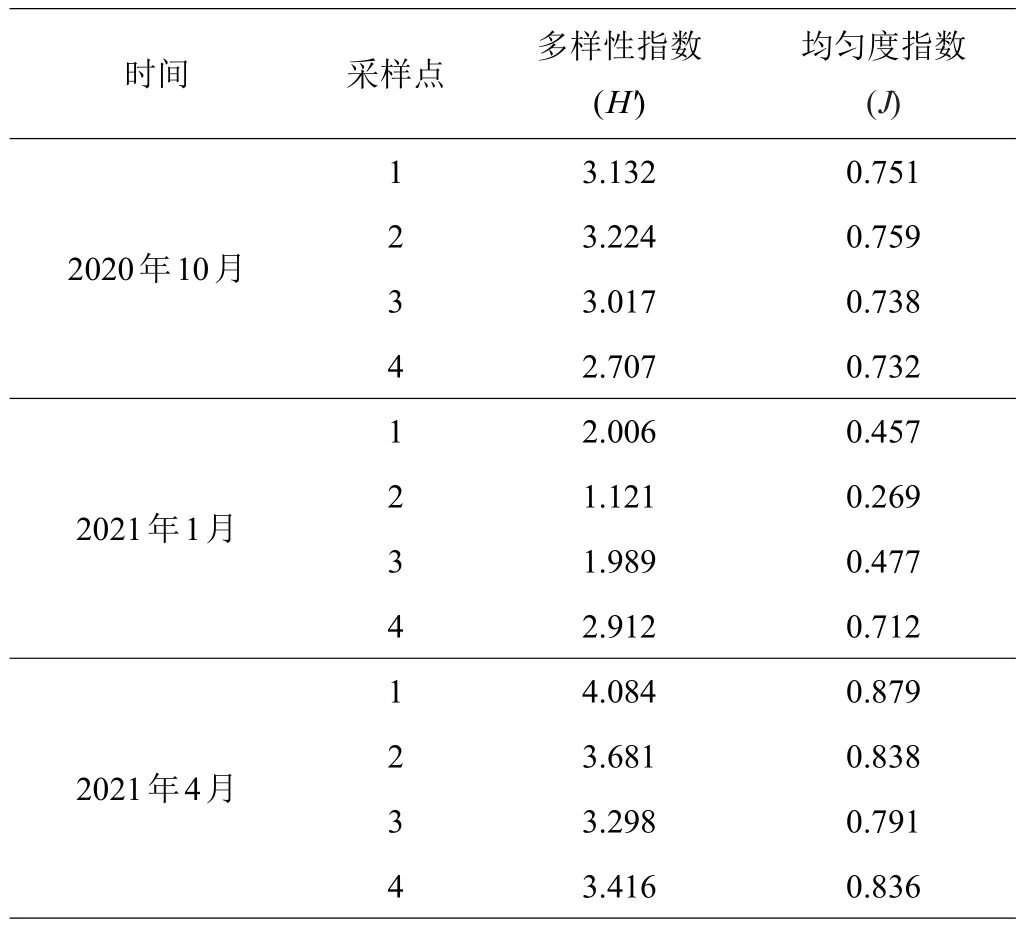
表2 浮游植物群落多样性指数
2.5 营养盐
养殖期间,池塘水体无机氮含量均处于低值,并低于对照点;10月、1月活性磷酸盐含量大多处于低值,并均低于对照点,但4月各池塘水体活性磷酸盐含量出现大幅度升高现象,并远高于对照点。对照点氮磷含量均大幅超过海水水质二类标准[15],且波动较小(见表3)。
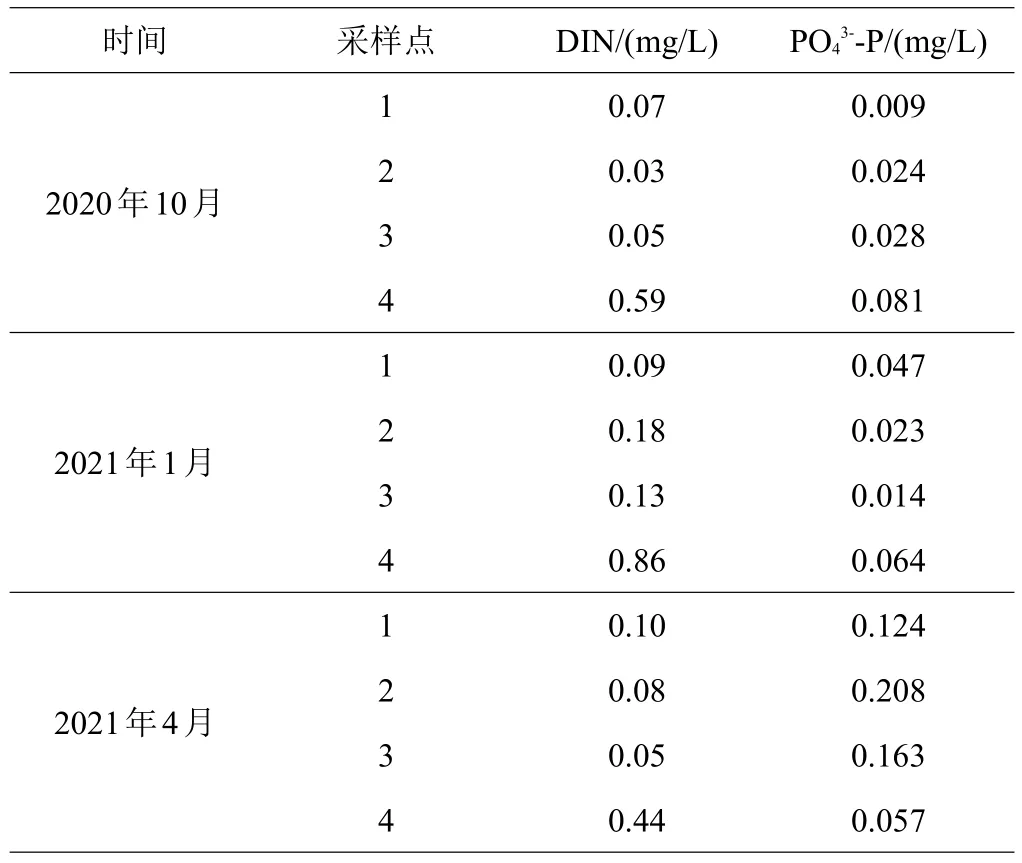
表3 氮磷含量
2.6 浮游植物主要种类与氮磷含量的相关性分析
选取在养殖池塘中占浮游植物总细胞密度比例较高的3个主要种类:中肋骨条藻、锥状斯克里普藻、米氏凯伦藻,分析其细胞密度与氮磷含量之间的相关性。分析结果显示,米氏凯伦藻细胞密度与活性磷酸盐含量极显著相关(P<0.01),但与无机氮含量相关性不显著(P>0.05)。中肋骨条藻、锥状斯克里普藻细胞密度与氮磷含量相关性均不显著(P>0.05)(见表4)。
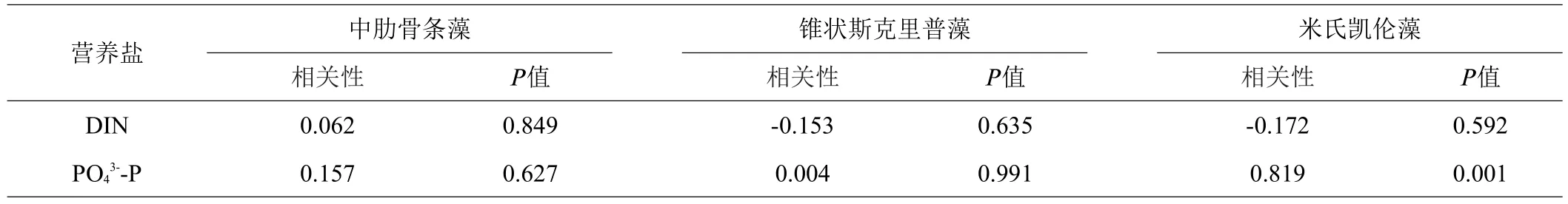
表4 浮游植物主要种类细胞密度与氮磷含量相关性
3 讨论
3.1 浮游植物群落特征
养殖池塘和对照点的浮游植物从种的数量上来看,种类数以硅藻占绝大多数,甲藻次之,金藻最少,池塘浮游植物种类数均大于对照点。从浮游植物细胞密度上来看,养殖初期池塘水体浮游植物以中肋骨条藻为优势种,中期以锥状斯克里普藻为绝对优势种,后期以中肋骨条藻、米氏凯伦藻、锥状斯克里普藻为主要种,呈现由硅藻占优势向甲藻占优势的演替趋势。而对照点均以硅藻占绝对优势,但在中后期由于锥状斯克里普藻占总细胞密度的比例较大,导致甲藻比例有所上升。
郭丰等[16]研究表明,虾池浮游植物种类数多于海区,硅藻、甲藻、绿藻是构成厦门潘涂垦区虾池浮游植物的三类主要成分,虾池甲藻类的种类数和细胞数量都明显高于自然海区;硅藻类比甲藻类有较高的营养盐需求,虾池的营养盐含量较低、有机质含量丰富和水体扰动性较差,可能是决定硅藻类和甲藻类种类和数量比例的重要因素。查广才等[17]研究表明,低盐度对虾养殖池浮游植物种类以绿藻和蓝藻为主,数量以蓝藻占绝对优势。本研究结果表明池塘浮游植物以硅藻、甲藻为主,未见绿藻和蓝藻,这与采样池塘离河流入海口较远、盐度较高有关;池塘浮游植物种类数、甲藻类的种类数和细胞数量都高于对照点,这与潘涂垦区一致,也说明了池塘与自然海域之间环境条件存在差异。
3次采样结果显示,各养殖池塘的浮游植物种类多样性指数、均匀度指数均以1月为最低,这与1月锥状斯克里普藻占绝对优势有关。多样性指数和均匀度是反映群落结构复杂性和稳定性的重要指标,其值越大,群落结构越复杂和稳定,而浮游植物的多样性指数和均匀度指数低,不利于养殖水环境的稳定,可能导致病害的发生[5]。对照点多样性指数、均匀度指数波动较小,说明池外自然海域水体环境较为稳定。李由明等[18]研究发现,对虾高位池养殖期间,水体中浮游植物的多样性指数较低,平均在0.83~1.48,浮游植物群落结构比较简单。本研究中池塘水体浮游植物的多样性指数变动范围为1.121~4.084,显示对虾高位池水体环境稳定性不如多营养层次养殖池塘。
3.2 浮游植物主要种类和营养盐的关系
氮磷营养盐是影响浮游植物的生长、数量变动和群落演替的重要因素,而浮游植物是养殖池塘中菲律宾蛤仔的主要饵料来源,因此,池塘水体的氮磷含量也是影响养殖生产的重要因素。在整个养殖过程中,3口池塘水体无机氮含量均处于低值,而活性磷酸盐含量在初期、中期基本处于低值,但在养殖后期剧增,这与菲律宾蛤仔在生长过程中代谢排放[19]及浮游植物对氮磷吸收[20]的不平衡有关。
中肋骨条藻是典型的广温广盐性种类,是福建沿岸海域近年来分布最广泛的、细胞密度最大的赤潮生物,几乎一年四季都是该海域的优势种[21],在秋季以后易成为虾池优势种[16];中肋骨条藻在氮磷比高的水体中生长状态较氮磷比低的水体更好,表明在富营养化条件下氮比磷更易诱发中肋骨条藻赤潮[22]。本研究中池塘水体无机氮均处于低值,而中肋骨条藻的细胞密度最高值出现于10月,为1.12×105cells/L,1月和4月的最高值仅为104cells/L量级,说明较低的无机氮含量和氮磷比限制了中肋骨条藻的大量繁殖。
锥状斯克里普藻为世界广布性近岸赤潮生物,也是中国沿海常见优势甲藻类。张玉娟等[23]研究结果表明,N、P浓度分别超过10µg/L、1.5µg/L就能维持锥状斯克里普藻一定的种群数量,而营养元素充足时,锥状斯克里普藻可以长期保持较高细胞密度。本研究中,养殖初期、中期池塘水体氮含量处于低值,磷含量大多处于低值,1月锥状斯克里普藻成为绝对优势种,其最高密度2.56×105cells/L,已接近赤潮的基准密度(1×106cells/L)[24],说明该藻对低氮磷含量具有适应性。
米氏凯伦藻是中国海域常见的赤潮甲藻,为世界广布种。龙华等[25]研究表明,米氏凯伦藻适温范围是20.5~24℃,最高的细胞密度出现在23℃左右,高营养指数(E)、丰富的磷酸盐含量和低氮磷比是米氏凯伦藻赤潮形成的重要条件。姚炜民等[26]认为米氏凯伦藻生长所需最佳的水温为23.4~23.8℃,磷酸盐浓度的变化与以米氏凯伦藻为主的赤潮生物的密度呈正相关关系。刘志国等[27]研究了中国沿海米氏凯伦藻分布和引发赤潮历史,2002—2012年,福建省沿海米氏凯伦藻赤潮都发生在5月初至6月中旬。在本研究中,10月、1月均未发现米氏凯伦藻,而4月下旬在池塘和对照点均发现较大数量的米氏凯伦藻,并成为养殖池塘浮游植物的主要种之一,其最高密度2.93×104cells/L,低于赤潮的基准密度(1×106cells/L)[24];相关性分析结果显示,其细胞密度与活性磷酸盐含量之间具有显著相关性。总之,米氏凯伦藻繁盛的季节及其活性磷酸盐含量相关性与前人研究结果相符合。
3.3 米氏凯伦藻的危害及治理措施
米氏凯伦藻属甲藻门、裸甲藻目,是一种有毒有害赤潮藻类,能分泌溶血性毒素,引起鱼类、贝类等生物死亡,造成严重渔业损失[26-27]。目前,对米氏凯伦藻赤潮发生及危害的报道均为开放式海域,而福建省沿海区域是米氏凯伦藻赤潮的主要发生区域[27]。本研究中,池塘水体出现较大数量的米氏凯伦藻,可能是自然海域中的米氏凯伦藻随着池塘换水而进入,在适宜水温、营养盐等环境条件下大量繁殖。尽管其密度尚未达到赤潮的基准密度,但也应引起重视,提前采取治理措施,避免养殖生物出现死亡。
目前,对于池塘赤潮或水华的治理,一般都是采用化学、物理、生物方法[28-30]。在养殖实践中,可利用海风、光照等因素引起的米氏凯伦藻局部聚集,在聚集区泼洒杀藻药物如氧化钙、二氧化氯、漂白粉、过氧化氢、高锰酸钾等,除可直接杀灭米氏凯伦藻外,还可通过强氧化作用,降低毒素危害[30];投放杀藻细菌如Thalassospira sp.ZR-2,该细菌可通过分泌苯甲酸,促进米氏凯伦藻细胞的分解[31-32];泼洒絮凝剂如粘土悬浊液[29-30];也可采取机械移除措施,如打捞、水泵抽出等[30]。此外,还应及时换水,减轻米氏凯伦藻的不利影响。
当然,对于池塘米氏凯伦藻的危害,应首先采取必要的预防措施。春季水温回升,“鱼-虾-贝”多营养层次养殖池塘水体中活性磷酸盐逐渐积累,形成有利于米氏凯伦藻爆发的条件,养殖户应加强巡池观察,注意海洋与渔业部门发布的赤潮监测预警信息,避免在赤潮发生时进水。在养殖中后期,可根据池塘水体扰动性较差及氮磷含量变动等特点,采取适当增大换水量[30]、酌情补充氮肥[10]、施加除磷微生物制剂[33]等必要的生态调控措施,并调整鱼虾贝配比,适当降低贝类养殖密度,优化多营养层次养殖模式,构建有利于养殖生物生长的水生态环境。
4 结论
“鱼-虾-贝”多营养层次养殖的3口海水池塘共鉴定浮游植物60种,包括变种、变型,种类数高于进水对照点的35种。养殖初期部分池塘的浮游植物细胞密度低于对照点,但在养殖中后期均高于对照点。养殖期间池塘水体的浮游植物群落结构呈现由硅藻占优势向甲藻占优势的演替趋势,而对照点均以硅藻占绝对优势。1月池塘浮游植物种类多样性指数、均匀度指数明显低于10月和4月,这与1月锥状斯克里普藻占绝对优势有关。对照点种类多样性指数、均匀度指数波动较小。池塘水体中的米氏凯伦藻细胞密度与活性磷酸盐含量极显著相关,但与无机氮含量相关性不显著;中肋骨条藻、锥状斯克里普藻细胞密度与氮磷含量相关性均不显著。
