BMP8B对人牙周膜干细胞分化增殖的影响
2023-02-01吴帅楠李召宝王建琪郭香君曹素敏林秀雅褚亚辉
吴帅楠, 李召宝, 王建琪, 郭香君, 曹素敏, 林秀雅, 褚亚辉
(河北省沧州市中心医院口腔门诊, 河北 沧州 061000)
牙周炎是一种高度流行的慢性感染性疾病,可造成牙槽骨、牙周膜和牙骨质的损坏,引起牙周附着丧失,牙龈萎缩,最终导致牙齿松动和脱落[1]。牙周炎的治疗不仅包括局部炎症的控制,还包括牙周组织的再生及重建其功能。干细胞疗法是通过移植具有特异性修复组织潜能的细胞使有缺陷组织再生的一种方法[2]。人牙周膜干细胞(periodontal ligament stem cells,PDLSCs)是牙周组织中的一组间充质干细胞(Mesenchymal stem cells,MSCs),可分化为牙骨质形成细胞和成骨细胞等,是牙周组织再生的关键细胞。因此,研究PDLSCs成骨分化的机制有助于牙周组织再生的治疗和研究。骨形成蛋白(Bone morphogenetic protein,BMP)是一种生长因子,包括BMP2、BMP3、BMP4、BMP7、BMP8 和 BMP9等多种BMP,在骨诱导和骨维持中有重要作用,其有助于间充质细胞向软骨细胞和成骨细胞的增殖和分化[3]。BMP8B是其中重要一种,是骨代谢中重要的信号分子,能够调节机体的产热和能量平衡[4],本研究通过体外分离并培养PDLSCs细胞,并转染BMP8B,探讨BMP8B沉默或过表达对PDLSCs细胞成骨分化的影响。
1 材料与方法
1.1药品与试剂:α-MEM培养基(32561037),美国Gibco公司;总RNA提取试剂盒(AM1830),美国Thermo Fisher Scientific;L-抗坏血酸(A4403)、β-甘油酸钠(G9422)、地塞米松(D4902)、逆转录试剂盒(5091284001)、十二烷基硫酸钠(SDS,71725)、茜素红染色试剂盒(A5533),美国Sigma-Aldrich;实时荧光定量PCR试剂盒(216213),德国QIAGEN;兔源一抗BMP8B(P34280),苏州百远生物;Osterix(ab229258)、骨钙蛋白(Osteocalcin,OCN)(ab13420)、Runt相关转录因子2(Runt - related transcription factor 2,Runx2)(ab76956)、单抗细胞周期蛋白D1(CyclinD1,ab40754)、周期蛋白依赖性激酶2(cyclin-dependent kinases 2,CDK2)(ab32147)、GAPDH(ab181602)、山羊抗兔IgG H&L(ab150079),英国abcam;CCK-8细胞增殖检测试剂盒(C0038)、细胞蛋白提取试剂盒(P0033)、BCA蛋白定量试剂盒(P0012)、碱性磷酸酶(alkaline phosphatase,ALP)活性检测试剂盒(P0321S);4',6-二脒基-2-苯基吲哚(4',6-diamidino-2-phenylindole,DAPI)(C0060),北京索莱宝;BMP8B、Osx、OCN、Runx2 mRNA、BMP8B-shRNA、BMP8B-shRNA阴性对照、GAPDH,上海吉玛制药技术有限公司。荧光倒置显微镜,日本Olympus;5424台式高速离心机,德国Eppendorf;多功能酶标仪、Applied Biosystems PCR仪、蛋白凝胶成像仪,美国Thermo Fisher Scientific。
1.2方 法
1.2.1PDLSCs分离与培养:2019年3月至2019年11月,收集河北省沧州市中心医院口腔科的10例需正畸治疗患者的第一、第二前磨牙,患者年龄为18~25岁,且无已知的牙周疾病,无吸烟史,无系统性疾病,近6个月无特殊服药史。本研究经本院伦理委员会批准。从牙根中部1/3处分离牙周膜组织,加入I型胶原酶和分散酶II,37℃下消化1h,过滤细胞并获得PDLSCs单细胞悬液,将细胞加至含有10%胎牛血清的α-MEM培养基,在37℃、5% CO2培养箱中培养,每3天更换一次培养基,直到细胞从组织块中长出。当接近90%融合时,进行传代培养,第三代细胞用于后续一系列实验。
1.2.2集落形成试验:将1000个PDLSCs接种到直径为10cm的含10%胎牛血清的培养皿中。10d后,用4%多聚甲醛固定细胞,0.1%结晶紫染色,检测细胞克隆情况,若50个或更多细胞的聚集体被认为是细胞克隆。
1.2.3流式细胞术分析:将第三代PDLSCs培养在含有2%胎牛血清和磷酸盐的FACS缓冲液中。然后加入以下抗体:MSC阳性混合物(抗CD90-FITC、抗CD105-PerCP-Cy5.5、抗CD73-APC、抗CD44-PE)和PE阴性混合物(抗CD34-PE、抗CD45-PE、抗CD19-PE、抗CD11b-PE、抗HLA-DR-PE)。在此步骤之后,用流式细胞仪检测标记细胞的信号,并用对数据进行分析。
1.2.4PDLSCs细胞的成骨潜能和成脂诱导的检测:成骨潜能检测:将密度为2×105个/孔的PDLSCs接种于6孔板中,用成骨诱导培养基(含10%胎牛血清、50μg/mL维生素C、10 nmoL/L地塞米松、10mmoL/lβ-甘油磷酸酯)进行成骨诱导培养。培养基每3天更换一次。培养28d后,用4%多聚甲醛固定30min,然后用2%茜素红染色。成脂诱导检测:在成脂诱导培养基(含10%胎牛血清、0.2mM消炎痛、2μM地塞米松、0.5mM 3-异丁基-1-甲基黄嘌呤、0.01mg/mL胰岛素的α-MEM培养基)中培养2×105个PDLSCs/孔。28d后,细胞用油红O染色并在显微镜下观察。
1.2.5细胞分组及处理:转染前24h,收集上述1.2.1中对数期PDLSCs细胞,以1×105个/孔细胞浓度接种至含血清培养基的24孔板中。每孔细胞浓度为2×105个/孔时,更换为不含血清的新鲜培养基,之后进行慢病毒感染,将细胞分为对照组(仅添加新鲜培养基)、阴性转染组(感染si-NC)、BMP8B沉默组(转染BMP8B shRNA)和BMP8B过表达组(转染BMP8B siRNA),继续培养48h。
1.2.6CCK-8法检测细胞增殖情况:收集1.2.2中各组细胞,以6×105个/mL的密度接种于96孔板中,用含0.5%胎牛血清的DMEM培养基在37℃、5% CO2培养箱中培养48h。每孔加入10μL CCK-8溶液,37℃培养3h,用酶标仪测定450nm处的吸光度。
1.2.7ALP活性的检测:收集1.2.2中各组细胞,以6×105个/mL的密度接种于96孔板中,然后用含0.5%胎牛血清的DMEM培养基培养48h。然后用PBS洗涤,并用100μL裂解缓冲液(0.2% TritonX100)在冰上孵育2h。收集30μL裂解液进行ALP活性检测。将30μL裂解液与20μL buffer和50μL底物在96孔板中37℃孵育10min。然后,加入100μL终止液,用酶标仪测定405nm处的吸光度。
1.2.8茜素红染色法检测成骨细胞矿化结节:将各组PDLSCs细胞浓度调整为1×105个/mL接种于6孔板中,细胞融合度达80%时,更换为含10%胎牛血清、抗坏血酸(50μg/mL)、β-甘油酸钠(10mM)、地塞米松(10nM)诱导培养基,37℃、5% CO2培养箱中培养21 d,弃培养液,PBS洗涤,95%酒精固定,1%茜素红染料室温孵育30min,PBS洗涤,随机选取6个视野,显微镜下观察细胞,矿化结节呈亮红色。400μL 10%的十六烷基吡啶氯化铵加入至10mM磷酸钠溶液中萃取PSLSCs中的茜素红染料,使茜素红染料从细胞质基质中释放,10min后,560nm处分光光度计测量PSLSCs细胞释放的茜素红,并进行定量分析。
1.2.9实时荧光定量聚合酶链反应(Quantitative Real-time PCR,qRT-PCR)检测BMP8B、Osterix、OCN、Runx2 mRNA表达水平:RNA提取试剂盒提取细胞RNA,并进行反转录,qRT-PCR扩增。反应条件为预变性 95℃ 60s;变性 95℃ 30s,退火60℃ 45s;延伸72℃ 30s共40个循环。以GAPDH为内参,2-ΔΔCt法计算BMP8B、Osterix、OCN、Runx2 mRNA的相对表达量。引物序列见表1。

表1 引物序列
1.2.10蛋白免疫印迹(western blot,WB)法检测蛋白表达:提取1.2.5中诱导培养的各组细胞总蛋白并进行浓度检测,蛋白样品上样到10%十二烷基硫酸钠-聚丙烯酰胺(SDS-PAGE)凝胶电泳上,电泳结束后印迹到PVDF膜上,5%脱脂牛奶封闭;分别加入一抗稀释液(BMP8B、Osterix、OCN、Runx2、CyclinD1,均为1∶1000稀释、CDK2为1∶5000稀释),4℃孵育过夜,TBST洗膜3次,加入山羊抗兔IgG H&L二抗稀释液(1∶2000),室温孵育1h,TBST洗膜3次。GAPDH蛋白为内参,蛋白凝胶成像仪分析蛋白表达。
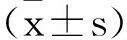
2 结 果
2.1PDLSCs细胞鉴定:显微镜下观察PDLSCs细胞为单核、多边形或梭形(图1A),茜素红染色结果显示显微镜下可见红色染色的矿化结节,证明PDLSCs具有成骨潜能(图1B)。油红O染色结果显示有红色的脂滴形成,证明PDLSCs具有成脂潜能(图1C)。此外,克隆形成实验表明,获得的细胞具有形成克隆的能力(图1D,E)。流式细胞仪分析显示,PDLSCs的MSC标记(CD90、CD44、CD105、CD73)呈阳性表达,而造血标记(CD34、CD11b、CD19、CD45、HLA-DR)呈阴性表达(图2)。

图1 PDLSCs细胞鉴定注:A:PDLSCs的形态学特征;B:茜素红染色;C:油红O染色;D:结晶紫染色;E:PDLSCs的单个克隆的代表性图像

图2 PDLSCs的流式结果图
2.2转染后各组PDLSCs中BMP8B mRNA的表达:与对照组和阴性转染组相比,BMP8B沉默组细胞BMP8B mRNA的表达显著降低(P<0.05),BMP8B过表达组细胞BMP8B mRNA的表达显著升高(P<0.05)。见表2。

表2 转染后各组PDLSCs中BMP8B mRNA的表达(n=9)
2.3BMP8B对PDLSCs细胞增殖的影响:与对照组和阴性转染组相比,BMP8B沉默组细胞增殖情况显著降低(P<0.05),BMP8B过表达组细胞增殖情况显著升高(P<0.05)。见表3。

表3 BMP8B表达对细胞PDLSCs细胞增殖的影响(n=9)
2.4BMP8B对PDLSCs细胞ALP活性的影响:与对照组和阴性转染组相比,BMP8B沉默组细胞ALP活性显著降低(P<0.05),BMP8B过表达组细胞ALP活性显著升高(P<0.05)。见表4。

表4 BMP8B对PDLSCs细胞ALP活性的影响(n=9)
2.5BMP8B对PDLSCs细胞钙化结节的影响:与对照组和阴性转染组相比,BMP8B沉默组细胞茜素红染色程度显著降低(P<0.05),BMP8B过表达组细胞茜素红染色程度显著升高(P<0.05)。见表5、图3。

图3 BMP8B对PDLSCs细胞钙化结节的影响注:A:对照组;B:阴性转染组;C:BMP8B沉默组;D:BMP8B过表达组

表5 BMP8B对PDLSCs细胞钙化结节的影响(n=9)
2.6BMP8B对PDLSCs细胞中成骨标记蛋白Osterix、OCN、Runx2 mRNA表达的影响:与对照组比较和阴性转染组相比,BMP8B沉默组细胞Osterix、OCN、Runx2 mRNA表达显著降低(P<0.05),BMP8B过表达组细胞Osterix、OCN、Runx2 mRNA表达显著升高(P<0.05),见表6。

表6 BMP8B对PDLSCs细胞中成骨标记Osterix OCN Runx2 mRNA表达的影响(n=9)
2.7BMP8B对PDLSCs增殖、成骨蛋白表达的影响:与对照组比较和阴性转染组相比,BMP8B沉默组细胞细胞CyclinD1、CDK2、Osterix、OCN、Runx2蛋白表达显著降低(P<0.05),BMP8B过表达组细胞CyclinD1、CDK2、Osterix、OCN、Runx2蛋白表达显著升高(P<0.05),见图4、5。

图4 蛋白相对表达水平的比较注:与对照组比较,*P<0.05;与阴性转染组比较,#P<0.05

图5 BMP8B对PDLSCs增殖、成骨相关蛋白表达的影响注:A:对照组;B:阴性转染组;C:BMP8B沉默组;D:BMP8B过表达组
3 讨 论
牙周炎是成人最常见的口腔疾病之一,也是牙齿损坏和掉落的主要原因之一,牙周炎会增加患者患心血管疾病等的风险[5]。其发病机理主要是由于细菌或其产物聚集在牙根或牙周组织表面,侵入牙周组织导致慢性炎症,最终引起牙齿脱落。牙周炎治疗最终目标是恢复牙周组织的结构和再生功能,应用有利的种子细胞再生受损组织可能是牙周炎治疗的有效方法[6]。PDLSCs具有分化为多种细胞类型并进行稳健的克隆的自我更新的潜力,在成骨相关激素、细胞因子或其他诱导剂作用下,PDLSCs可分化为骨原细胞、成骨细胞、成骨细胞前体细胞等,被认为是牙周组织再生治疗中的潜在干细胞群体。研究与PDLSCs增殖和分化有关的分子机制可能为PDLSCs细胞的成骨分化机制研究提供理论基础。本研究对PDLSCs细胞进行分离与培养,经流式细胞术分析及成骨潜能和成脂诱导的检测后表明,所分离细胞是PDLSC,具有干细胞特性。
BMP是转化生长因子β(transforming growth factor β,TGF-β)家族成员,最初被鉴定为骨提取物中的骨诱导成分,BMP信号传导是MSCs细胞成骨分化和骨形成调节的重要途径之一[7],研究发现TGF-β家族成员BMP2具有促进PDLSCs等细胞成骨分化的作用[8]。增殖是细胞分化的基础,本研究结果显示BMP8B过表达能够促进PDLSCs的增殖,沉默则抑制细胞增殖。ALP在细胞外基质矿化过程中有重要的作用,是成骨细胞成熟的标志物之一[9]。根据实验结果进一步显示,BMP8B沉默或过表达均会影响细胞ALP活性和钙化程度,表明BMP8B可能与细胞的PDLSCs增殖和成骨分化有关,具体机制有待进一步研究。Runx2、Osterix通常被认为是早期成骨分化的标志,Runx2是MSCs向成骨细胞分化过程中的关键转录因子,属于runt结构域基因家族的转录因子,参与不同阶段成骨细胞分化成熟过程[10]。Runx2通过参与调控成骨细胞分裂周期和ALP、OCN、I型胶原等成骨细胞分化成熟表型标志物的表达并启动矿化[11]。Osterix是Runx2的下游信号传导基因,也是是成骨分化过程必不可少的转录因子。已有研究发现,BMP2/Runx2/Osterix信号通路通过参与成骨分化早期阶段,促进MSC的成骨分化[12]。OCN是由成骨细胞产生的骨骼中最丰富的蛋白质之一,主要在矿化过程中产生,被认为是成骨细胞成熟的标志[13]。本研究qRT-PCR及WB实验结果表明沉默BMP8B会降低Osterix、OCN、Runx2表达,BMP8B过表达则促进Osterix、OCN、Runx2水平,提示BMP8B可能通过影响成骨分化过程中相关转录因子进而参与成骨细胞分化的过程,具体机制还需进一步的研究。
综上所述,BMP8B能够影响PDLSCs细胞增殖和分化,BMP8B可能是成骨分化过程中的必要生物因子之一,是牙周炎治疗的潜在研究方向。本研究仅探讨了BMP8B沉默或过表达对体外细胞的影响,下一步需结合体内实验,进一步探讨BMP8B在成骨分化中的可能作用和相关机制。
