Insights into the relations between cell wall integrity and in vitro digestion properties of granular starches in pulse cotyledon cells after dry heat treatment
2023-01-21PingLiBinZhngRuiLiuLiDingXiongFuHitngLiQingHungXiowi
Ping Li,Bin Zhng,Rui Liu,Li Ding,Xiong Fu,c,Hitng Li,Qing Hung,c,Xiowi H,d
a School of Food Science and Engineering,Guangdong Province Key Laboratory for Green Processing of Natural Products and Product Safety,South China University of Technology,Guangzhou 510640,China
b Sericultural&Agri-Food Research Institute Guangdong Academy of Agricultural Sciences/Key Laboratory of Functional Foods,Ministry of Agriculture and Rural Affairs/Guangdong Key Laboratory of Agricultural Products Processing,Guangzhou 510610,China
c Overseas Expertise Introduction Center for Discipline Innovation of Food Nutrition and Human Health (111 Center),Guangzhou 510640,China
d Institute of Food and Nutrition Development,Ministry of Agriculture and Rural Affairs,Beijing 100081,China
e Center for Nutrition and Food Sciences,The University of Queensland,St Lucia,QLD 4072,Australia
Keywords:Cell wall integrity In vitro starch digestion Pulse Dry heat treatment
ABSTRACT Natural foods,such as whole pulses,are recommended in the dietary guidelines of the US and China.The plant cell wall structure in whole pulses has important implications for the nutritional functionalities of starch.In this study,garbanzo bean cells with varying degrees of cell wall integrity were subjected to dry heat treatment (DHT) and used to elucidate the food structure-starch digestion properties of pulse food.The morphological features suggested that all cell samples do not exhibit remarkable changes after being subjected to DHT.Molecular rearrangement and the crystallite disruption of starch granules entrapped in cells occurred during DHT as assessed by the crystal structure and thermal properties.DHT decreased the inhibitory effects of enzymes of both the soluble and insoluble components,but the digestion rate and extent of slightly and highly damaged cell samples did not exhibit significant differences compared with their native counterparts.We concluded that the starch digestion of pulse cotyledon cells is primarily determined by the intactness of the cellular structure.This study reveals the role of food structure on the ability to retain the desirable nutritional properties of starch after subjection to physical modification.
1.Introduction
Whole pulses exhibit a remarkably low glycemic response among carbohydrate-rich foods,which is beneficial to control metabolic disorders such as diabetes [1-4].As the main glycemic carbohydrate in whole pulses,starches are enclosed by thick cotyledon cell walls that hinder enzyme diffusion under bothin vitro[5]andin vivoconditions [6].After mechanical or thermal disruption of the cellular structure,digestive enzyme susceptibility is greatly promoted to liberate the nutritional components.A wide range of food processing techniques,such as boiling and frying,has been applied to investigate thein vitrostarch digestion of pulse cotyledon cells [7-10].Pallares et al.[11]reported that different intensities of thermal treatment resulted in cell wall permeability discrepancies,thus affecting thein vitrostarch digestion.Our previous study also found that cell wall permeability resulting from variable processing greatly contributed to the starch digestibility of pulse cells [12].
In addition to limiting enzyme diffusion,intact cellular structure also restricts starch gelatinization in pulse cells during thermal treatment to reduce digestibility when compared with fully gelatinized starches.Additionally,variousα-amylase inhibitors in whole pulses,such as lectins (0.6% -3.0%),phytic acid (1% -3%) and proteinaceous inhibitors,contribute to reducing the digestion of starch in pulses [13,14].Generally,the decreased bioavailability of starch in pulses is a consequence of multiple factors working concurrently,and the cell walls would act as a key characteristic to cause the slow digestion of starch in whole pulses.
Despite intensive researches have evaluated the digestion of starch entrapped in pulse cotyledon cells [6,15,16],the impact of cell wall modifications on starch hydrolysis is really limited.Most information is focused on the starch behaviors being subjected to physical (hydrothermal) breakdown of the cellular or granular structure.The contribution of cell wall to starch digestion in pulses has not been conclusively assessed,especially for the assessment of cell wall integrity on starch digestive behavior.In our previous study,we have investigated the digestibility of several commercially available pulses (e.g.,pinto bean,garbanzo bean and black-eyed pea et al.),and found that the garbanzo bean cells exhibited relatively lower digestion properties [17].Thus in the present study,we chose garbanzo beans as representative to investigate the starch digestion mechanism in pulses.Intact cells isolated under pulse starch gelatinization temperatures were set as the whole pulse food model.Different degrees of cell wall integrity were obtained via controlled cell wall modifications,following with dry heat treatment (DHT) to simulate baking conditions.Structural features and starch digestion kinetics were investigated to clarify the relationship between cell wall integrity and digestion properties.Additionally,we also assessed the enzyme inhibitory effects of both the soluble and insoluble components in the pulses after DHT to understand starch digestion behaviors more comprehensively.The present work will provide a theoretical basis for designing the next generation of whole food products and functional ingredients from pluses.
2.Material and methods
2.1 Materials
Garbanzo bean (Cicer arietinum) seeds were obtained at a local supermarket (Guangzhou,China).Pancreatin (Sigma P-1750,from the porcine pancreas),calcofluor white stain (Sigma 18909),viscozyme®L enzyme (Sigma,V2010),4-hydroxybenzoic acid hydrazide (PAHBAH,Sigma H9882),and fluorescein isothiocyanate conjugate tagged dextrans (FITC-dextran,20 kDa) were obtained from Sigma-Aldrich,China.A total starch assay kit (K-TSTA) was purchased from Megazyme (Bray,Ireland).All other chemicals(Na2CO3,NaHCO3,HCl,NaOH,etc.) used in this study were of analytical grade.
2.2 Preparation of intact garbanzo bean cells and starch granules
Intact garbanzo bean cells were prepared according to a slightly modified method [5].Seeds were kept in an admixture solution of Na2CO3(0.5 g/100 mL) and NaHCO3(1.5 g/100 mL) overnight to loosen and remove the outer hulls.Then,dehulled bean seeds(200 g) were incubated at 60 °C for 1 h in 2 L of distilled water.The thermally treated seeds were gently mashed using a mortar and pestle device and rinsed with flowing water passing through a series of 250,150 and 90 μm sieves to obtain intact cell factions.The cells retained on the 90 μm sieves were recovered and freeze-dried.Purified pulse starch was obtained on the basis of the following method [14].
2.3 Preparation of dry heat-treated cell samples with different cell wall integrities
Intact garbanzo bean cells (IC) were incubated with the Viscozyme®L enzyme for different lengths of time [18].Briefly,the pH of intact cell suspensions (10 g/100 mL) was adjusted to 5.0 with hydrochloric acid solution (HCl,0.5 mol/L),followed by the addition of the Viscozyme®L enzyme (50 FBG,0.25 mL) solution.Then,the mixture was incubated at 40 °C for 30 or 60 min without stirring,and was denoted as slightly damaged cells (SDC) and highly damaged cells (HDC),respectively,as confirmed by light microscopy.The cell samples were collected by centrifugation at 2 000 ×gfor 5 min and freeze-dried.
Broken cells (BC) were prepared by the constant magnetic stirring of HDC,confirmed by light microscopy,and consequently recovered as described [5].
All cell and isolated starch samples were oven-heated at 160 °C for 2 h.These heated samples were then cooled to room temperature in a desiccator for further use.
2.4 Microscopic observations
Light microscopy of the cell and starch samples was observed using a BX-51 microscope (Olympus,Japan).Samples were dispersed in deionized water (0.1%,wt),and then a drop of suspension was transferred to a glass slide followed with covering by a glass cover slip.The microscope pictures were captured at 200 × magnifications.
ZEISS EVO18 (Germany) equipment was used to observe the scanning electron microscopy pictures.Samples were mounted onto a metal stub covered with double-sided tape to coat with a thin film of gold.The pictures were acquired at an accelerating voltage of 20 kV at 500 × magnifications.
2.5 Fluorescein isothiocyanate-dextran diffusion experiment
The fluorescein isothiocyanate-dextran diffusion experiment was conducted following the modified report [12].Briefly,cell samples were suspended in fluorescein isothiocyanate-dextran solution (20 kDa,1 mg/mL) at 4 °C in the dark overnight.Then,the cells were transferred onto a microscope slide followed by the addition of calcofluor white staining solution before observation.The fluorescence emitted was collected at the excitation wavelengths of 488 nm (FITC) and 405 nm (calcofluor white).A confocal microscope(TCS-SP5,Leica,Germany) was used to concurrently observe both the cell wall permeability and starches.
2.6 X-ray diffraction (XRD)
XRD of the cell and starch samples was performed at 40 kV and 30 mA with CuKα radiation (λ=0.154 nm) using a D8 Advance diffractometer (Bruker,Germany).Diffractograms were obtained in the 2θrange of 4° to 40° with a scan rate of 0.5°/min at atmospheric temperature.The crystalline and amorphous areas were separated by PeakFit software (Version 4.0,Systat Software Inc.,San Jose,CA,USA) following the method [19].The percent of the crystalline peak area to the total diffraction area was determined to be the relative crystallinity.
2.7 Thermal properties
A differential scanning calorimeter 3 (Mettler Toledo,Switzerland) was employed to determine the thermal behaviors of garbanzo bean cell and starch samples.Cells and starches (~3 mg)were precisely weighed into a standard aluminum pan with distilled water (~7 mg) and hermetically sealed to equilibrate for 4 h at atmospheric temperature.The samples were heated to 120 °C at a rate of 10 °C/min.The temperatures of onset (To),peak (Tp),and conclusion (Tc) and the enthalpy change (ΔH) were calculated by STARe evaluation software.
2.8 Inhibition of enzymatic activity of the soluble and insoluble components
The water-soluble components inside garbanzo bean cells were isolated to determine the inhibitory effects on the activity of pancreatic amylase based on previously reported modified methods [20].Briefly,cells were mixed with deionized water (1 g/100 mL) for incubation at 37 °C for 12 h.The mixture was then centrifuged(4 000 ×g) for 20 min to collect the supernatant,following evaluating the inhibitory effects on the enzymatic activity of the water-soluble components.Specifically,the supernatant (250 μL) was mixed with pancreatin solution (250 μL) and incubated at 37 °C for 5 min.Then,this mixture was used to determine theα-amylase activity following methods described in detail elsewhere [21].Theα-amylase activity was calculated according to the slope of the linear least-squares fit of the hydrolysis curve of maize starch (Sigma) into reducing sugar.
The water-insoluble proteinaceous component was separated according to the modified method [22].Generally,dehulled pulse seed flour was soaked in distilled water (10 g/100 mL) and the pH value of the solution was adjusted to 9.0 with sodium hydroxide solution(NaOH,1.0 mol/L).The cell suspension was magnetically stirred at 1 000 r/min for 60 min,and the supernatant was separated by centrifugation at 5 000 ×gfor 20 min.The proteinaceous component in the supernatant was precipitated by pH adjustment to 4.6 with hydrochloride solution (1.0 mol/L).The precipitate was collected by centrifugation at 5 000 ×gfor 20 min,followed by washing with distilled water and oven-drying.
The water-insoluble component was subjected to DHT and then cooled to atmospheric temperature in a desiccator.Approximately 30 mg of the insoluble component was suspended in phosphatebuffered saline (PBS,10 mL),followed by incubation with pancreatin at 37 °C for 5 min.After centrifugation (4 000 ×g) of the resulting mixture for 1 min,the residual enzymatic activity of the supernatant was examined following the above methods.
2.9 In vitro starch digestion kinetics
The procedure for thein vitrodigestion of the garbanzo bean cell and starch samples was adapted from a method reported previously [12].Samples (100 mg,dsb) were dispersed in 10 mL of PBS with 200 units of pancreatin at 37 °C without magnetic stirring.Tubes were shaken gently during the hydrolysis process.At predetermined time intervals from 0 to 180 min,50 μL aliquots were immediately transferred to 250 μL of ice-cold Na2CO3solution (0.5 mol/L) to terminate the catalytic process followed by centrifugation (2 200 ×g)to separate the undigested samples.The end product (reducing sugar)concentration in the supernatant was analyzed based on the PAHBAH method.The maltose (reducing sugar) equivalent released during sample digestion was calculated using the following equation:
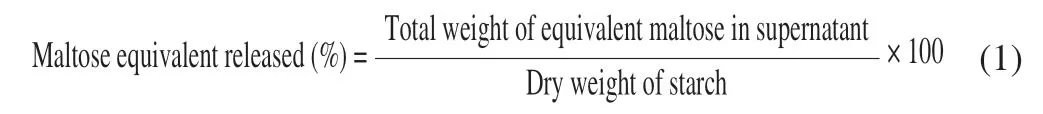
The reducing sugar profile was then fitted to the following first-order equation to calculate the apparent digestion rate coefficient (k,min-1) [23].

wheretis the digestion time (min),Cis the amount of digested starch at digestion timet,andkis the apparent digestion rate coefficient (min-1).
2.10 Statistical analysis
Experiments were conducted with duplicate or triple measurements,and the results were analyzed by the least significant difference (LSD) test with a significance level ofP<0.05.
3.Results and discussion
3.1 Morphological changes in the cell and starch samples subjected to DHT
The pectin networks among pulse cotyledon cells could be softened by soaking in a solution of Na2CO3and NaHCO3.Intact cells could then be isolated by thermal treatment under the gelatinization temperature of starch (60 °C) to protect the intactness of its cell wall and crystal structure [24].As Figs.1 and 2 show,most isolated pulse cells remained intact with negligible broken cells or free starch granules (Figs.1 and 2,IC).Generally,the IC was oval or elongated in shape with a particle size that varied between 150-200 μm(Fig.1,IC).The IC exhibited a wrinkled surface (Fig.2,IC),which could be attributed to the freeze-drying process [15].The starch granules in the IC showed obvious birefringence intensity as observed by polarized light microscopy.After incubation with the cell wall degradation enzyme,the cell walls gradually dissolved into fragments with an increase of the leaching of starch granules (Fig.1,SDC and HDC).However,most of the starch granules were still encapsulated by the cell membrane even after subjection to prolonged cell wall degradation (Fig.1,HDC).After being subjected to mechanical force,the hydrolyzed cells were totally broken with scattered protein and starch components (Fig.1,BC).No obvious changes in birefringence intensity were observed among the cell samples treated with Viscozyme®L.Additionally,the birefringence intensity of the cell and starch samples did not show a remarkable decrease after DHT,indicating retention of the crystal structure.Notably,under microscopic observation,the protein components of the totally BC samples formed large lumps (Figs.1 and 2,BC-DHT) due to protein denaturation during DHT.
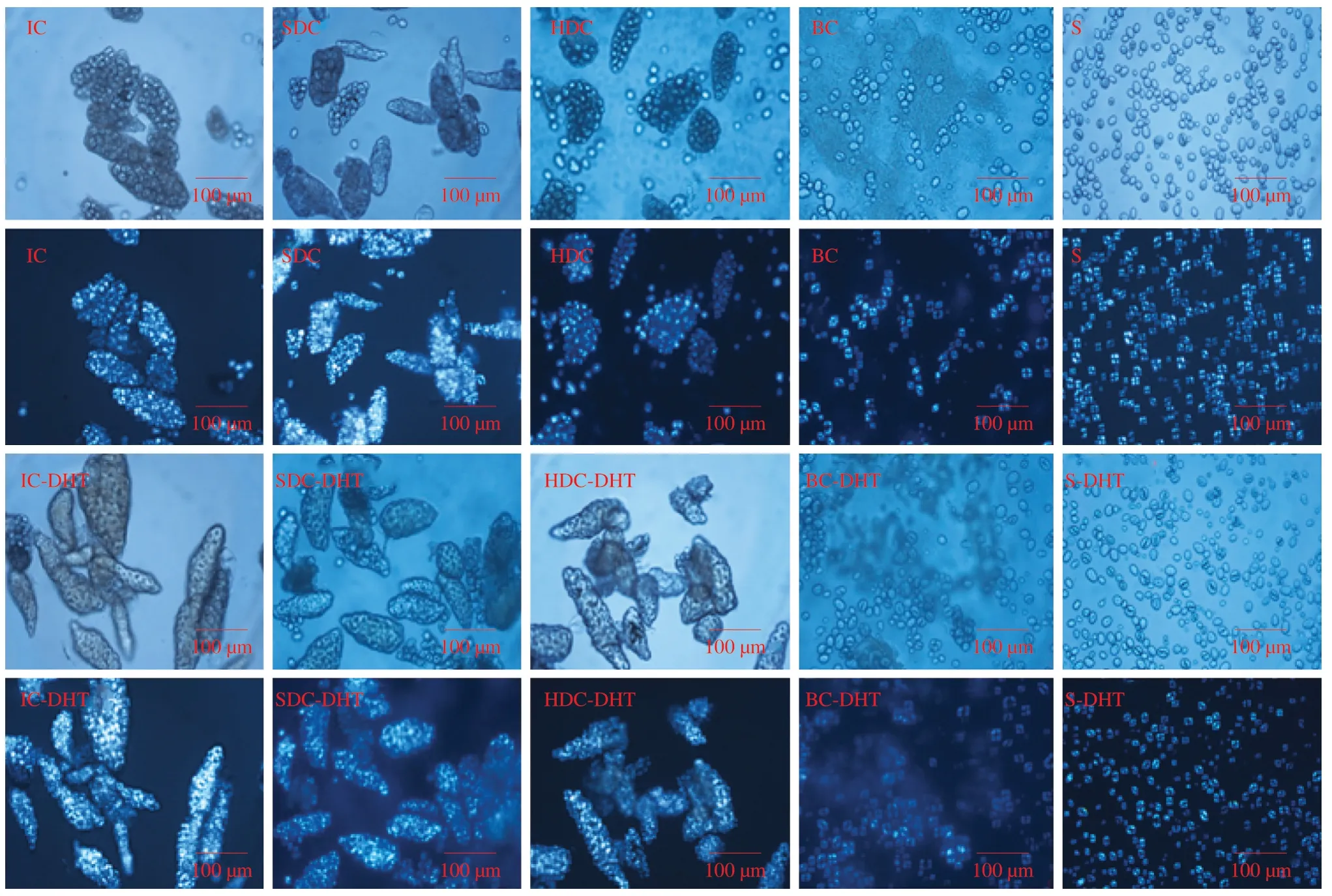
Fig.1 Light micrograph pictures of isolated garbanzo bean cells and starches before and after dry-heat-treatment.(S,starch;scale bar=100 μm)
To visualize the permeability of cell wall structure,pulse cells samples were labelled with calcofluor-white (specific to cell wall polymers,blue fluorescence in Fig.2) and FITC-dextran (green fluorescence in Fig.2).A 20 kDa molecular weight fluorescent FITC-dextran probe was used to assess the permeability of the cell structure to the digestive enzyme,as the FITC-dextran probe has a hydrodynamic size that is similar to that ofα-amylase [25].As shown in Fig.2,negligible fluorescence (green) was observed within the IC samples,whereas noticeable fluorescence (green) was detected within the damaged cell samples (Fig.2,SDC and HDC).This result could be attributed to cell wall degradation,which was also observed from calcofluor white labeling (blue fluorescence). More fluorescence(green) inside the cells would indicate the higher permeability of cell structure.Similar evaluation method has also been used to investigate the cell wall permeability in our previous study.Alteration of cell wall permeability induced by different processing conditions is evaluated through the diffusion of fluorescence-tagged dextran probes.The double labeling of the FITC-dextran probe and calcofluor white could determine the micron-scale integrity of the plant cell wall[26].The structural features (folds and ruptures) could be highlighted with calcofluor white labeling,which are invisible when subjected to bright-field microscopy without labeling.When subjected to DHT,all cell samples (IC-DHT,SDC-DHT and HDC-DHT) showed a remarkably higher degree of penetration from the FITC-dextran probe,suggesting increased permeability of the cellular structure.
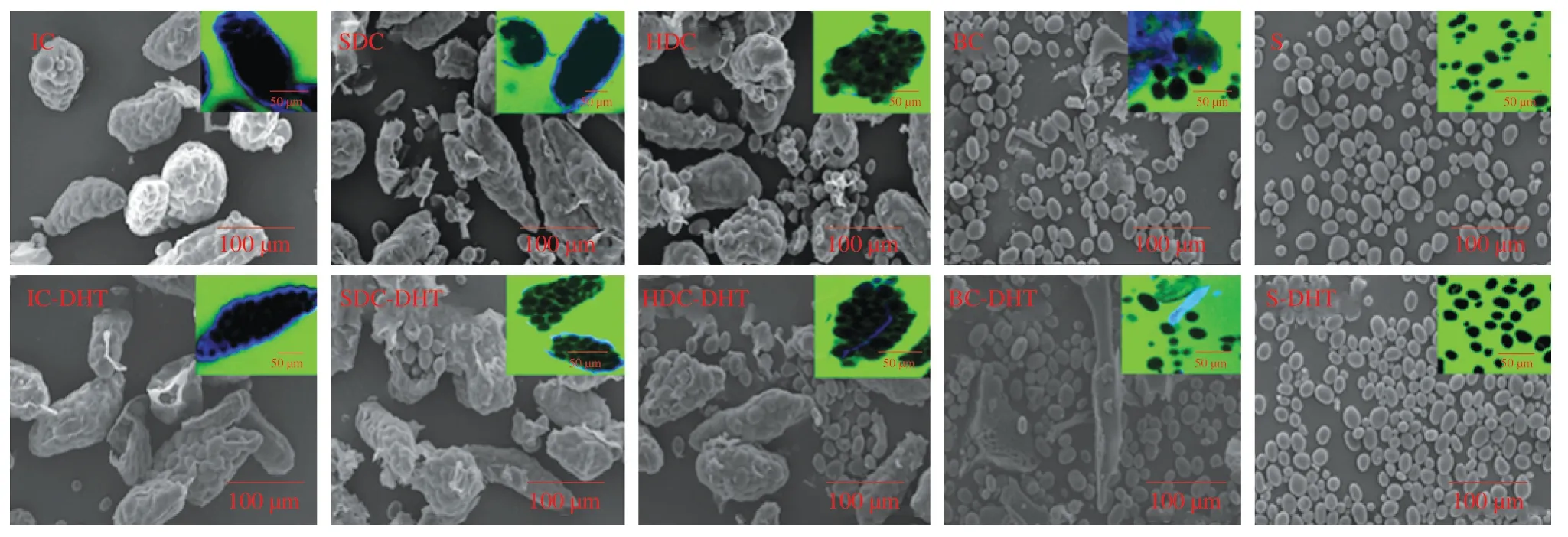
Fig.2 Scanning electron micrographs and confocal light scanning micrographs of garbanzo bean cells and starches before and after dry-heat-treatment.Scale bar in scanning electron micrographs is 100 μm,and scale bar in confocal light scanning micrographs is 50 μm.
3.2 Crystalline structure properties
Fig.3 summarizes the X-ray diffraction profiles of the garbanzo bean cells with different cell wall integrities before and after DHT.All of the cell and starch samples before DHT exhibited a classic C-type pattern with main peaks at~5.03° and 15.2°,17.2°,18.1°,and 23.4°2θ[27,28].However,the peak at approximately 5.03° of the B-type characteristic polymorph is hardly discernible after freeze-drying processing,as reported before [29].With increased degradation of the cell wall barriers,the C-type polymorphic pattern of the cell samples(Fig.3A,SDC,HDC and BC) became sharper compared with their native counterparts (Fig.3A,IC).However,no obvious changes in the relative crystallinity (labelled in Fig.3) were observed among the cell samples with different degrees of cell integrity,which is consistent with the microscopic results.
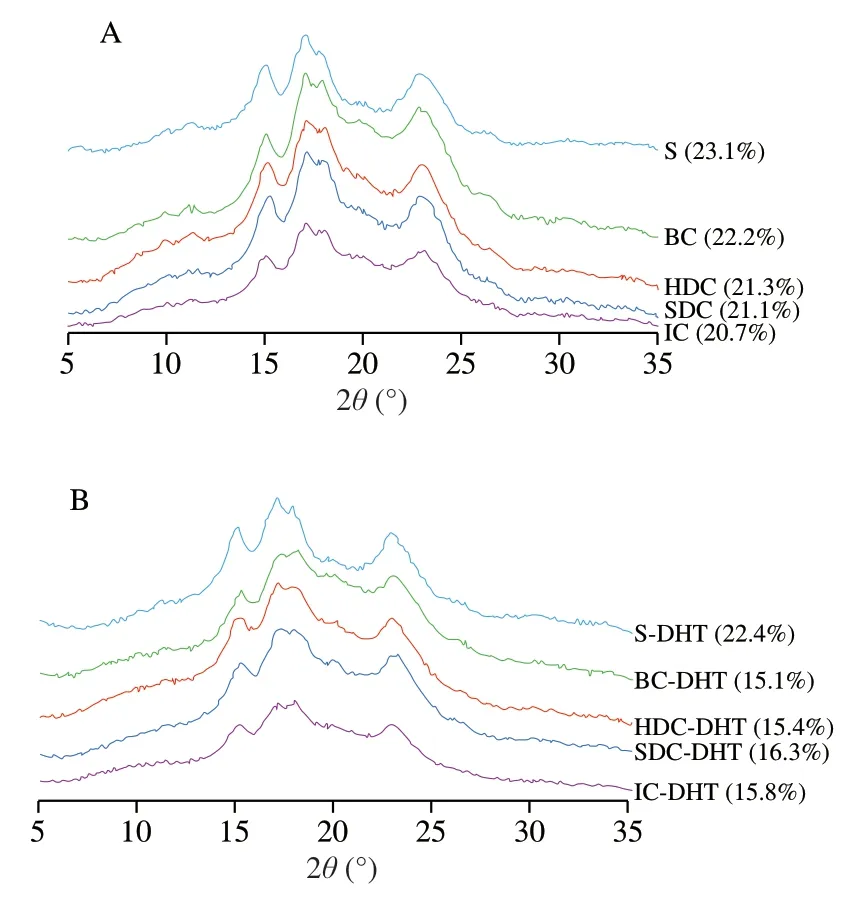
Fig.3 X-ray diffractograms of garbanzo bean cells and starches before (A)and after (B) dry-heat-treatment.The percent of relative crystallinity of cell and starch samples was labelled after the sample code.
All cell and starch samples subjected to DHT maintained a C-type crystalline structure.However,the diffraction peaks became weaker and broader as the relative crystallinity decreased (Fig.3B).Cell samples with lower cell wall integrity exhibited a larger decrease in crystallinity,whereas pure starches samples did not show obvious changes in crystallinity.Specifically,BC-DHT samples (Fig.3B,BC-DHT,15.1%) exhibited an obvious decrease (approximately 7%)in relative crystallinity when compared with their BC counterparts(Fig.3A,BC,22.2%),whereas other cell samples only exhibited approximately 5% decrease (calculated by the relative crystallinity shown in Fig.3) in crystallinity.This result could be possibly attributed to the increased exposure to heat without cellular structure encapsulation during DHT.It is worth noting that no obvious changes in birefringence intensity were observed among the cell and starch samples compared with their native counterparts.Thus,we hypothesize that crystalline structure disruption and starch molecule rearrangement may simultaneously occur upon subjection to DHT,as the double-helical chains of starch granule may increase to disrupt the starch crystallites and/or change the crystallite orientation [30].In addition,partial melting of imperfect crystals from the crystalline region may also result in a decrease in crystallinity [31].
3.3 Thermal properties of the cell and starch samples subjected to DHT
Table 1 presents the thermal behaviors of the cell and starch samples with different cell wall integrities before and after DHT.All cell samples exhibited higher gelatinization temperatures and lower enthalpy changes than the pure starches (Table 1),which is consistent with our previous reports [12,17].IC exhibited the highest onset gelatinization temperature (To) and lowest ΔHamong all cell samples before DHT,which was attributed to the delayed gelatinization induced by the limited water absorption into the plant cells.The gelatinization temperatures (To,Tp,andTc) of the cell samples showed a decreasing trend with reduced cell wall integrity,which would promote water ingress as well as swelling space for the starch within pulse cells.
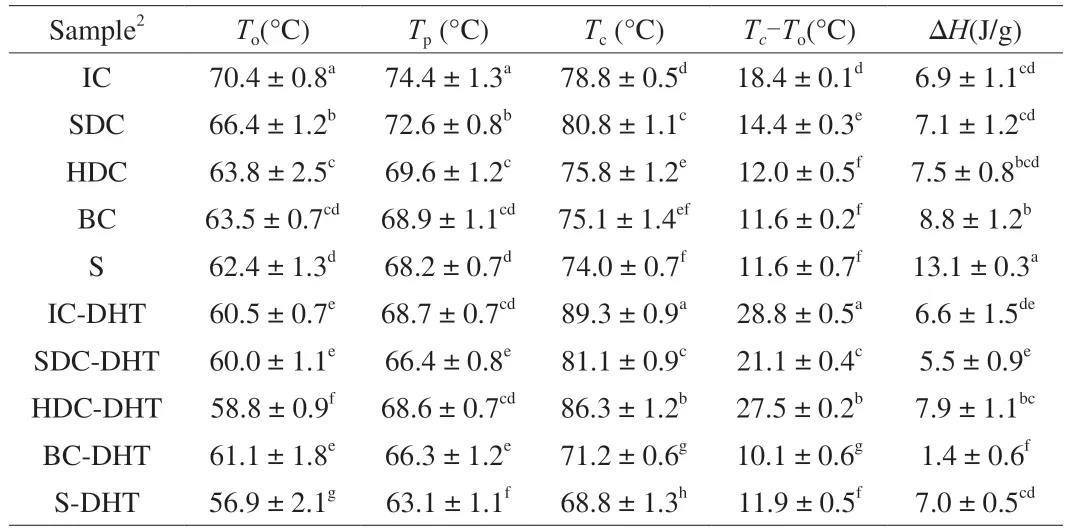
Table 1 Thermal properties of garbanzo bean cells and starches before and after dryheat-treatment1.
After DHT,all cell samples showed a reducedTocompared with their native counterparts,and theTc(IC-DHT,SDC-DHT,HDC-DHT) shifted towards a higher value,resulting in an increase in the gelatinization temperature range (Tc-To).These results could be attributed to the interactions between the amylose and amylopectin chains that occurred during DHT.The enthalpy changes (ΔHvalues)of the cell samples (IC-DHT,SDC-DHT,HDC-DHT) showed no obvious differences compared with their native counterparts(IC,SDC,HDC),whereas the BC samples and pure starches subjected to DHT (BC-DHT,S-DHT) exhibited a remarkable decrease in their ΔHvalues (Table 1).These results indicate that the rearrangement of starch molecules most likely occurs inside cell samples with intact or less damaged cellular structures.The disruption of starch double helices would be further intensified for BC and pure starch samples.
3.4 Inhibitory effects of the soluble and insoluble components on enzymatic activity
Whole pulses contain severalα-amylase inhibitors,including lectins,tannins and proteinaceous inhibitors [13].Thus,we evaluated the inhibitory effects of the soluble and insoluble components of the pulses on the activity of pancreaticα-amylase in the present study,and the result was shown in Fig.4.We supposed the soluble components loss during pulse cells isolation process is minimized to exclude the following influence.The soluble components of the IC(Fig.4A) showed the lowest inhibitory (approximately 5.8%,data not shown) effects on the activity of pancreaticα-amylase.Then the inhibitory effects increased with reduced cell wall integrity,indicating that more soluble components diffused into the system.However,the inhibitory activity of the soluble components decreased dramatically when all the cell samples were subjected to DHT (Fig.4A).This result could be attributed to the inactivation or degradation of the soluble components under high temperature [32,33].
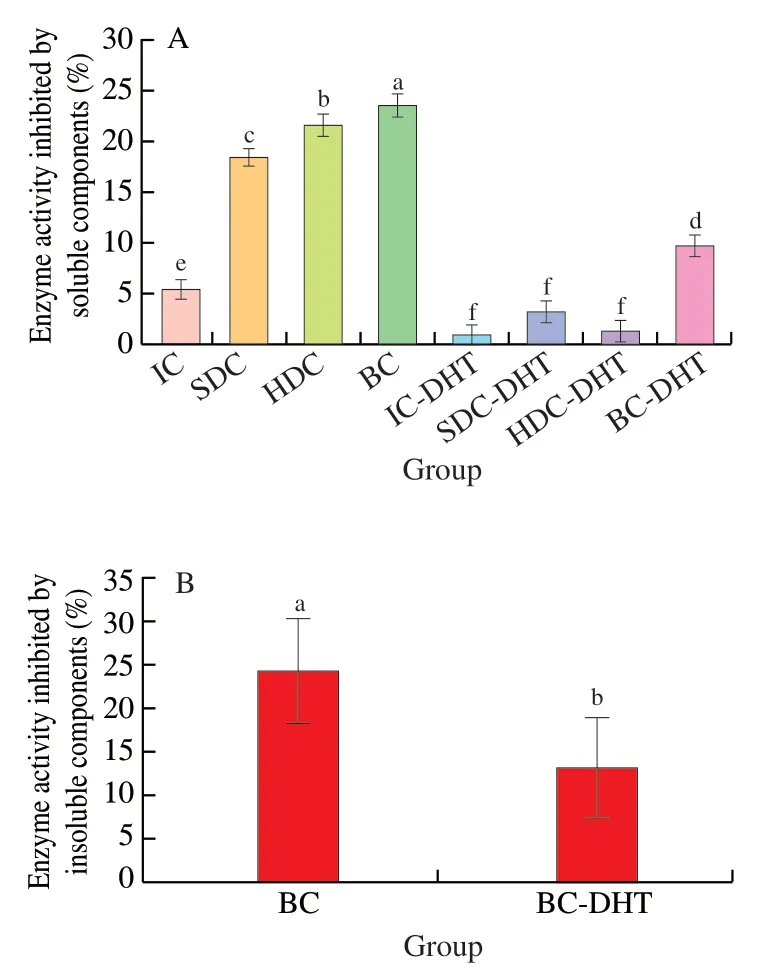
Fig.4 Inhibition of soluble (A) and insoluble components (B) from garbanzo bean cells with different cell wall breakage and starches before and after dry-heat-treatment.
Notably,the inhibitory activity of only the insoluble proteinaceous component isolated from the BC was evaluated due to technical difficulties in removing starch from the samples with different cell wall integrities.As shown in Fig.4B,the inhibitory activity of the insoluble component for BC subjected to DHT decreased from 24.5% (BC) to 13.3% (BC-DHT).This result could be attributed to the denaturation of the insoluble proteinaceous component during the heating process.The insoluble component after DHT also retained a certain amount of inhibitory activity,which was perhaps attributed to thermostable inhibitors,as reported before in wheat [34].
3.5 In vitro digestion kinetics of the cell and starch samples subjected to DHT
The digestion behaviors of the garbanzo bean cell and starch samples subjected to DHT were evaluated without stirring to lessen the degree of cell wall breakage.Samples were incubated with the pancreatin enzyme,and the catalytic curve of reducing sugars (Fig.5)was fitted to a general first-order model (data not shown) to determine the apparent digestion rate coefficients and extent [5,17].The results were summarized in Table 2.
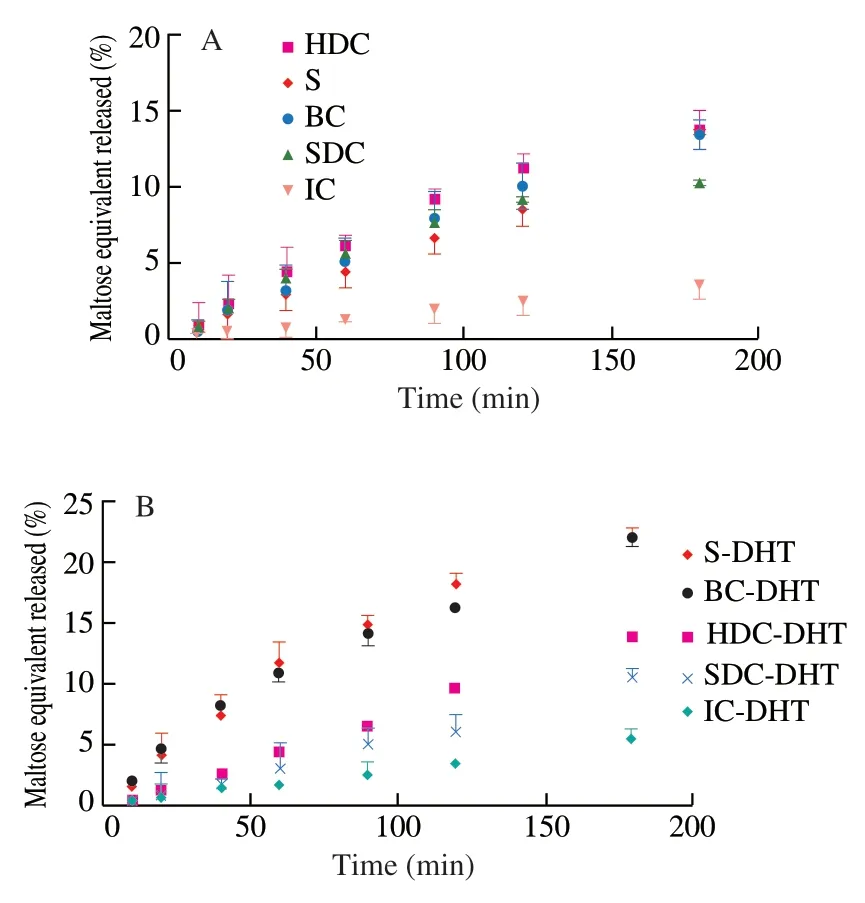
Fig.5 Digestion profiles of garbanzo bean cells and starches with different cell wall breakage before (A) and after (B) dry-heat-treatment.
The IC exhibited the lowest digestion kinetics (Table 2,0.000 2 min-1,3.6%) among all cell and starch samples.The increased starch digestion rate and extent of the cell samples with reduced cell wall integrity was observed compared with the IC,although the inhibitory activities of the soluble components in the cell samples were also enhanced (Fig.4A).Thus,the barrier functions of the cell wall would be the key factor for starch digestion in the cell samples.Although our previous study regarding the starch digestion in pulse cells processed with different methods exhibited that higher cell wall permeability would result higher digestion properties,the HDC,BC and pure starch samples in present study exhibited similar digestion rates and extents.This result could be attributed to higher damage degree of the cell wall structure,which greatly reduced the hindrance for digestion enzymes.
After subjection to DHT,the cell and starch samples all exhibited higher digestion rates than their native counterparts,which could be attributed to the lower inhibitor effects on enzymatic activity resulting from the soluble and insoluble components of the cell samples.It is worth noting that the increase in the digestion rate coefficients and extent of BC and pure starches after DHT (BC-DHT,S-DHT)compared with their native counterparts (BC,S) were significantly higher than that of other cell samples (IC-DHT,SDC-DHT and HDC-DHT).As Table 2 shows,the digestion extent increase(approximately 8%) of the BC-DHT samples was about 4 times greater than that of IC-DHT samples (approximately 2%),although the soluble components of the BC-DHT samples exhibited remarkably greater inhibition of enzymatic activity than their IC-DHT counterparts (Fig.4A).This result further supports the protective effects of the cellular structure on starch granules during the digestion process.Cells with a higher degree of cell wall integrity provide barriers for digestive enzymes to access the starch substrate,thus resulting in reduced digestion.The enzymatic susceptibility to a starch substrate entrapped in pulse cells would be the key factor affectingin vitrostarch digestion [35,36].In addition to the barrier effects from the cell wall,the greater retention in ΔHas well as the relative crystallinity of the starches inside the cells subjected to DHT(IC-DHT,SDC-DHT and HDC-DHT) could also contribute to the lower enzymatic susceptibility when compared with the BC and pure starch counterparts.
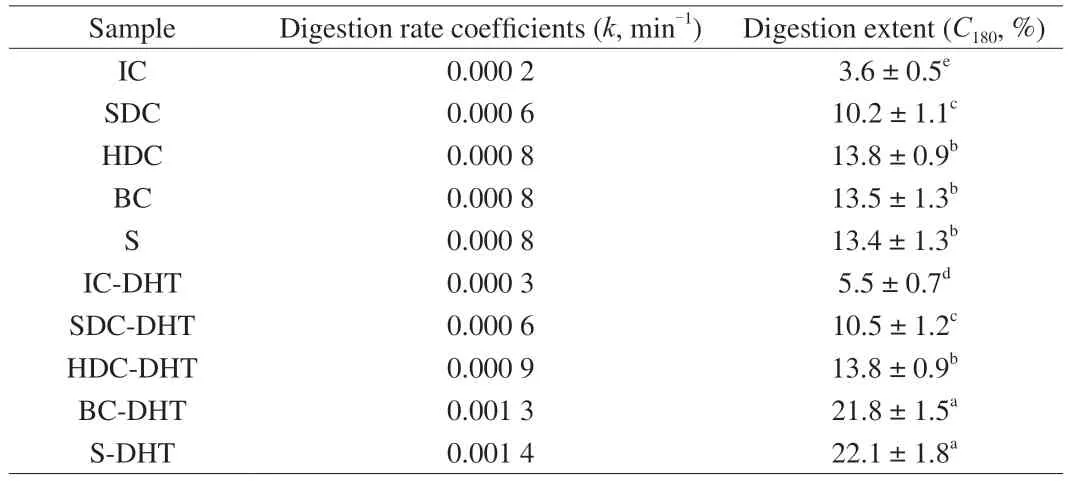
Table 2 Digestion rate coefficients (k) and reducing sugar released after 180 min (C180)digestion of garbanzo bean cells with different cell wall integrity before and after dry-heat-treatment1.
4.Conclusions
The structural andin vitrostarch digestion properties of garbanzo bean cells with variable degrees of cell wall integrity subjected to DHT were investigated.The morphological features suggested that starches in all cell samples do not exhibit remarkable changes after subjection to DHT.However,molecular rearrangement and crystallite disruption of the starch granules entrapped in the cell samples would concurrently occur upon after subjection to DHT,as analyzed by differential scanning calorimetry (DSC) and XRD.Although DHT decreased the inhibitory activity of both soluble and insoluble components on enzyme,the digestion rate and extent of the cell samples (SDC-DHT and HDC-DHT) did not exhibit significant differences when compared with their native counterparts (SDC,HDC).In contrast,the digestion rate and extent of the BC-DHT and S-DHT samples was increased dramatically when compared with native samples.Thus we concluded that the starch digestion of pulse cotyledon cells is mainly determined by the intactness of the cellular structure.This study revealed the role of the cell wall structure during physical modification to obtain starch with desirable nutritional properties,which will provide a fundamental basis for designing the next generation of whole food products and functional ingredients from pluses.
Declaration of competing interest
The authors declare no competing financial interest.
Acknowledgments
The authors thank the National Natural Science Foundation of China (31701546),the Fundamental Research Funds for the Central Universities of China (2019ZD40) and the 111 Project(B17018) for financial support.Bin Zhang thanks Pearl River Talent Recruitment Program of Guangdong Province (2017GC010229)and the Pearl River Nova Program of Guangzhou (201906010079).Ping Li thanks the National Natural Science Foundation of China(32001691),the special fund for scientific innovation strategyconstruction of high-level academy of agriculture science (R2019YJYB1001),the Application-oriented Projects of Guangdong Province(2017B020232002).
杂志排行

食品科学与人类健康(英文)的其它文章
- Colloidal nanoparticles prepared from zein and casein:interactions,characterizations and emerging food applications
- Biological factors controlling starch digestibility in human digestive system
- Preparation methods,biological activities,and potential applications of marine algae oligosaccharides: a review
- Development of hyaluronic acid-based edible film for alleviating dry mouth
- Mushroom β-glucan and polyphenol formulations as natural immunity boosters and balancers: nature of the application
- Preparation of multicore millimeter-sized spherical alginate capsules to specifically and sustainedly release fish oil