吊罗山石碌含笑群落木本植物种间联结性研究
2023-01-16魏亚情宋希强史佑海
魏亚情,宋希强,赵 莹,史佑海*
吊罗山石碌含笑群落木本植物种间联结性研究
魏亚情1,2,宋希强1,3,赵 莹1,3,史佑海1,3*
1. 海南大学海南省热带特色花木资源生物学重点实验室,海南海口 570228;2. 海南大学生态与环境学院,海南海口 570228;3. 海南大学林学院,海南海口 570228
石碌含笑()是海南特有树种和国家Ⅱ级濒危保护植物,野外仅见分布于吊罗山、鹦哥岭、五指山等海南中南部山区。吊罗山是石碌含笑最集中的自然分布区,开展吊罗山石碌含笑群落木本植物种间联结性研究,有助于了解该树种在群落中的地位与作用,为制定合理的就地保护措施提供参考。本研究基于2×2列联表,综合运用2检验、Pearson相关系数检验和Spearman秩相关系数检验对吊罗山石碌含笑与群落内其他伴生种的种间关系进行了探讨分析。结果表明:石碌含笑种群与灌木层的正关联比乔木层多;石碌含笑与群落内绝大多数植物的种间联结不显著,种间关系表现出总体比较松散的趋势,种群间相互独立的趋势明显,乔木层和灌木层总体上与石碌含笑正关联物种多于负关联物种数,表明群落的发展朝着有益于石碌含笑种群发育的趋势。研究结果对制定合理的就地保护措施和恢复计划具有参考价值。
石碌含笑;种间联接性;群落;吊罗山; 海南
种间联结性是不同物种之间在空间环境分布上的关联性[1],是植物种群间相互联系和相互影响的综合反映[2],是群落构建的结果。通过对物种间联结性的研究,能帮助我们对群落的正确认识,在群落功能分类等方面也有着举足轻重的意义[3-8]。
海南特有种石碌含笑()为国家Ⅱ级重点保护植物和优良的园林观赏树种,具有重要的保护价值和园林应用潜力。对石碌含笑的研究目前仅限于对其系统分类、生长特性、化合物提取、资源分布和遗传多样性等方面[9-15],尚未见有关石碌含笑群落种间联结性的研究报道。
1 材料与方法
1.1 研究区域自然概况
吊罗山(18°50N, 109°50E)地处海南东南部,位于东亚季风区,年均温在600 m高海拔处为20.8℃,一年中最热月平均温度和最冷月平均温度分别23.9℃和16.3℃,全年气温高于10℃以上的年积温达到7989℃。干季和湿季气候特征分化明显,年降雨量2566 mm,当年12月至次年1月为旱季,2—3月为过渡季,4—10月为雨季,2566 mm的年降雨量中,其中绝大部分来自雨季的降水[16]。300 m以下低海拔主要为砖红壤,300 m以上高海拔主要是山地黄壤[17]。
1.2 方法
1.2.1 样地设置与调查 选取石碌含笑具有代表性的地段设置20 m×30 m样地6个,采用相邻格子法将每个样地划分成6个10 m×10 m的样方,共36个样方。乔木记录所有物种的种名、树高、胸径、盖度、枝下高、冠幅和生长情况等。灌木选取每样方四角及中间共5个5 m×5 m小样方,灌木层记录植物的高度、盖度、株数、物候期等。同时记录样地的坡向、坡位、海拔等生境因子。对所调查的物种,选取乔木层、灌木层出现样方频度前20物种(表1,表2),计算分析种间的联结性[18]。
1.2.2 总体关联性检验 采用SCHLUTER的方差比率()检验关联的显著性[19]。
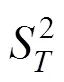
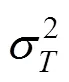
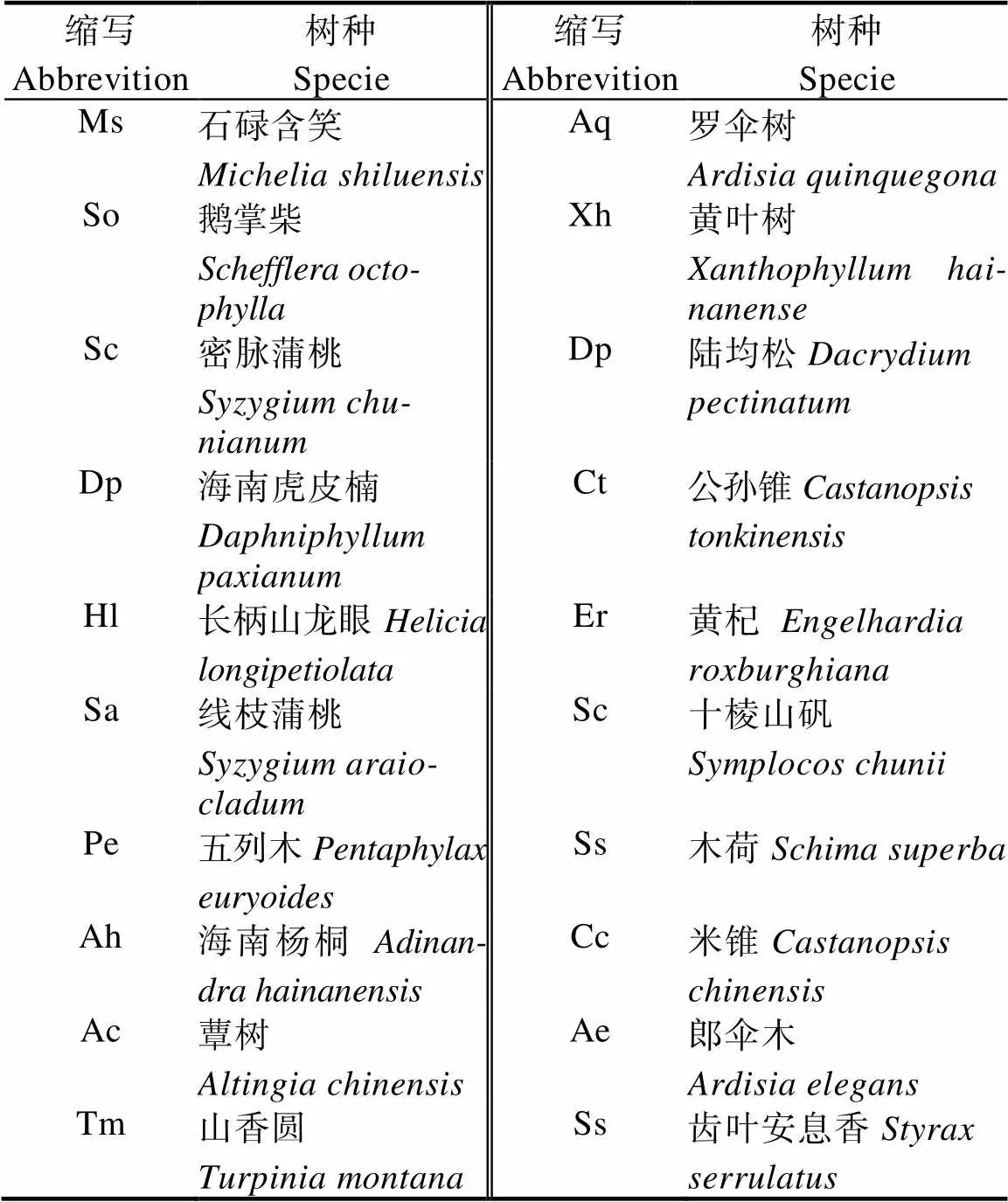
表1 乔木层主要树种种名及缩写
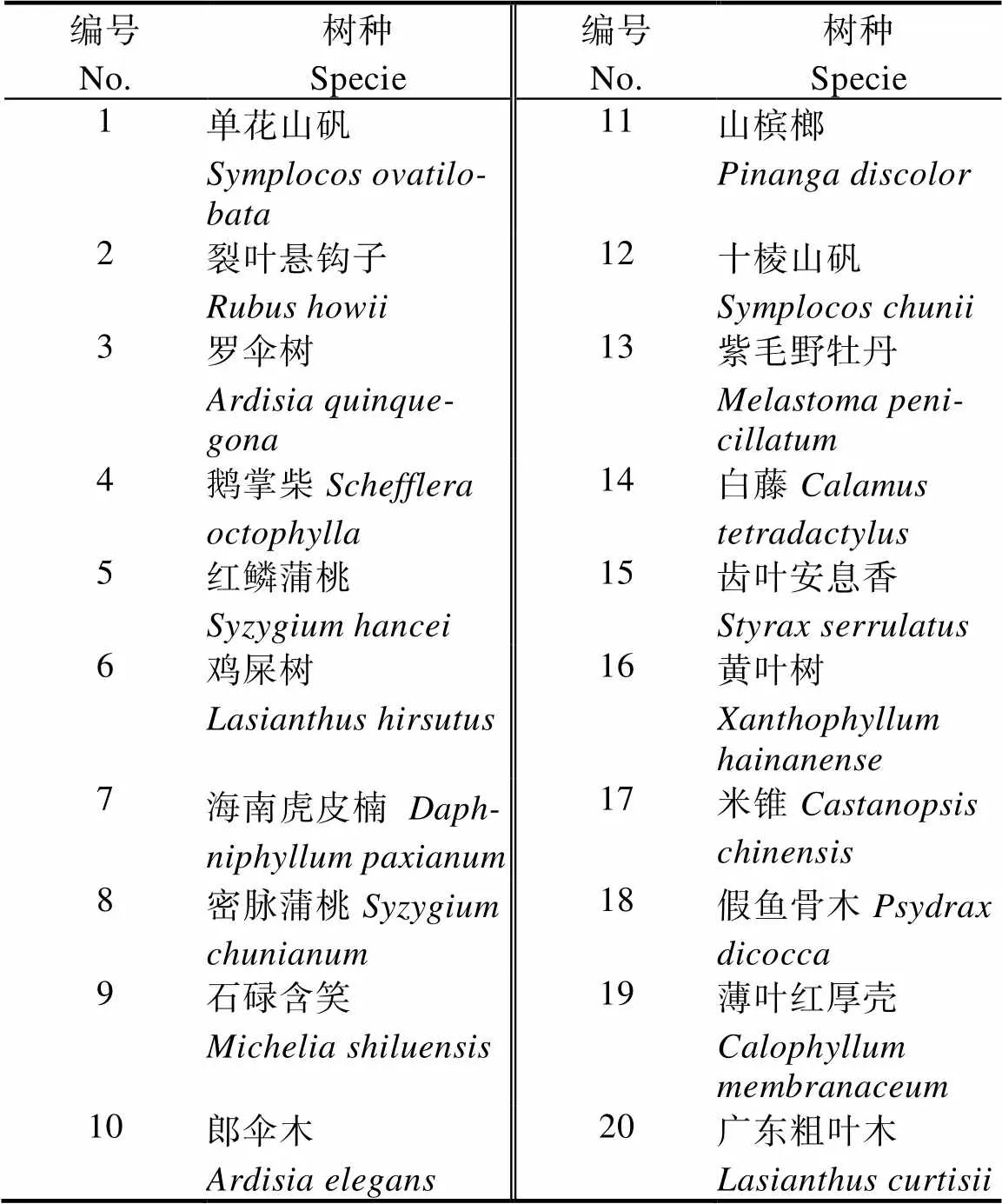
表2 灌木层主要树种种名及编号
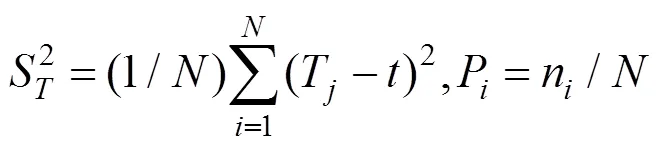
式中,:物种总数;:样方总数;n:物种出现的样方数;P:物种出现的频度;T:样方出现的物种总数;:全部样方种的平均数;S:所有样方物种数的方差;σ:为所有物种出现频度的方差。
在独立性假设条件下群落内植物种间的总体联结指数期望值为1,若=1,种间总体无关联;若>1,种间总体关联;若<1,物种总体呈负关联。为检验偏离1是否显著,引入新的统计量,=×。若<20.95N或>20.95N,则物种总体关联显著;若<20.95N<,则物种间总体关联不显著。
1.2.3 种对间关联性分析 用2检验来对种间关系进行定性研究,先将调查的原始数据的20个乔木转化成20×36的0,1二元数据矩阵,0,1分别表示物种在样方内的存在情况(0:不存在,1:存在)[20]。再将2个种出现与否的观测值填入2×2列联表(表3)[21]。

表3 2×2列联表
分别计算、、、值。其中为A、B两个种都出现的样方数,为种A出现而种B不出现的样方数,为种A不出现而种B出现的样方数,为2个种都不出现的样方数[2]。2值根据Yates[22]连续性校正公式计算:
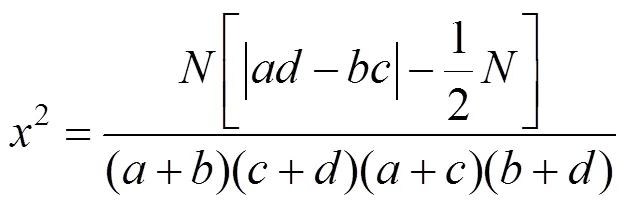
式中,表示总样方数。当2<3.841(>0.05)时,种对间相互独立,即种对间联结不显著;当3.841<2<6.635(0.01≤≤0.05)时,种对间联结性显著;当2>6.635(<0.01),种对间联结性极显著。
2检验只能定性地描述种间关联显著与否,忽略了关联不显著的种对间的关联性,且2检验无法判断联结强度的大小,无法清楚地反映种间关联性的差异性[23]。与之相比,Pearson相关系数检验和Spearman秩相关系数检验则是基于数量数据,能够清楚地反映物种间的线性关系,这2个相关系数检验可以对2检验进行有效地补充和完善。因此,在进行种间关联性2检验后,需进一步利用Pearson相关系数检验和Spearman秩相关系数检验测定其关联强度。
Pearson相关系数为:
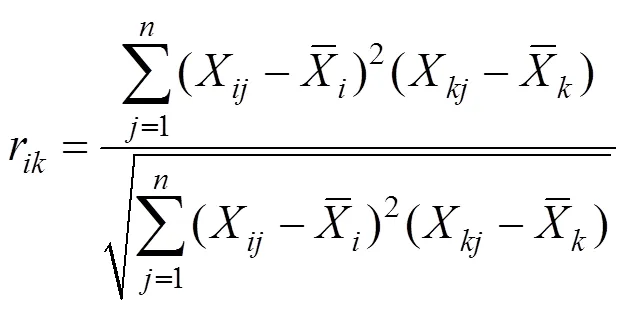
Spearman秩相关系数是首先需要将多度向量转化成秩化向量,然后将秩化向量代入公式计算得出。公式如下:
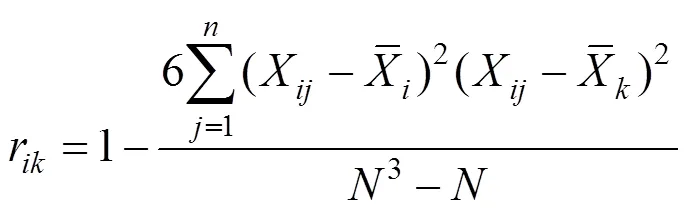
2 结果与分析
2.1 总体关联性
石碌含笑群落主要物种总体关联性见表4,乔木层和灌木层总体关联性的>1,>20.05N,即石碌含笑群落乔木层和灌木层主要种群在总体上是显著正关联。反映出吊罗山石碌含笑群落正处于较稳定的顶极阶段。
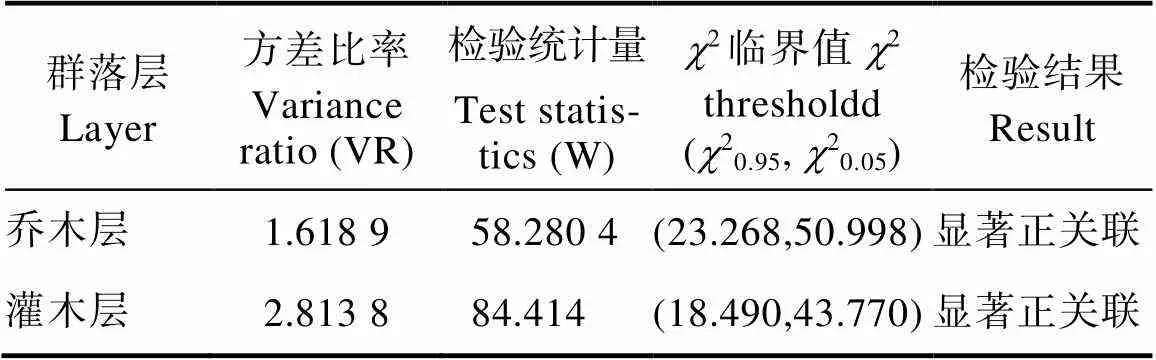
表4 石碌含笑群落乔木层和灌木层主要物种总体关联性
2.2 乔木层种间关联性
分别运用2检验、Pearson检验和Spearman秩相关检验法对石碌含笑群落乔木层种间关联性进行分析,结果见表5。
表5 石碌含笑群落乔木层主要树种种间关联性
2.2.1 乔木层2检验 乔木层2检验结果显示(图1),190对乔木种对中,线枝蒲桃与黄叶树、石碌含笑与密脉蒲桃、密脉蒲桃与线枝蒲桃、五列木与十棱山矾、海南杨桐与十棱山矾5对呈显著正关联。海南杨桐与蕈树、山香圆与齿叶安息香、鹅掌柴和陆均松3对呈显著负相关。无关联或不显著关联182对,占总种对数的95.9%,表明群落中乔木种群间关联不强,物种间的分布上呈现很强的独立性。石碌含笑与密脉蒲桃呈显著正关联,表明它们具有一致的生物学特征和生态适应性。石碌含笑与绝大多数伴生种联结显著的比例较低,连接性弱,石碌含笑群落相对独立。
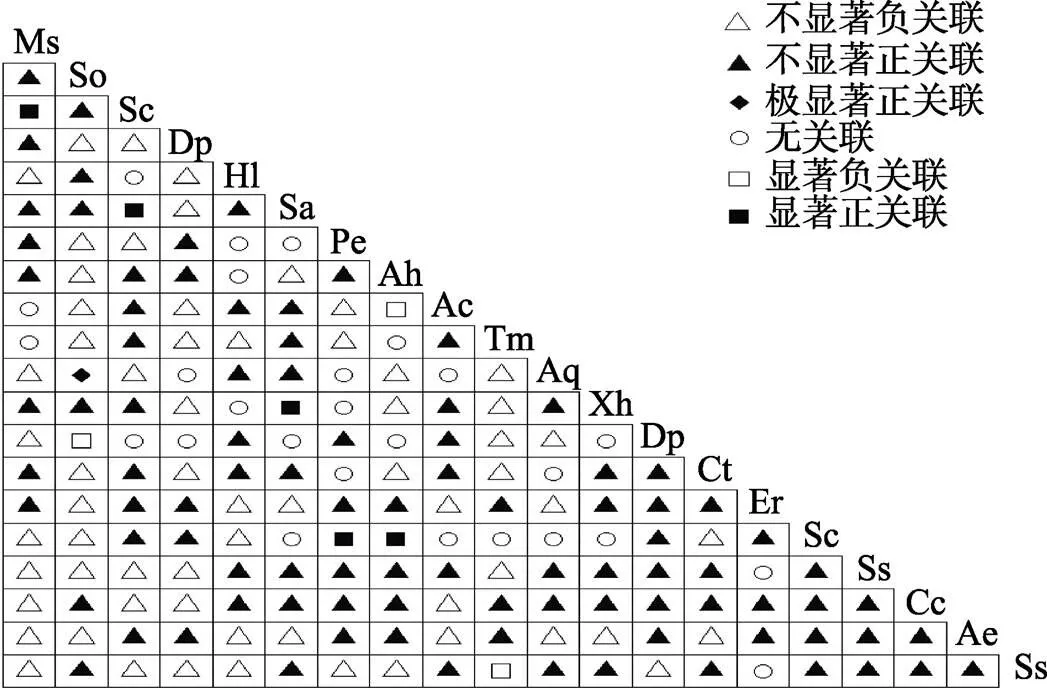
图1 石碌含笑乔木层种间关联χ2检验半矩阵图
2.2.2 乔木层Pearson检验 乔木层Pearson检验显示(图2),在所有190个种对中,极显著正关联有5对,分别是五列木与十棱山矾、海南杨桐与十棱山矾、线枝蒲桃与黄叶树、鹅掌柴与罗伞树、密脉蒲桃与线枝蒲桃,占总种对数的2.6%。显著正关联10对,即石碌含笑与密脉蒲桃、鹅掌柴与齿叶安息香、密脉蒲桃与黄杞、长柄山龙眼与陆均松、线枝蒲桃与蕈树、五列木与海南杨桐、罗伞树与齿叶安息香、黄叶树与齿叶安息香、黄叶树与米锥、黄杞与郎伞木,占总种对数的5.2%;显著负关联7对,即鹅掌柴与陆均松、密脉蒲桃与海南虎皮楠、山香圆与罗伞树、山香圆与齿叶安息香、公孙锥与郎伞木、五列木与山香圆等,占总种对数的3.6%。可见,种间关联性不强的种对占绝大多数,其中仅密脉蒲桃与石碌含笑呈显著正相关,结果与2检验一致。
2.2.3 乔木层Spearman秩相关检验 乔木层Spearman秩相关检验显示(图3),极显著正联结5对,占总数的2.6%;显著正关联10对,占总种对数的5.2%,其中石碌含笑仅与密脉蒲桃显著正关联。显著负关联7对,占总种对数的3.6%,结果与2检验、Pearson相关系数一致。
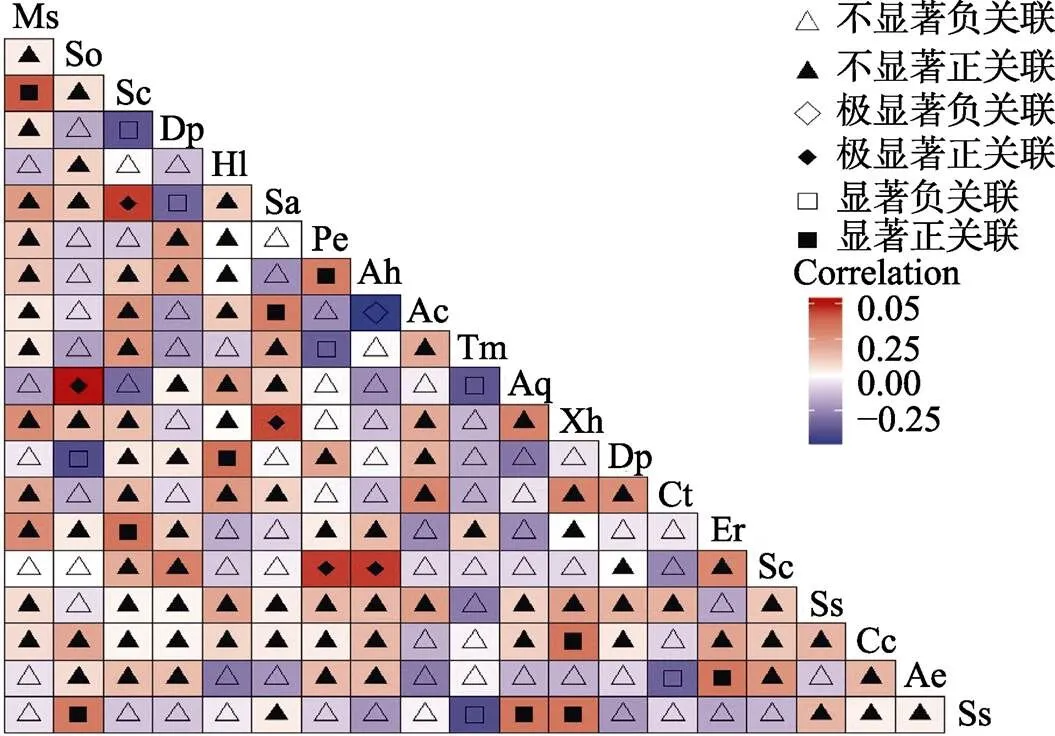
图2 乔木层Pearson相关系数检验
2.3 灌木层种间关联性
分别运用2检验、Pearson检验和Spearman秩相关检验法对石碌含笑群落灌木层种间关联性进行分析,结果如表6所示。
2.3.1 灌木层2检验 灌木层种对间2检验显示(图4),190个种对中,极显著正关联1个,即罗伞树与红鳞蒲桃,占总种对数的0.6%。罗伞树与密脉蒲桃、罗伞树与黄叶树、鹅掌柴与鸡屎树、鸡屎树与黄叶树、红鳞蒲桃与密脉蒲桃、红鳞蒲桃与白藤、红鳞蒲桃与黄叶树、鸡屎树与紫毛野牡丹、密脉蒲桃与薄叶红厚壳、石碌含笑与薄叶红厚壳、山槟榔与白藤、齿叶安息香与黄叶树、齿叶安息香与米锥13个种对呈显著正关联,占总种对数的6.8%。裂叶悬钩子与薄叶红厚壳、海南虎皮楠与米锥、山槟榔与十棱山矾、十棱山矾与白藤、十棱山矾与齿叶安息香5对呈显著负相关,占总种对数的2.6%。无关联或不显著关联171对,占总种对数的90%,表明群落种灌木种群间关联较弱,个物种间呈独立分布。石碌含笑仅与薄叶红厚壳呈显著正关联,表明它们具有一致的生物学特征和生态适应性。无极显著关联,石碌含笑与绝大多数伴生种联结显著的比例较低,连接性弱,说明石碌含笑在群落中相对独立。
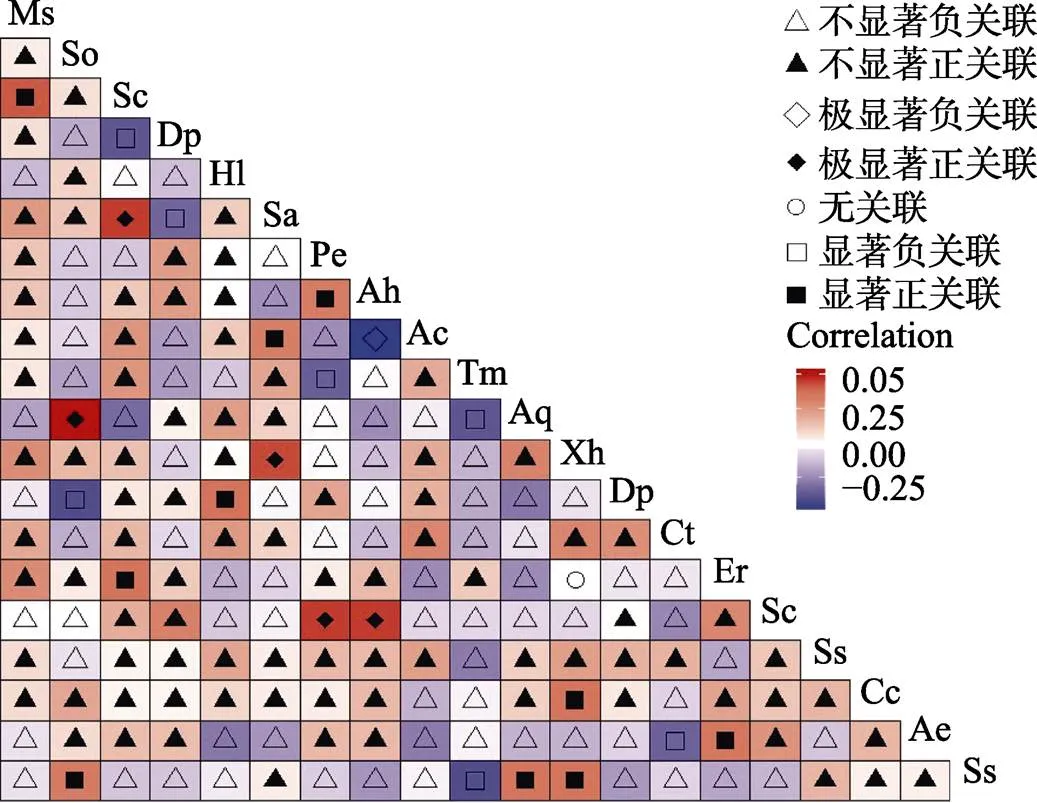
图3 乔木层Spearman秩相关检验
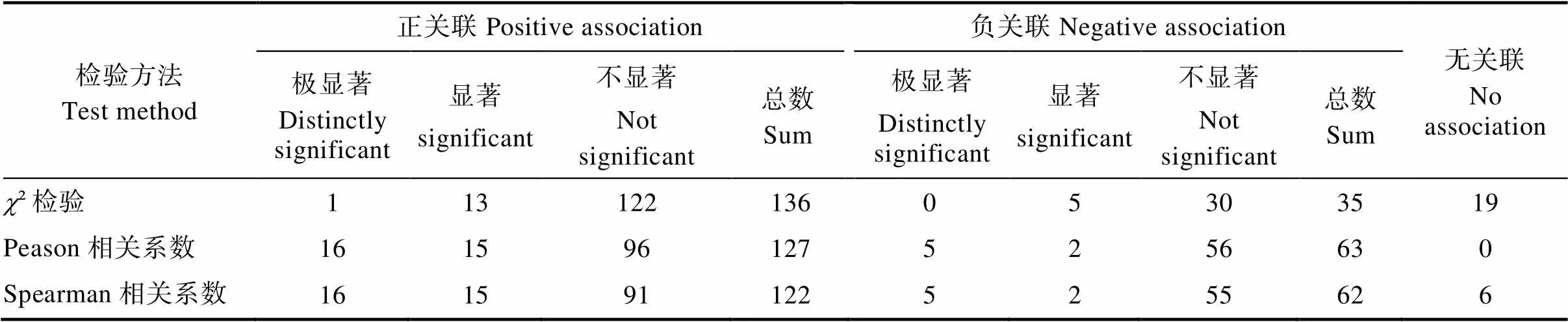
表6 石碌含笑群落灌木层主要树种种间关联性
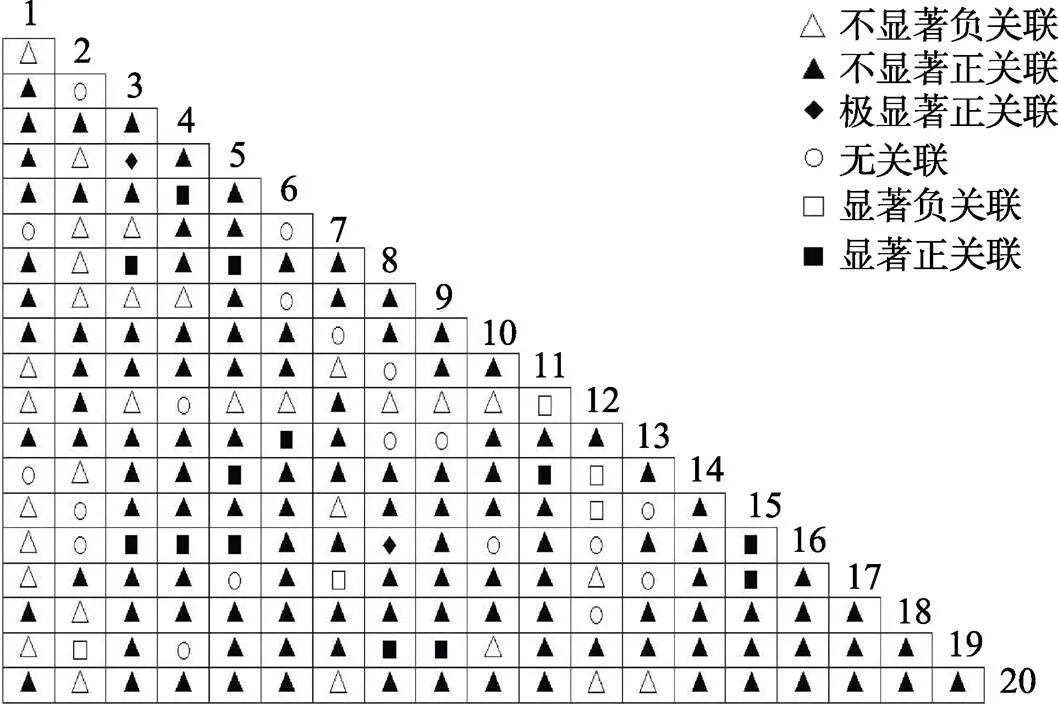
图4 灌木层种间关联χ2检验半矩阵图
2.3.2 灌木层Pearson检验 灌木层Pearson相关系数检验显示(图5),16个种对呈极显著正关联,占总种对数的8.4%;5个种对呈极显著负关联,占总种对数的2.6%;15个种对呈显著正关联,占总种对数的7.9%;2种对呈显著负关联,占总种对数的1.1%;无关联或不显著关联152对,占总种对数80%。种间关联性不强的种对占大多数,其中石碌含笑仅与鱼骨木呈显著正相关,与薄叶红厚壳呈极显著正关联,结果与2检验相似。
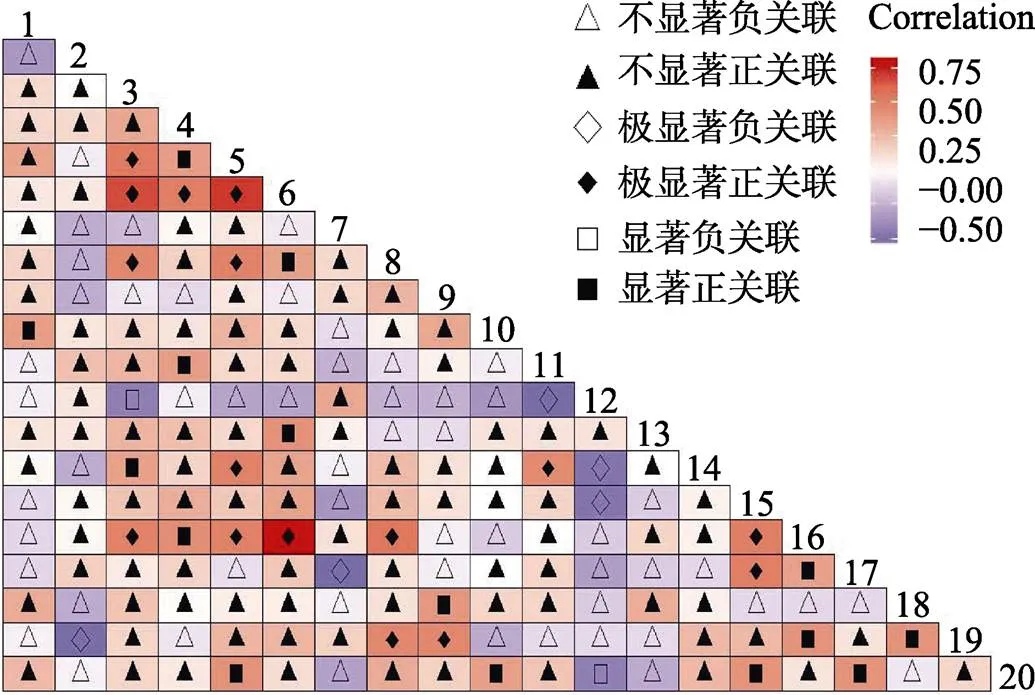
图5 灌木层Pearson相关系数检验
2.3.3 灌木层Spearman秩相关系数检验 灌木层Spearman秩相关系数检验显示(图6),16个种对间呈极显著正关联,占总种对数的8.4%;5个种对呈极显著负关联,占总种对数的2.6%;15个种对呈显著正关联,占总种对数的7.9%;2种对呈显著负关联,占总种对数的1.1%;无关联或不显著关联152对,占总种对数80%。种间关联性不强的种对占大多数,其中石碌含笑仅与鱼骨木为显著正相关,与薄叶红厚壳为极显著正关联,结果与2检验及Pearson检验结果相一致。
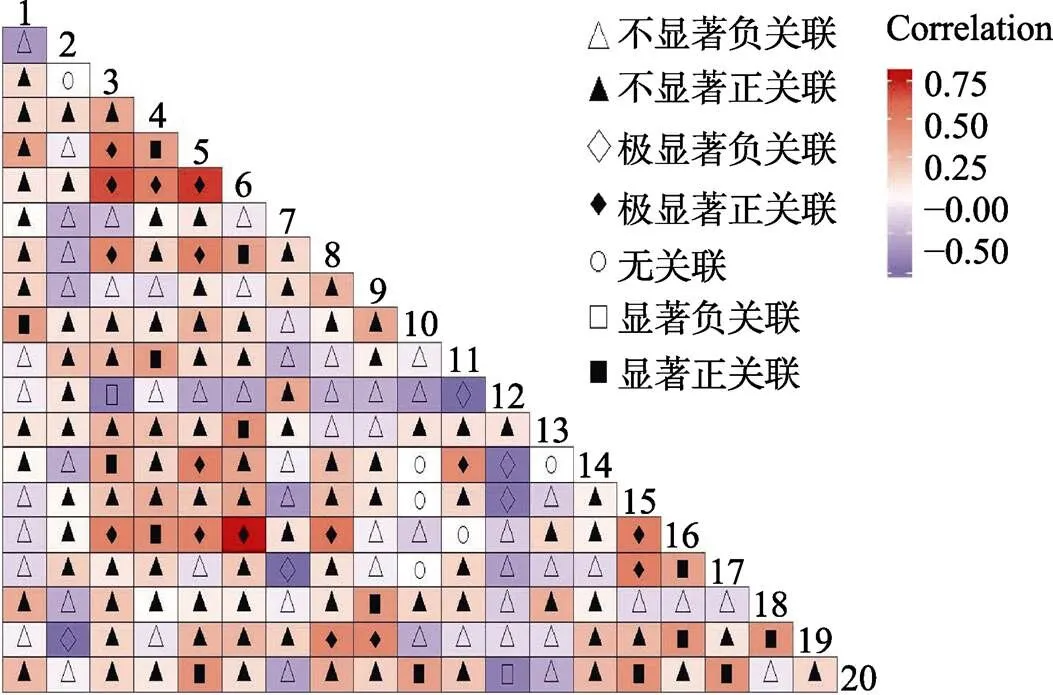
图6 灌木层Spearman秩相关系数
3 讨论
石碌含笑群落种间联结结果表明,乔木层和灌木层种间总体关联性呈显著正关联。3种相关系数结果表明,乔木层、灌木层190个种对中正负联结种对数比值均大于1,正联结种对数大于负联结,反映出吊罗山石碌含笑群落正处于较稳定的顶级阶段[24]。乔木层和灌木层绝大部分种对间表现出关联性呈不显著,种对间关联性较弱,联结松散,趋于独立分布。现阶段石碌含笑大部分群落结构,已处于和当地环境条件相适应的相对稳定状态,各种群之间都能占据与自己相对契合的生态位,物种对相互之间的依赖性以及竞争性极大地弱化了[25]。这一结果与邓贤兰等[26]得出的结论类似。在检验结果上,3种检验方法表现出一致性,总体上石碌含笑与群落内绝大多数植物的种间联结不显著,种间关系表现出总体比较松散的趋势,种群相互间独立的趋势明显,随机性程度很大,群落中各物种与石碌含笑间联结松散。乔木层和灌木层总体上与石碌含笑正关联物种多于负关联物种数。综上所述,石碌含笑群落处于较稳定的顶级阶段,种对间多数未达到显著联结性,种对间独立性强,联结松散,群落的发展朝着有益于石碌含笑的趋势。
若物种间有相似的生存生境,或者相似的利用资源的方式,则植物种对往往通过物种间的正联结体现[27-28]。石碌含笑群落的乔木层15对种对正联结程度较强,达到显著水平,其中5对种对联结关系达到极显著的水平;灌木层所有种对中,较强正联结31对,且都达到了显著水平,其中达到极显著关联的种对有16对。显著正关联表明两物种对间相互有利,或是在应对环境的变化时有相似的响应策略,种群间能够共同分享资源,二者生态位产生重叠而呈现显著正联结,乔木层石碌含笑与密脉蒲桃呈显著正关联,石碌含笑为喜光树种,密脉蒲桃耐荫蔽,石碌含笑为密脉蒲桃提供了荫蔽的同时,避免了二者对光照竞争,分别占据群落中不同空间,避免了种间的激励竞争。类似的种对还有灌木层石碌含笑与薄叶红厚壳呈极显著正关联,与鱼骨木呈显著正关联。植物种对的负联结体现了物种间的排斥性,这是长期适应不同微环境,利用不同资源空间的结果,也是生态位分离的反映[29]。3种检验结果中,均未出现与石碌含笑显著负关联的物种,但存在不显著负相关物种,种间联结性结果侧面反映了物种间复杂的关系,以及植物对环境等因子的适应程度。石碌含笑与密脉蒲桃、薄叶红厚壳、鱼骨木呈显著正关联,是有益的伴生种。在进行石碌含笑种群的同时,也应进行适度的人为干扰对与密脉蒲桃、薄叶红厚壳、鱼骨木加以保护,同时也注意监控负联结物种等措施来保护石碌含笑的生存环境[27]。
2检验、Spearman秩相关分析、Pearson相关系数检验3种方法比较反应较为相似的结果,但也存在一定的差异。2检验是以二元数据作为联结性判断的标准,且+检验只是对种间联结性作出了一个概括性的结论,这样会导致种间相关的多度信息的丧失,不能区分联结强度的大小。但其分析较简单,指标明确,能比较准确客观地反映出种间联结性。Pearson分析需要呈正态分布的基础上进行,而Spearman秩相关分析与物种如何分布不相关,所以为补充相关系数检验的不足,需引入Spearman秩相关分析做补充[28]。在进行种间关系研究时,应先进行2检验,在其检验结果的基础上,再结合另外2种相关系数(Pearson和Spearman)等方法进行检验,共同分析群落中各种对的种间关系,从而相对全面、准确地反映植物群落的种间联结性[30]。
吊罗山自然保护区是石碌含笑种群分布保存最完好的地区,本研究显示石碌含笑群落中正关联数大于负关联,说明吊罗山分布区群落目前正朝着有利于石碌含笑种群的方向发展,因而,应该对吊罗山石碌含笑种群加强就地保护,并建立永久样地,纵向比较群落的种间关系变化[31]。
[1] 王伯荪. 植物种群学[M]. 广州: 中山大学出版社, 1989: 115-120.
WANG B S. Phytopopulology[M]. Guangzhou: Sun Yat-sen University Press, 1989: 115-120 (in Chinese)
[2] 张忠华, 梁士楚, 胡 刚. 桂林喀斯特石山阴香群落主要种群的种间关系[J]. 山地学报, 2007, 25(4): 475-482.
ZHANG Z H, LIANG S C, HU G. Interspecific relationships among main species ofcommunity on Karst hills of Guilin[J]. Journal of Mountain Science, 2007, 25(4): 475-482. (in Chinese)
[3] 王伯荪, 彭少麟. 鼎湖山森林优势种群数量动态[J]. 生态学报, 1987, 7(3): 214-221.
WANG B S, PENG S L. Quantitative dynamics of the dominant population in the forest communities of Dinghushan[J]. Acta Ecologica Sinica, 1987, 7(3): 214-221. (in Chinese)
[4] 李先琨, 黄玉清, 苏宗明. 南方红豆杉群落主要树木种群间联结关系初步研究[J]. 生态学杂志, 1999, 18(3): 10-14.
LI X K, HUANG Y Q, SU Z M. Preliminary studies on interconnections among the main tree populations ofvar.community[J]. Chinese Journal of Ecology, 1999, 18(3): 10-14. (in Chinese)
[5] 杨兆静, 张钦弟, 李 豪, 白玉宏, 毕润成. 山西五鹿山自然保护区暴马丁香群落木本植物种间联结性分析[J]. 植物科学学报, 2013, 31(1): 42-48.
YANG Z J, ZHANG Q D, LI H, BAI Y H, BI R C. Inter-specific correlations among dominant populations of woodycommunities in Wulu Mountain Nature Reserve, Shanxi province of China[J]. Plant Science Journal, 2013, 31(1): 42-48. (in Chinese)
[6] 张金屯, 焦 蓉. 关帝山神尾沟森林群落木本植物种间联结性与相关性研究[J]. 植物研究, 2003, 23(4): 458-463.
ZHANG J T, JIAO R. Interspecific association between woody plants in Shenweigou of Guandi Mountains, Shanxi province[J]. Bulletin of Botanical Research, 2003, 23(4): 458-463. (in Chinese)
[7] 潘 高, 张合平, 潘 登. 中亚热带南酸枣林优势种群的种间联结性[J]. 生态学杂志, 2017, 36(4): 892-901.
PAN G, ZHANG H P, PAN D. Interspecific associations of dominant plant populations in a mid-subtropicalforest[J]. Chinese Journal of Ecology, 2017, 36(4): 892-901. (in Chinese)
[8] 叶权平, 张文辉, 于世川, 薛文艳. 桥山林区麻栎群落主要乔木种群的种间联结性[J]. 生态学报, 2018, 38(9): 3165-3174.
YE Q P, ZHANG W H, YU S C, XUE W Y. Interspecific association of the main tree populations of thecommunity in the Qiaoshan forest area[J]. Acta Ecologica Sinica, 2018, 38(9): 3165-3174. (in Chinese)
[9] 魏亚情, 洪 峰, 袁浪兴, 孔焱焱, 史佑海. 海南特有濒危植物石碌含笑的分布现状与种群年龄结构特征[J]. 热带作物学报, 2017, 38(12): 2280-2284.
WEI Y Q, HONG F, YUAN L X, KONG Y Y, SHI Y H. Population distribution and age structure characteristics ofan endangered and endemic species in Hainan Island[J]. Chinese Journal of Tropical Crops, 2017, 38(12): 2280-2284. (in Chinese)
[10] 刘玉壶, 周仁章, 曾庆文. 木兰科植物及其珍稀濒危种类的迁地保护[J]. 热带亚热带植物学报, 1997, 5(2): 1-12.
LIU Y H, ZHOU R Z, ZENG Q W.conservation of Magnoliaceae including its rare and endangered species[J]. Journal of Tropical and Subtropical Botany, 1997, 5(2): 1-12. (in Chinese)
[11] 刘仁林. 石碌含笑(Chun et Y. F. Wu)[J]. 江西林业科技, 2005, 33(5): 64.
LIU R L.Chun et Y. F. Wu[J]. Jiangxi Forestry Science Technology, 2005, 33(5): 64. (in Chinese)
[12] 包淑云, 周守标, 喻永红. 含笑属叶片的比较解剖学研究[J]. 广西植物, 2002, 22(2): 140-144, 193.
BAO S Y, ZHOU S B, YU Y H. Comparative anatomy of the leaves in[J]. Guihaia, 2002, 22(2): 140-144, 193. (in Chinese)
[13] 易润华, 陈玉娟, 韩静仪, 胡 倩, 李慧琪, 吴海燕. 石碌含笑枯梢病菌的鉴定及生物学特性[J]. 林业科学, 2018, 54(9): 80-88.
YI R H, CHEN Y J, HAN J Y, HU Q, LI H Q, WU H Y. Identification and biological characteristics ofcausing dieback disease on[J]. Scientia Silvae Sinicae, 2018, 54(9): 80-88. (in Chinese)
[14] XIONG J, WANG L J, QIAN J C, WANG P P, WANG X J, MA G L, ZENG H, LI J, HU J F. Structurally diverse sesquiterpenoids from the endangered ornamental plant[J]. Journal of Natural Products, 2018, 81(10): 2195-2204.
[15] DENG Y W, LIU T T, XIE Y Q, WEI Y Q, XIE Z C, SHI Y H, DENG X M. High genetic diversity and low differentiation in, an endangered magnolia species in south China[J]. Forests, 2020, 11(4): 469.
[16] 何园球, 王明珠, 赵其国. 我国热带亚热带森林土壤的水热动态[J]. 土壤, 1988, 20(5): 225-231.
HE Y Q, WANG M Z, ZHAO Q G. Hydrothermal dynamics of tropical and subtropical forest soils in China[J]. Soils, 1988, 20(5): 225-231. (in Chinese)
[17] 曾昭璇, 曾宪中. 海南岛自然地理[M]. 北京: 科学出版社, 1989.
ZENG Z X, ZENG X Z. Physical geography of the Hainan Island [M]. Beijing: Science Press, 1989. (in Chinese)
[18] 林长松, 李玉英, 朱万斌, 程 序. 珍稀保护植物十齿花群落灌木种间联结性分析[J]. 中国农学通报, 2008, 24(3): 106-111.
LIN C S, LI Y Y, ZHU W B, CHENG X. Analysis on interspecific association among shrub species of rare and precious plantcommunities in Yushe National Forest Park, Guizhou[J]. Chinese Agricultural Science Bulletin, 2008, 24(3): 106-111. (in Chinese)
[19] SCHLUTER D. A variance test for detecting species associations, with some example applications[J]. Ecology, 1984, 65(3): 998-1005.
[20] DICE L R. Measures of the amount of ecologic association between species[J]. Ecology, 1945, 26: 297-302.
[21] 考克斯. 普通生态学实验手册[M]. 蒋有绪, 译. 北京: 科学出版社, 1979.
COX G W. Laboratory manual of general ecology[M]. JIANG Y X, trans. Beijing: Science Press, 1979. (in Chinese)
[22] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2004.
ZHANG J T. Quantitative ecology[M]. Beijing: Science Press, 2004. (in Chinese)
[23] 简敏菲, 刘 琪, 朱 笃, 游 海. 九连山常绿阔叶林乔木优势种群的种间关联性分析[J]. 植物生态学报, 2009, 33(4): 672-680.
JIAN M F, LIU Q, ZHU D, YOU H. Inter-specific correlations among dominant population of tree layer species in evergreen broad-leaved forest in Jiulianshan Mountain of subtropical China[J]. Chinese Journal of Plant Ecology, 2009, 33(4): 672-680. (in Chinese)
[24] 杜道林, 刘玉成, 李 睿. 缙云山亚热带栲树林优势种群间联结性研究[J]. 植物生态学报, 1995, 19(2): 149-157.
DU D L, LIU Y C, LI R. Studies on the interspecific association of dominant species in a suntropicalforest of Jinyun Mountain, China[J]. Chinese Journal of Plant Ecology, 1995, 19(2): 149-157. (in Chinese)
[25] 刘润红, 姜 勇, 常 斌, 李娇凤, 荣春艳, 梁士楚, 杨瑞岸, 刘星童, 曾惠帆, 苏秀丽, 袁海莹, 傅桂焕, 吴燕慧. 漓江河岸带枫杨群落主要木本植物种间联结与相关分析[J]. 生态学报, 2018, 38(19): 6881-6893.
LIU R H, JIANG Y, CHANG B, LI J F, RONG C Y, LIANG S C, YANG R A, LIU X T, ZENG H F, SU X L, YUAN H Y, FU G H, WU Y H. Interspecific associations and correlations among the main woody plants in acommunity in a riparian zone of Lijiang River, Guilin, Southwest China[J]. Acta Ecologica Sinica, 2018, 38(19): 6881-6893. (in Chinese)
[26] 邓贤兰, 刘玉成, 吴 杨. 井冈山自然保护区栲属群落优势种群的种间联结关系研究[J]. 植物生态学报, 2003, 27(4): 531-536.
DENG X L, LIU Y C, WU Y. Interconnection among dominant plant populations of castanopsis comimuntty in Jinggang Mountain Nature Reserve[J]. Chinese Journal of Plant Ecology, 2003, 27(4): 531-536. (in Chinese)
[27] 黄祥童, 王绍先, 黄炳军, 尹 航, 崔凯峰, 赵 伟, 范宇光, 顾德峰. 珍稀植物对开蕨与其伴生物种的联结性及群落稳定性[J]. 生态学报, 2015, 35(1): 80-90.
HUANG X T, WANG S X, HUANG B J, YIN H, CUI K F, ZHAO W, FAN Y G, GU D F. Analyses of community stability and inter-specific associations between the rare plantand its associated species[J]. Acta Ecologica Sinica, 2015, 35(1): 80-90. (in Chinese)
[28] 徐满厚, 刘 敏, 翟大彤, 刘 彤. 植物种间联结研究内容与方法评述[J]. 生态学报, 2016, 36(24): 8224-8233.
XU M H, LIU M, ZHAI D T, LIU T. A review of contents and methods used to analyze various aspects of plant interspecific associations[J]. Acta Ecologica Sinica, 2016, 36(24): 8224-8233. (in Chinese)
[29] 孙中伟,赵士洞. 长白山北坡椴树阔叶红松林群落木本植物种间联结性与相关性研究[J]. 应用生态学报, 1996, 7(1): 1-5.
SUN Z W, ZHAO S D. Interspecific association and correlation of lime-broad leaved Korean pine forest on the northern slope of Changbai Mountain[J]. Chinese Journal of Applied Ecology, 1996, 7(1): 1-5. (in Chinese)
[30] 张 峰, 上官铁梁. 山西翅果油树群落种间关系的数量分析[J]. 植物生态学报, 2000, 24(3): 351-355.
ZHANG F, SHANGGUAN T L. Numerical analysis of interspecific relationships in ancommunity in Shanxi[J]. Chinese Journal of Plant Ecology, 2000, 24(3): 351-355. (in Chinese)
[31] 胡理乐, 江明喜, 党海山, 黄汉东. 从种间联结分析濒危植物毛柄小勾儿茶在群落中的地位[J]. 植物生态学报, 2005, 29(2): 258-265.
HU L L, JIANG M X, DANG H S, HUANG H D. Community studies on the status of the endangered plant,var., using interspecific association analysis[J]. Chinese Journal of Plant Ecology, 2005, 29(2): 258-265. (in Chinese)
Interspecific Association of Woody Plants inCommunity, Diaoluoshan Nature Reserve, Hainan Province
WEI Yaqing1,2, SONG Xiqiang1,3, ZHAO Ying1,3, SHI Youhai1,3*
1. Hainan Key Laboratory for Biology of Tropical Ornamental Plant Germplasm, Hainan University, Haikou, Hainan 570228, China; 2. College of Ecology and Environment, Hainan University, Haikou, Hainan 570228, China; 3. College of Foresty, Hainan University, Haikou, Hainan 570228, China
is one of the endemic plants and national class II endangered protected species in Hainan Island. It has a sporadic distribution in central and southern mountains of Hainan Island, including Diaoluoshan Mountain, Yinggeling Mountainand, Wuzhi Mountain, etc.Diaoluoshan Nature Reserve is the most concentrateddistribution area ofStudy on the interspecific association of woody plants incommunity in Diaoluoshan Nature Reserve and analysis of its status in the community is of great significance to in-situ conservation of the species.On the basis of a 2×2 contingency table,2-test, Pearson correlation coefficient test and Spearman rank correlation coefficient test were used to analyze the interspecific relationship betweenand other plants in the community.There were more positive correlations in the shrub layer than in the tree layer. There was no significant interspecific association betweenand most other plants in the community, the interspecific relationship showed a relatively loose trend in general, and the trend of mutual independence among populations was obvious.The tree layer and shrub layer were generally more positively associated with, indicating that the development of the community was beneficial to the development of thepopulation.
; interspecific association; community; Diaoluoshan Nature Reserve;Hainan province
Q948.114
A
10.3969/j.issn.1000-2561.2022.12.023
2022-03-28;
2022-05-07
海南省自然科学基金项目(No. 314069);海南省重点科技计划项目(No. ZDYF2020099)。
魏亚情(1991—),男,博士研究生,研究方向:植物资源保护与利用。*通信作者(Corresponding author):史佑海(SHI Youhai),E-mail:shiyouhai@163.com。
