大厂茶优势株系芽叶性状与生化成分的鉴定与评价
2023-01-16李彩云宋勤飞牛素贞赵支飞何应琴马安权
李彩云,宋勤飞,牛素贞*,赵支飞,何应琴,李 芳,马安权
大厂茶优势株系芽叶性状与生化成分的鉴定与评价
李彩云1,宋勤飞1,牛素贞1*,赵支飞1,何应琴1,李 芳1,马安权2
1. 贵州大学茶学院,贵州贵阳 550025;2. 普安县泰安白龙古茶专业合作社,贵州普安 561500
为科学合理选育出优质大厂茶无性系新品种(系),本研究以7个大厂茶优势株系为研究材料,以福鼎大白茶为对照,对其芽叶性状、主要生化成分、儿茶素组分及氨基酸组分进行综合分析和系统评价。结果表明,7个大厂茶优势株系芽叶性状呈中叶或大叶种,芽叶黄绿,无茸毛或少毛,叶椭圆或长椭圆形,叶质硬脆,叶齿浅且钝,叶面微隆,叶缘微波,叶厚有光泽等特征。生化成分分析表明,7个大厂茶优势株系水浸出物含量、游离氨基酸含量和ECG含量较高,属优良茶树种质资源。其水浸出物含量、咖啡碱含量、茶多酚含量、游离氨基酸含量、粗纤维含量、维生素C含量、蛋白质含量、水溶性碳水化合物含量变化范围分别为47.02%~48.70%、2.91%~3.42%、19.03%~21.07%、4.07%~4.84%、7.43%~8.38%、208.00~332.00 mg/100 g、33.82~35.07 mg/100 g、3.23%~3.85%,酚氨比变化范围为3.98~5.04。主要儿茶素组分含量大小顺序为C>ECG>EC>EGCG>EGC,C和ECG含量较高,均显著高于CK,其变化范围分别为5.25%~9.02%和6.11%~7.64%,而EGCG和EGC含量较低,显著低于CK,其变化范围分别为0.08%~1.44%和0.05%~0.31%。7个大厂茶优势株系共有19种氨基酸被检出,氨基酸各组分之间的种类和含量都存在一定的差异性,含量较高的5个氨基酸组分依次是茶氨酸>谷氨酸>精氨酸>天冬氨酸>丝氨酸,占氨基酸总量的82.14%以上。各芽叶性状与生化成分之间有一定相关性。基于16个生化成分的主成分分析显示,16个生化成分被简化为5个因子,累计贡献率达95.68%,包含16个生化成分的大部分信息。综合评价显示,7个大厂茶优势株系中排名前三的依次是L17、L22和L06,为大厂茶新品种(系)的选育奠定基础。本研究还鉴定出L06、L17和L203个具有适制绿茶潜在特性的大厂茶优势株系,L12、L13、L21和L224个具有红绿茶兼制潜在特性的大厂茶优势株系。
大厂茶;芽叶性状;生化成分;鉴定
贵州因其立体高原气候特征、茶树异花授粉特性以及人们科学的自然资源保护意识而形成和保留了大量的野生茶树资源,这为研究茶树起源、演化提供了宝贵资源[1-2]。大厂茶(F. S. Zhang)为山茶科(Theaceae)山茶属()茶组(.)植物[3],是滇黔桂地区特有珍稀茶树种质资源,在贵州西南部及南部地区均广泛分布[4],主要生长于低纬度、高海拔、温湿度较高的原始森林内[5]。其在植物学分类上大多属于乔木或小乔木型,叶革质、叶面隆起富光泽、叶形长圆(椭圆或长椭圆形)、子房五室、无毫的野生茶系[6]。大厂茶在系统进化上处于原始位置,是茶树起源中心的典型种,表型及生化成分分析发现其遗传多样性丰富且蕴含特异的茶叶功能生化成分,在抗逆、产量和生化相关性状上具有很好的优势,可用于新品种选育或者为茶树遗传改良提供优异基因来源[7-10]。近年来,随着人们逐渐重视对珍稀野生茶树种质资源(包括大厂茶、秃房茶等)的保护和利用,政府也不断完善相关法律法规,为大厂茶种质资源的收集、保护、鉴定评价和开发利用奠定了基础。目前,学者们对大厂茶种质资源的分布、生化多样性分析和优良单株鉴选已开展大量研究[11-12],但对其野生驯化和优势株系选育还较少涉及,迄今为止尚未鉴选出大厂茶新品种,在一定程度上制约了当地茶产业发展。
因此,本课题组前期于2016—2017年对分布在贵州黔西南州大厂茶群体种开展了调查筛选,并构建了普安古茶树核心种质,从中选育出22个优良古树单株,通过无性系扦插扩繁培育建成大厂茶株系试验园。于2019—2021年连续3 a对22个优良古树单株的植物学特性、物候期、产量构成因子及主要生化成分的观测,进一步选育出产量高、品质优、物候期较早的7个大厂茶优势株系。本研究以7个大厂茶优势株系为材料,采用传统鉴定技术和高效液相色谱技术相结合的手段,开展其芽叶性状及生化成分的鉴定和系统评价,以期为大厂茶新品种(系)的选育提供理论依据,同时为优化贵州茶产业品种资源配置、产业提质增效及可持续发展奠定基础。
1 材料与方法
1.1 材料
试验材料来源于种植在贵州省普安县内的7个四年生大厂茶优势株系,分别为L06、L12、L13、L17、L20、L21、L22,对照(CK)品种为同等栽培条件下的福鼎大白茶。各优势株系以单行单株条栽,行株距为2.00 m×2.00 m,每个株系种植200株,植株均长势良好,以常规肥水管理。于2021年4月初采摘试验材料春梢一芽二叶,蒸汽杀青固样,装入自封袋密封低温保存备用。
1.2 方法
芽叶性状观察和测定参照NY/T 1312—2007;茶多酚含量和儿茶素组分含量测定参照GB/T 8313—2018;游离氨基酸总量测定参照GB/T 8314—2013;咖啡碱含量测定参照GB/T 8312— 2013;水浸出物含量测定参照GB/T 8305—2013;粗纤维含量测定参照GB/T 8310—2013;氨基酸组分测定参照刘苇等[12]的方法;水溶性碳水化合物含量和维生素C含量测定参照张正竹[13]的方法;蛋白质含量测定参照王艾平等[14]的方法。
1.3 数据处理
参照陈亮等[15]的方法对芽叶描述型性状进行标准化赋值,数量型性状指标按实际测量值进行记录和统计。多重比较分析、相关性分析和主成分分析均采用SPSS 26.0软件进行,参照董胜君等[16]的方法进行综合评价。将7个大厂茶优势株系的16个生化成分的标准化值代入各主成分中,计算各主成分得分,利用模糊隶属函数对各主成分得分进行归一化处理,根据各主成分的贡献率确定权重系数,最后运用综合得分公式计算各个优势株系的综合得分值(IFI)。
=11+22+………+XY(1)
IFI=11+22+………+λF(2)
(1)和(2)式中表示单个主成分得分;表示标准化值;表示各主成分的得分系数;表示对应主成分的贡献率。
2 结果与分析
2.1 大厂茶优势株系芽叶性状分析
通过对7个大厂茶优势株系的21个芽叶性状进行分析。结果表明(表1),7个大厂茶优势株系的芽叶色泽黄绿,芽叶茸毛无或少,叶色绿或浓绿,叶身大多内折,叶质一般或硬脆,叶基为楔形或顿形,叶尖主要为渐尖,叶缘多为微波状,叶齿锐度以中或钝为主,叶齿密度以中或密为主,叶齿多以浅为主,叶脉中或粗,叶面隆起或微隆,除L13稍暗外其余株系的叶光泽度为中或强,叶片多以厚为主,叶脉对数6~9对,叶长变化范围为7.03~14.67 cm,叶宽变化范围为4.33~5.87 cm,叶面积变化范围为25.24~60.28 cm2,平均叶片长宽比介于2~3之间,大多属椭圆形,除了L12为特大叶种外,其余株系主要以中叶种或大叶种为主。
2.2 大厂茶优势株系主要生化成分分析
通过对7个大厂茶优势株系的9个主要生化成分进行分析。结果表明(表2),7个大厂茶优势株系的水浸出物含量总体较高,变化范围为47.02%~48.70%,均显著低于CK(<0.05)。L20的水浸出物含量最高,为48.70%,其次是L22和L17,均为48.43%,最低是L12,为47.02%;咖啡碱含量变化范围为2.91%~3.42%,均显著低于CK。L22的咖啡碱含量最高,为3.42%,最低是L20,为2.91%;茶多酚含量变化范围为19.03%~ 21.07%,L06、L12、L13、L17、L21和L22均显著高于CK,L20与CK差异不显著。L21的茶多酚含量最高,为21.07%。最低是L20,为19.03%;游离氨基酸含量变化范围为4.07%~4.84%,L17的游离氨基酸含量显著高于CK,L06、L20、L21与CK差异不显著,L12、L13和L22显著低于CK。L17的游离氨基酸含量最高,为4.84%,其次是L20,为4.78%,最低是L13,为4.07%;粗纤维含量变化范围为7.43%~8.38%,均显著低于CK。L21的粗纤维含量最高,为8.38%,其次是L12,为8.36%,最低是L17和L22,均为7.43%;维生素C含量变化范围为208.00~332.00 mg/100 g,均显著低于CK。L13的维生素C含量最高,为332.00 mg/100 g,其次是L20,为311 mg/100 g,最低是L22,仅为208.00 mg/100 g;蛋白质含量变化范围为33.82~ 35.07 mg/100 g,L06、L12、L17、L20和L22显著高于CK,L13、L21与CK差异不显著。L20的蛋白质含量最高,为35.07 mg/100 g,其次是L06和L22,均为34.46 mg/100 g,最低是L13,为33.82 mg/100 g;水溶性碳水化合物含量变化范围为3.23%~3.85%, L17、L20和L22与CK差异不显著,L06、L12、L13和L21显著低于CK。L17的水溶性碳水化合物含量最高,为3.85%。7个大厂茶优势株系的酚氨比变化范围为3.98~5.04,除L20的酚氨比与CK差异不显著外,其余株系均显著高于CK。
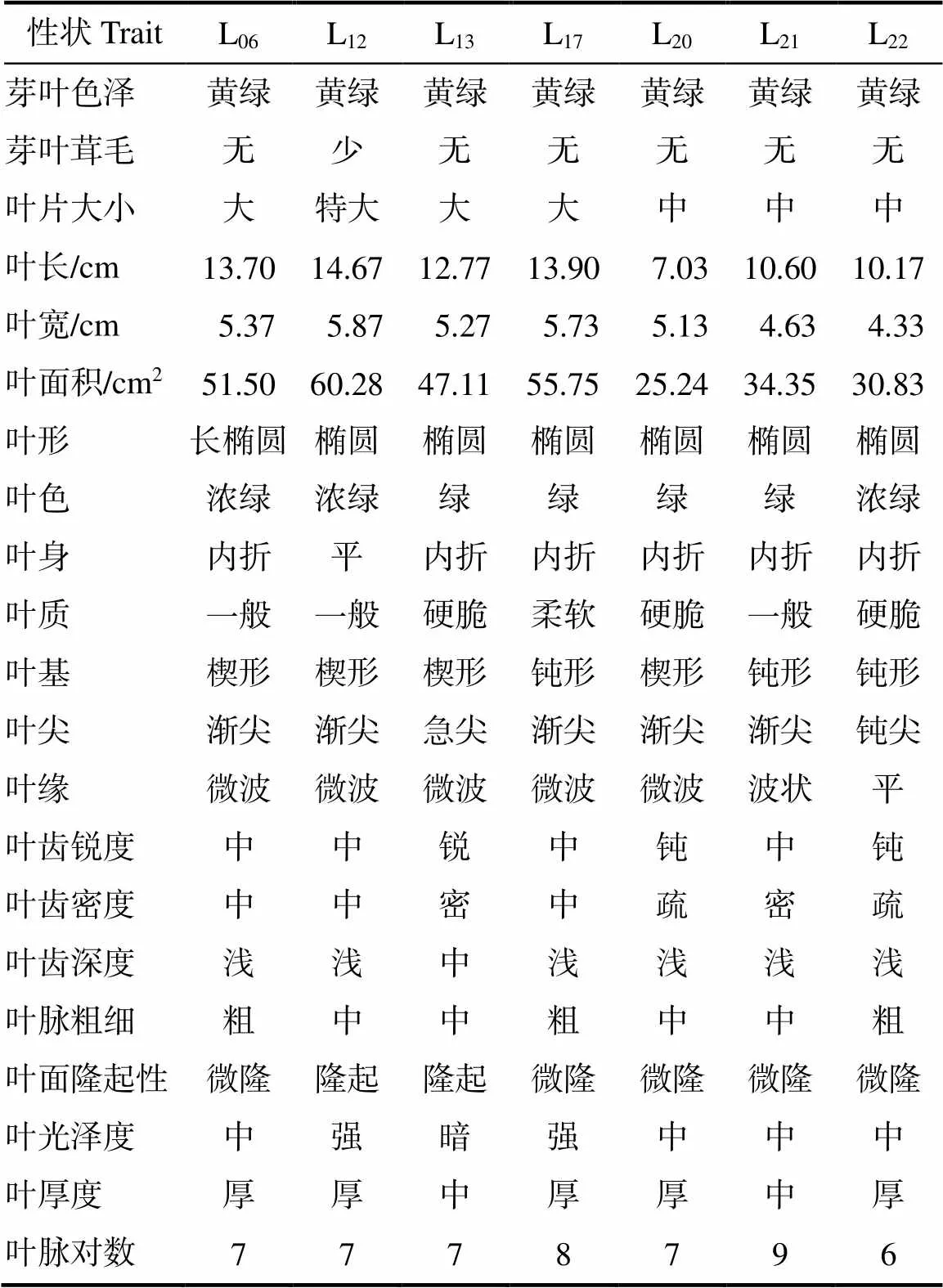
表1 21个芽叶性状分析
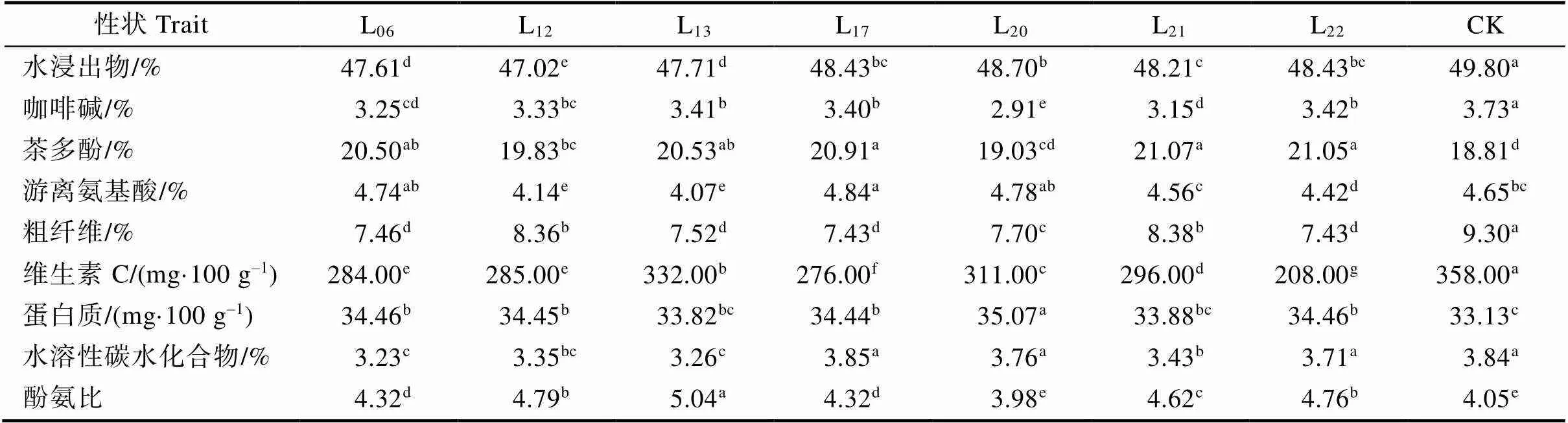
表2 主要生化成分分析
注:同列不同小写字母表示处理间差异显著(<0.05)。
Note: Different lowercase letters after the same column indicate significant difference (<0.05).
2.3 大厂茶优势株系儿茶素组分分析
通过对7个大厂茶优势株系的主要儿茶素组分进行分析。结果表明(表3),7个大厂茶优势株系中主要儿茶素组分含量大小顺序依次为C>ECG>EC>EGCG>EGC。儿茶素总量变化范围为14.68%~17.21%,除L12的儿茶素总量与CK差异不显著外,其余大厂茶优势株系均显著高于CK。L22的儿茶素总量最高,为17.21%,其次是L13和L17,分别为17.14%和16.77%,最低是L12,为14.68%;C含量变化范围为5.25%~9.02%,在儿茶素总量中的占比为35.76%~52.63%,7个大厂茶优势株系的C含量均显著高于CK,为CK的2.72~4.67倍。L13的C含量最高,为9.02%,其次是L22和L17,分别为8.83%和7.50%,最低是L12,为5.25%;ECG含量变化范围为6.11%~7.64%,在儿茶素总量中的占比为35.50%~ 52.04%,7个大厂茶优势株系的ECG含量均显著高于CK,为CK的2.51~3.14倍。L12的ECG含量最高,为7.64%,其次是L13,为7.41%,最低是L22,为6.11%;EGCG含量变化范围为0.08%~ 1.44%,在儿茶素总量的占比为0.47%~8.37%,7个大厂茶优势株系的EGCG含量均显著低于CK。为CK的0.01~0.18倍。L22的EGCG含量最高,为1.44%,其次是L20,为1.11%,最低是L13,仅为0.08%;EC含量变化范围为0.45%~1.48%,在儿茶素总量的占比为2.63%~9.07%,L06、L12、L17和L20的EC含量显著高于CK,L13和L21显著低于CK,L22与CK差异不显著。L06的EC含量最高,为1.48%,其次是L12和L17,均为1.39%,最低是L13,为0.45%;EGC含量变化范围为0.05%~0.31%,在儿茶素总量的占比为0.34%~ 2.10%,7个大厂茶优势株系的EGC含量均显著低于CK,为CK的0.04~0.23倍。L21的EGC含量最高,为0.31%,最低是L12,为0.05%;酯型儿茶素总量变化范围为7.49%~7.99%,非酯型儿茶素总量变化范围为6.69%~ 9.66%。其中,C含量占非酯型儿茶素的85.80%以上,是最主要的非酯型儿茶素组分,ECG占酯型儿茶素的90.26%以上,是最主要的酯型儿茶素组分。7个大厂茶优势株系的儿茶素品质指数存在较大差异,其变化范围为2471~15980,均显著高于CK,为CK的3.14~20.33倍。
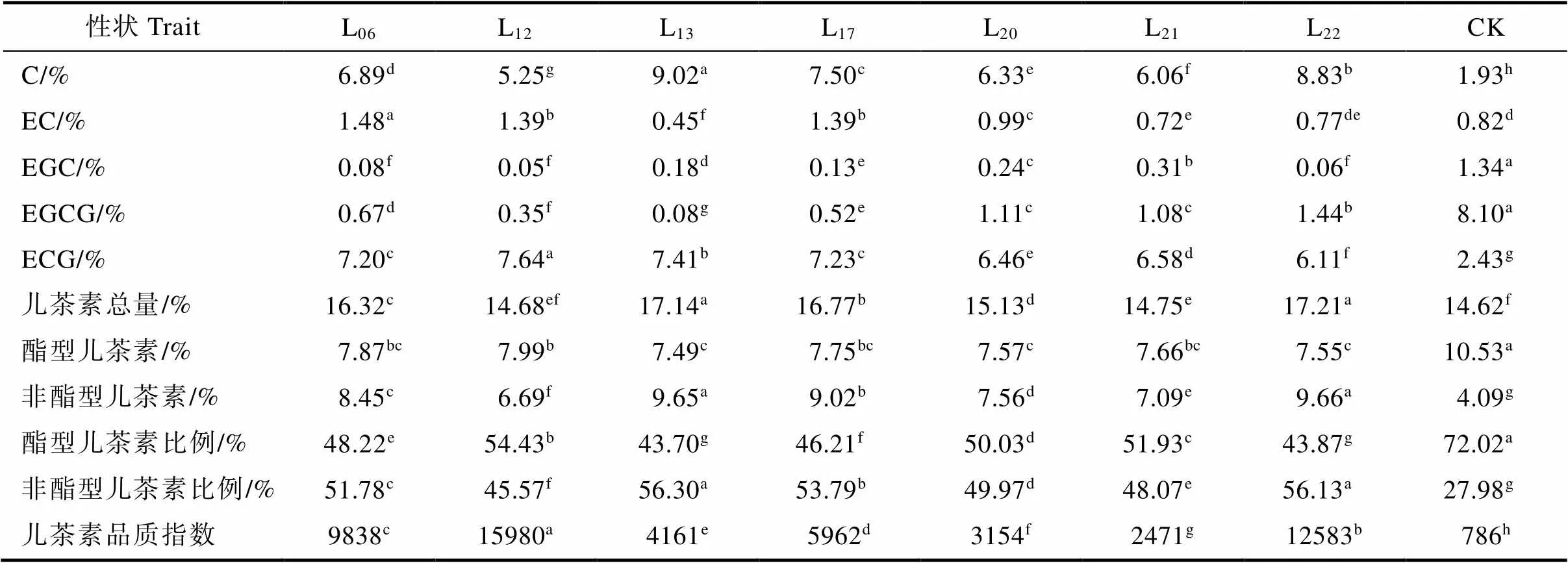
表3 主要儿茶素组分分析
注:C为(+)-儿茶素;EC为(–)-表儿茶素;EGC为(–)-表没食子儿茶素;EGCG为(–)-表没食子儿茶素没食子酸酯;ECG为(–)-表儿茶素没食子酸酯;酯型儿茶素为EGCG+ECG;非酯型儿茶素为C+EC+EGC;儿茶素品质指数=(EGCG+ECG)×100/EGC。
Note: C is (+)-catechin; EC is (–)-epicatechin; EGC is (–)-epigallocatechin; EGCG is (–)-epigallocatechin-3-gallate; ECG is (–)-epicatechin-3-gallate; Ester catechins is EGCG+ECG; Non-ester catechins is C+EC+EGC; Catecin quality index=(EGCG+ECG) ×100/EGC.
2.4 大厂茶优势株系氨基酸组分分析
通过对7个大厂茶优势株系的氨基酸组分进行分析。结果表明(表4),7个大厂茶优势株系共有19种氨基酸被检出,除了在L06中检测到15种外,在其余6个株系检测到18种。7个大厂茶优势株系含量较高的5个氨基酸组分依次是茶氨酸>谷氨酸>精氨酸>天冬氨酸>丝氨酸,占氨基酸总量的82.14%以上。7个大厂茶优势株系氨基酸总量变化范围为2.52%~3.31%,L06、L17、L20和L22的氨基酸总量显著高于CK,其余3个大厂茶优势株系显著低于CK。L06的氨基酸总量最高,为3.31%,其次是L20和L22,分别为3.26%和3.17%,最低是L12,仅为2.52%;茶氨酸含量变化范围为1.48%~2.01%,在氨基酸总量中的占比最高,为55.02%~61.35%,L06、L17、L20和L22的茶氨酸含量显著高于CK,其余大厂茶优势株系显著低于CK。L06的茶氨酸含量最高,为2.01%,其次是L20和L22,分别为1.94%和1.85%,最低是L13,为1.48%;谷氨酸含量变化范围为0.33%~0.46%,在总量中占比为11.75%~14.74%,L06、L17和L20的谷氨酸含量显著高于CK,L12、L13和L21显著低于CK。L17的谷氨酸含量最高,为0.46%,最低是L12,为0.33%;精氨酸含量变化范围为0.17%~0.42%,在总量中占比为6.77%~15.61%,L13的谷氨酸含量显著高于CK,其余大厂茶优势株系均显著低于CK;天冬氨酸含量变化范围为0.14%~0.20%,在总量中占比为4.44%~6.15%,L20的谷氨酸含量显著高于CK,L12、L21和L22显著低于CK;丝氨酸含量变化范围为0.04%~0.08%,在总量中占比为1.49%~2.56%,L12和L13的丝氨酸含量显著低于CK,其余大厂茶优势株系与CK差异不显著。其余氨基酸组分在7个大厂茶优势株系中含量极小,其中磷乙醇胺、组氨酸、胱氨酸等含量仅为0.01%。
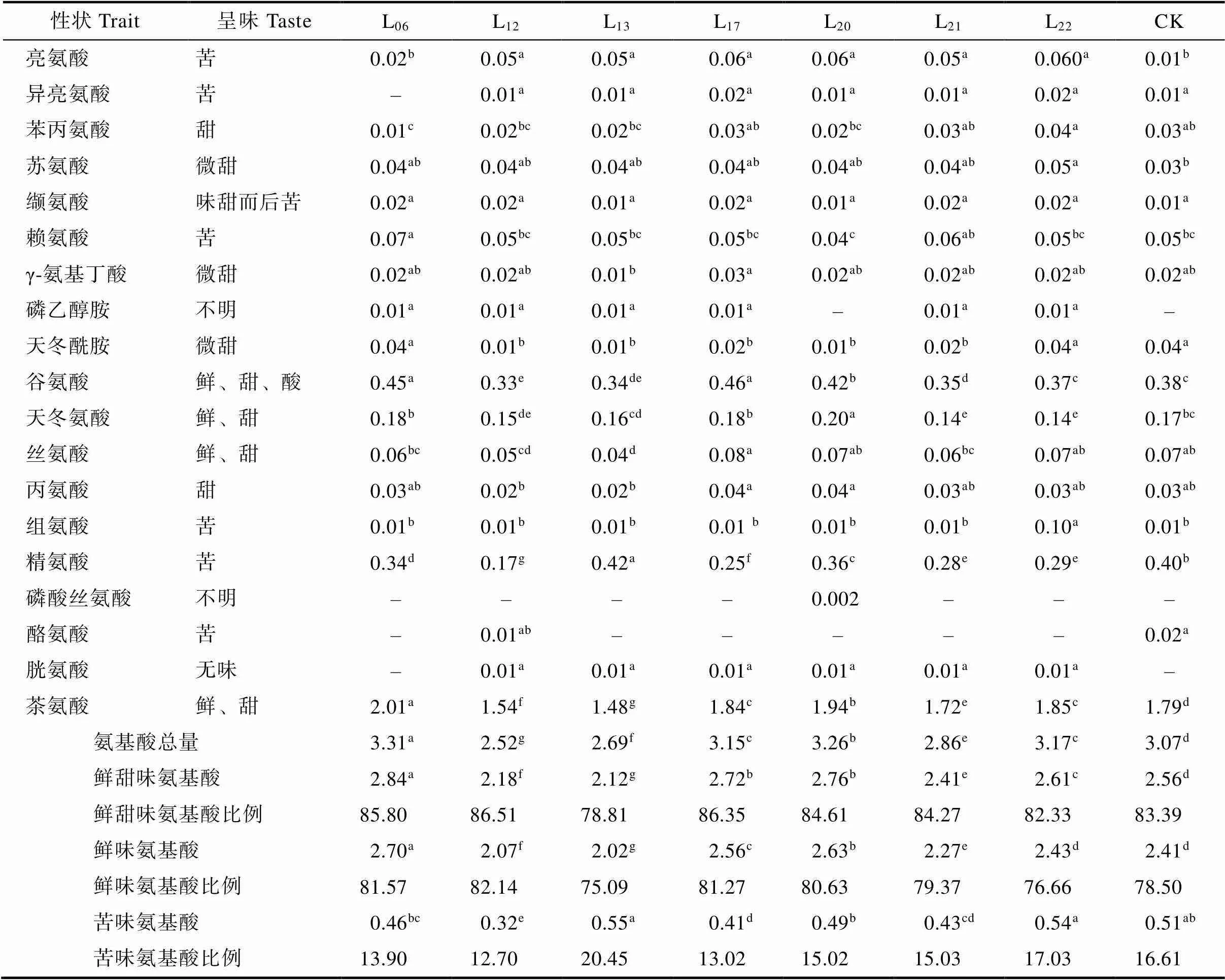
表4 氨基酸组分分析
注:–表示未检出。同列不同小写字母表示处理间差异显著(<0.05)。
Note:– indicates that it has not been checked out. Different lowercase letters after the same column indicate significant difference (<0.05).
7个大厂茶优势株系中呈鲜甜味的氨基酸组分有茶氨酸、苏氨酸、γ-氨基丁酸、天冬酰胺、谷氨酸、天冬氨酸、丝氨酸、丙氨酸和苯丙氨酸9种(表4),在氨基酸总量中占比为78.81%~86.51%。其中,呈鲜味的有茶氨酸、谷氨酸、天冬氨酸、丝氨酸4种,在氨基酸总量中占比为75.09%~ 82.14%;呈苦味的氨基酸组分中除了L06有5种(少异亮氨酸)和L12有7种(多酪氨酸)外,其余5个优势株系都有亮氨酸、异亮氨酸、缬氨酸、赖氨酸、组氨酸、精氨酸等6种,在氨基酸总量中占比为12.70%~24.45%。以上结果说明,在7个优势株系的氨基酸组分中,以呈鲜甜味的组分占的比重大,这是茶汤呈鲜爽味的重要因素。
2.5 芽叶性状与生化成分相关性分析
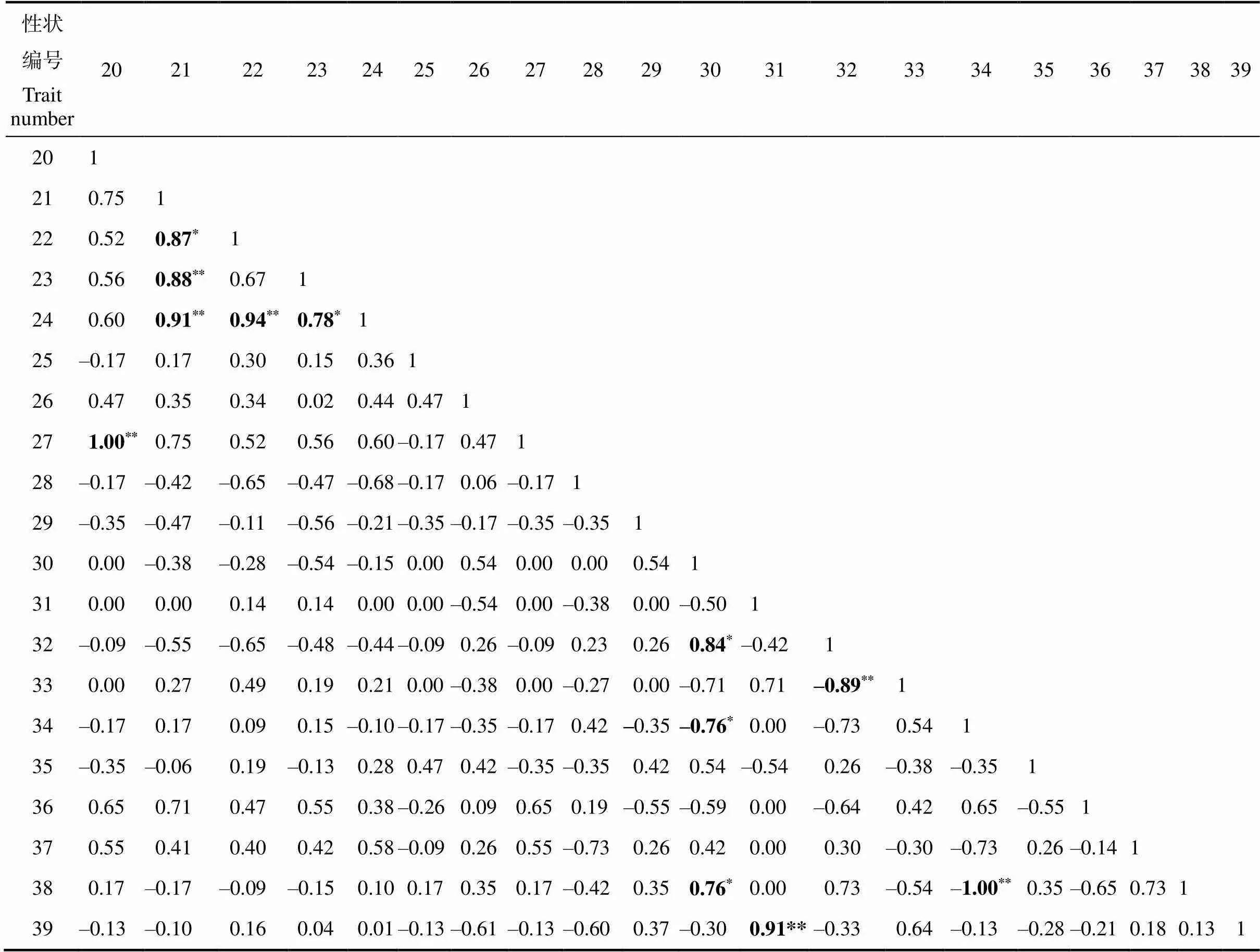
通过对7个大厂茶优势株系的芽叶性状与生化成分进行相关性分析。结果表明(表5),粗纤维与儿茶素总量呈极显著负相关,与非酯型儿茶素和C呈显著负相关,说明随着粗纤维含量逐渐积累,芽叶嫩度越低,儿茶素含量以及非酯型儿茶素越低。茶氨酸和游离氨基酸与叶面隆起性呈极显著负相关,EGCG与叶面隆起性呈显著负相关,说明茶树叶面隆起性越强,其EGCG、茶氨酸和游离氨基酸含量越低。水浸出物与水溶性碳水化合物呈显著正相关,与叶片大小呈显著负相关。EGC与儿茶素品质指数呈极显著负相关,与叶色呈显著负相关。ECG与叶片大小呈极显著正相关,与EGCG呈极显著负相关,与叶齿锐度和水浸出物呈显著负相关。EC与叶光泽度呈显著正相关,叶身与芽叶茸毛呈极显著正相关。叶尖与EGCG呈显著正相关,与维生素C呈极显著负相关。叶齿密度与蛋白质呈极显著负相关。叶齿深度与叶厚度呈极显著负相关,与叶尖呈显著负相关。叶脉对数与叶缘呈极显著正相关。
2.6 主成分分析和综合评价
通过对7个大厂茶优势株系的16个生化成分进行主成分分析。结果表明(表6),前5个主成分的特征值均在1.0以上,累计贡献率达95.68%,说明这5个主成分能代表所有性状所含有的大部分信息。第一主成分特征值最大,为5.13,贡献率为32.09%。绝对值较高的正向特征向量的性状有水浸出物含量、茶氨酸含量、EGCG含量、游离氨基酸含量和水溶性碳水化合物含量,其特征向量值分别为0.89、0.81、0.81、0.80和0.75,绝对值较高的负向特征向量的性状有ECG含量,特征向量值为–0.80。第二主成分特征值为4.05,贡献率为25.33%,绝对值较高的正向特征向量的性状有儿茶素总量、C含量、咖啡碱含量和茶多酚含量,其特征向量值分别为0.93、0.88、0.76、0.66,绝对值较高的负向特征向量的性状有粗纤维含量。第三主成分特征值为3.19,贡献率为19.94%。特征向量正向较高的性状是EC含量、儿茶素品质指数和蛋白质含量,其特征向量值分别为0.86、0.81、0.60,绝对值较高的负向特征向量的性状有EGC含量,特征向量值为–0.80;第四主成分特征值为1.81,贡献率为11.31%,特征向量正向较高的性状是维生素C含量和ECG含量。第五主成分特征值为1.12,贡献率为7.01%,主要由游离氨基酸含量和茶多酚含量决定。
用5个主成分对7个大厂茶优势株系进行综合评价,求得各株系的5个主成分得分,且第一个主成分线性方程为:1=0.891–0.302–0.323+ 0.634–0.145–0.576+0.757–0.018+0.00059+0.0410+0.3111+0.8112–0.8013–0.3614+0.8015+0.8116。其中,1~16表示生化成分的标准化值。根据各主成分的贡献率可得出综合评价函数为:=0.321+0.252+0.203+0.114+0.075,按综合评价函数计算出7个优势株系综合得分值(IFI)和排序。由表7可知,7个大厂茶优势株系的综合得分()的变化范围为0.27~0.67。其中排名前三的依次是L17、L22和L06。

表5 芽叶性状与生化成分相关性分析
续表5 芽叶性状与生化成分相关性分析
Tab. 5 Correlation analysis of leaf bud traits and biochemical components (continued)
注:*表示显著相关(<0.05),**表示极显著相关(<0.01)。1为水浸出物含量;2为粗纤维含量;3为维生素C含量;4为蛋白质含量;5为茶多酚含量;6为咖啡碱含量;7为水溶性碳水化合物;8为儿茶素总量;9为酯型儿茶素含量;10为非酯型儿茶素含量;11为儿茶素品质指数;12为C含量;13为EGC含量;14为EGCG含量;15为EC含量;16为ECG含量;17为游离氨基酸含量;18为茶氨酸含量;19为茎粗;20为芽叶茸毛;21为叶片大小;22为叶长;23为叶宽;24为叶面积;25为叶形;26为叶色;27为叶身;28为叶质;29为叶基;30为叶尖;31为叶缘;32为叶齿锐度;33为叶齿密度;34为叶齿深度;35为叶脉粗细;36为叶面隆起性;37为叶光泽度;38为叶厚度;39为叶脉对数。
Note:*indicates significant correlation (<0.05),**indicates extremely significant correlation (<0.01). 1 is water extracts content; 2 is crude fiber content; 3 is Vc content; 4 is protein content; 5 is tea polyphenols content; 6 is caffeine content; 7 is water soluable carbohydrate content; 8 is total catechin content; 9 is ester catechins content; 10 is non-ester catechins content; 11 is catecin quality index; 12 is C content; 13 is EGC content; 14 is EGCG content; 15 is EC content; 16 is ECG content; 17 is amino acids content; 18 is theanine content; 19 is stem thick; 20 is bud leaf pastel; 21 is leaf size; 22 is leaf long; 23 is leaf width; 24 is leaf area; 25 is leaf shape; 26 is leaf color; 27 is leaf body; 28 is leaf quality; 29 is leaf-based part; 30 is leaf tip; 31 is leaf margin; 32 is leaf tooth sharpness; 33 is leaf tooth density; 34 is leaf tooth depth; 35 is leaf vein thickness; 36 is leaf protrusion; 37 is leaf gloss; 38 is leaf thickness; 39 is leaf vein logarithm.
2.7 优良种质资源鉴定
通过对7个大厂茶优势株系生化成分的综合分析可知。结果表明(表8),7个大厂茶优势株系的水浸出物、游离氨基酸、C和ECG的含量均较高。参考《NY/T 2031—2011农作物优异种质资源评价规范茶树》,7个大厂茶优势株系的游离氨基酸含量均大于4.0%,L06、L13、L17、L21和L22的茶多酚含量均大于20%,为潜在的优良大厂茶种质资源。陈正武等[17]研究认为,酚氨比<4.5为适制绿茶,4.5<酚氨比<6.5为红绿茶兼制,酚氨比>6.5为适制红茶。本研究中,L06、L17和L20的游离氨基酸含量>4.5%、酚氨比<4.5、儿茶素品质指数均显著高于CK,说明该株系具有适制绿茶的潜在特性,而L12、L13、L21和L22的酚氨比介于4.5~6.5之间,具有红绿茶兼制的潜在特性。
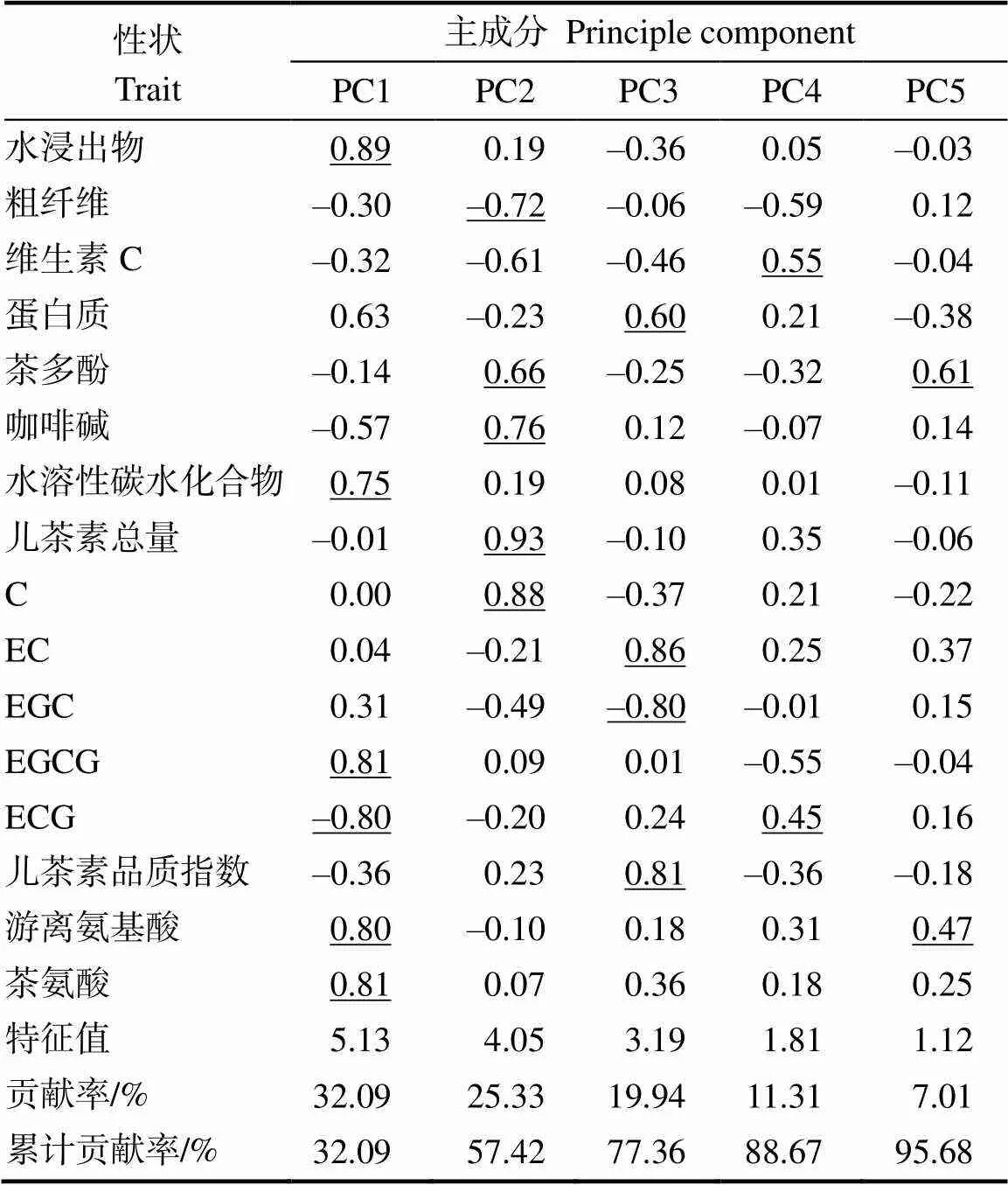
表6 主要生化成分主成分分析
注: 下划线表示每组选取特征向量绝对值较大的值,以反映其对应数量型性状的贡献率。
Note:The underline indicates that each group selects the largest values of the absolute value of the eigenvector to reflect the contribution.
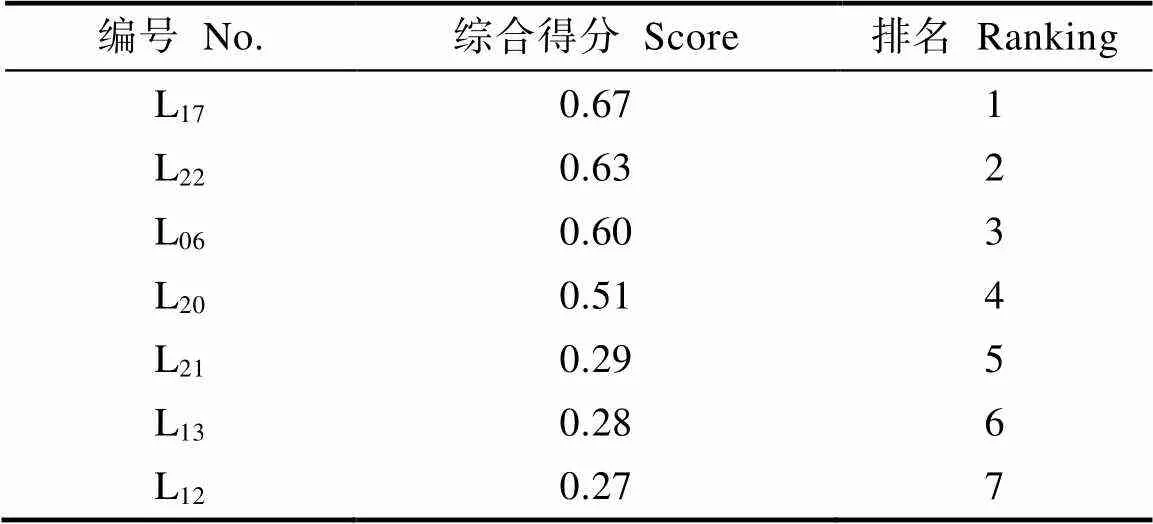
表7 7个大厂茶优势株系综合得分(IFI)
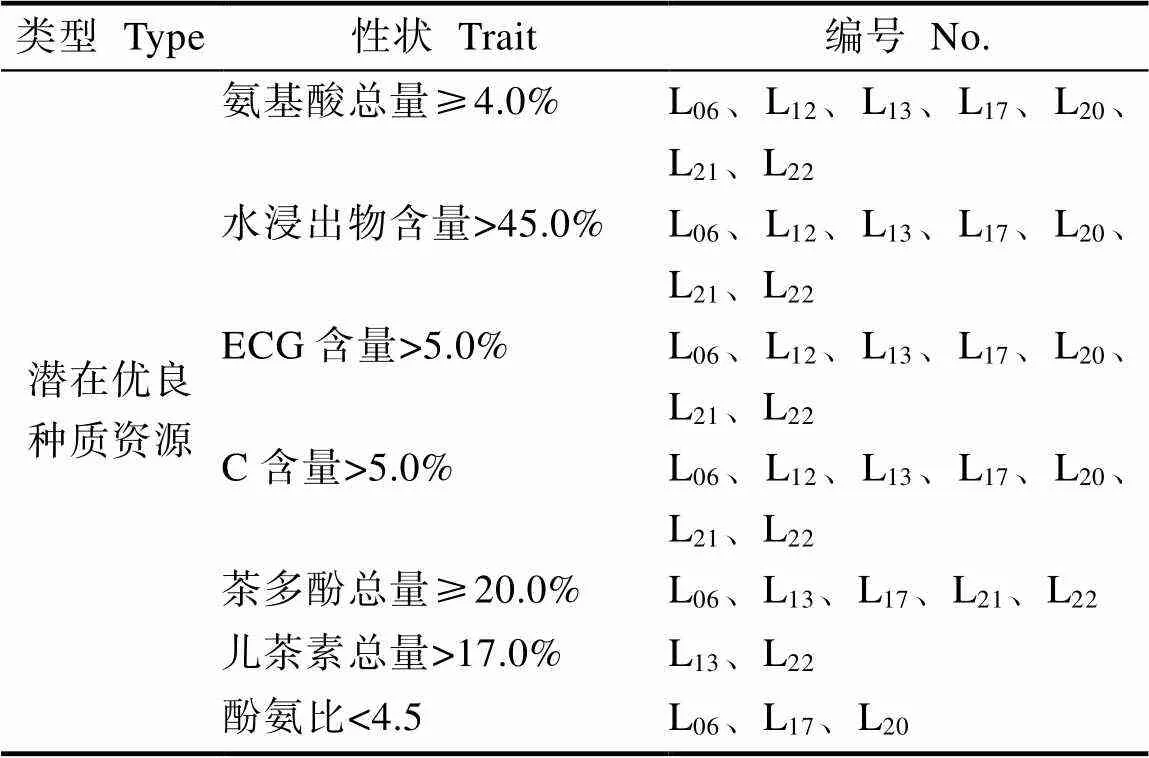
表8 大厂茶优势株系鉴定
3 讨论
植物的表型性状是基因与环境互作的综合体现,是鉴别种质资源分类及新品种选育的重要物质基础,具有直接、便捷等优点[18],在茶树新品种选育中应用较为广泛[19-20]。大厂茶种质资源是特产于滇黔桂交接地区的国家优质重点保护茶种植物,在茶树植物学的起源和演化中占有重要位置,对其进一步挖掘和鉴定评价利用具有重要的意义。本研究对大厂茶优势株系的芽叶性状调研显示,7个大厂茶优势株系主要为乔木大叶种、芽叶色泽黄绿无茸毛、叶形椭圆、叶身内折、叶面微隆、叶齿浅、叶尖多渐尖、叶光泽度好。
茶叶生化成分的含量是茶叶品质的重要影响因素,可根据含量组成和比例初步判定茶叶品质的优劣以及适制性,从而提高茶树种质资源的利用效率、价值和经济利益。林蒙嘉[21]研究发现大厂茶的咖啡碱、氨基酸和茶多酚等含量均较为丰富;刘苇等[12]研究发现大厂茶野生茶树的游离氨基酸和挥发性芳香物质的种类及各组分含量丰富且多样性,这可能是大厂茶茶汤呈鲜爽味和甜味,香气呈花香味、果香味、木香味、青草气、脂肪香味的重要因素。本研究显示7个大厂茶优势株系茶多酚、游离氨基酸及儿茶素组分等含量均较高,属优良茶树种质资源,其水浸出物含量的变异范围为47.02%~48.70%,蛋白质含量变化范围为33.82~35.07 mg/100 g。水浸出物可反映茶叶中可溶性物质总量,标志着茶汤的厚薄、滋味的浓强程度[22],蛋白质反映茶汤清亮程度,在茶汤中呈胶体状态[23],说明7个大厂茶优势株系内含成分较丰富,这一研究结果与上述报道的结果基本一致[24]。
儿茶素作为茶多酚的主体成分,占茶多酚总量的60%~80%,是茶叶中最重要的功能成分[25],决定茶叶品质及其功效,其组成差异也是作为茶树种质资源进化类型的一个重要指标。XIA等[26]研究认为茶组植物与其他山茶属植物中单体儿茶素积累存在显著差异,特别是酯型儿茶素,在非茶组茶属植物中几乎检测不到;JIN等[27]研究认为秃房茶()、四球茶()、大厂茶()等四室或五室茶类的儿茶素积累与其他茶组植物不同,相比于栽培种,其EGC和EGCG含量极低;林蒙嘉[21]研究认为大厂茶儿茶素组成中,EC、C含量高,EGCG含量低,认为其是较原始的茶树类型。本研究中7个大厂茶优势株系的儿茶素积累及单体配比与前人的研究结果一致,儿茶素单体含量大小依次为C>ECG> EC>EGCG>EGC,其ECG含量是福鼎大白茶的2.51~3.14倍,是最主要的酯型儿茶素。儿茶素品质指数能够较为准确地反映出茶叶的嫩度与茶叶品质,儿茶素品质指数越高,芽叶持嫩性越好且茶叶品质越高[28]。本研究中7个大厂茶优势株系的儿茶素品质指数均较高,最高可达对照的20.33倍,说明其持嫩性好、品质较好。综上所述,这7个大厂茶优势株系属高水浸出物、高游离氨基酸、高ECG茶树种质资源,同时具有较高的持嫩性优势,可作为特异种质资源进行开发利用,为研发和创制健康功能性饮品提供基础材料。
相关性分析是用来研究不同性状之间密切程度的一种统计分析方法[29],植物体内同一连锁基因群所控制的不同表型性状往往表现出相关关系[30]。胡月等[31]研究发现南涧地区古茶树资源的26个表型性状呈极显著正相关的有31对性状,呈极显著负相关的有2对性状。刘声传等[5]研究发现,茶树新梢色素含量与其类胡萝卜素、游离氨基酸、叶绿素、花青素等主要生化成分含量关系密切。本研究中,芽叶性状与主要生化成分之间存在紧密联系,表现出极显著正相关的有12对性状,呈显著正相关的有13对性状,呈极显著负相关的有9对性状,呈显著负相关的有10对性状。其中ECG含量与叶片大小和叶宽呈极显著正相关,与EGCG含量呈极显著负相关,与水浸出物含量和叶齿锐度呈显著负相关。C含量与儿茶素总量和非酯型儿茶素呈极显著正相关,与粗纤维含量呈显著负相关。这些性状控制的基因可能存在紧密连锁关系[32],可以作为进一步对ECG、C功能基因挖掘的理论依据。
主成分分析可将多个变量化为少数几个指标,通过简化描述指标,获得解释方差的主要性状,提高大厂茶优势株系的评价与选育鉴定工作的效率[33]。本研究基于16个生化成分对7个大厂茶优势株系进行综合评价显示,L12、L13、L20和L21的氨基酸、水浸出物和ECG含量均较高,品质较好,L06、L17和L22的综合品质表现优异。综上所述,7个大厂茶优势株系均具有潜在和较好的推广利用价值,但还需要进一步开展茶树良种选育品比试验,明确其性状的稳定性,为大厂茶新品种(系)的选育奠定基础。
[1] 庄晚芳. 茶树原产于我国何地[J]. 浙江农业大学学报, 1981(3): 115-119.
ZHUANG W F. Tea tree native to China[J]. Journal of Zhejiang Agricultural University, 1981(3): 115-119. (in Chinese)
[2] 牛素贞, 宋勤飞, 樊卫国, 陈正武. 干旱胁迫对喀斯特地区野生茶树幼苗生理特性及根系生长的影响[J]. 生态学报, 2017, 37(21): 7333-7341.
NIU S Z, SONG Q F, FAN W G, CHEN Z W. Effects of drought stress on leaf physiological characteristics and root growth of the clone seedlings of wild tea plant[J]. Acta Ecologica Sinica, 2017, 37(21): 7333-7341. (in Chinese)
[3] 陈 亮, 虞富莲, 童启庆. 关于茶组植物分类与演化的讨论[J]. 茶叶科学, 2000(2): 89-94.
CHEN L, YU F L, TONG Q Q. Discussions on phylogenetic classification and evolution of[J]. Journal of Tea Science, 2000(2): 89-94. (in Chinese)
[4] 虞富莲. 茶源贵州依据充分[J]. 当代贵州, 2019(27): 14.
YU F L. Sufficient evidence on Guizhou is the cradle of tea[J]. Guizhou Today, 2019(27): 14. (in Chinese)
[5] 刘声传, 曹 雨, 鄢东海, 魏 杰, 赵华富, 段学艺. 贵州野生茶树资源地理分布和形态特征与气候要素的关系[J]. 茶叶科学, 2013, 33(6): 517-525.
LIU S C, CAO Y, YANG D H, WEI J, ZHAO H F, DUAN X Y. Geographical distribution and morphology of wild tea germplasm resources in Guizhou and its relationship with climatic factors[J]. Journal of Tea Science, 2013, 33(6): 517-525. (in Chinese)
[6] 张宏达. 茶叶植物资源的订正[J]. 中山大学学报(自然科学版), 1984(1): 3-14.
ZHANG H D. Revision of tea plant resources[J]. Journal of Zhongshan University (Natural Science Edition), 1984(1): 3-14. (in Chinese)
[7] 杨 凤, 刘 霞, 尹 杰, 宋勤飞, 陈 娟, 牛素贞, 刘进平. 贵州野生茶树种质资源的主要生化成分及抗旱性评价[J]. 西南农业学报, 2018, 31(6): 1122-1127.
YANG F, LIU X, YIN J, SONG Q F, CHEN J, NIU S Z, LIU J P. Major Biochemical components and drought resistance of wild tea germplasm eesources in Guizhou[J]. Southwest China Journal of Agricultural Sciences, 2018, 31(6): 1122-1127. (in Chinese)
[8] 牛素贞, 安红卫, 刘文昌, 陈正武. 贵州古茶树种质资源的生育期及春梢萌发特性[J]. 贵州农业科学, 2020, 48(3): 10-15.
NIU S Z, AN H W, LIU W C, CHEN Z W. Growth stage and spring shoot germination characteristics of ancient tea germplasm resources in Guizhou[J]. Guizhou Agricultural Sciences, 2020, 48(3): 10-15. (in Chinese)
[9] NIU S Z, SONG Q F, KOIWA H, QIAO D H, ZHAO D G, CHEN Z W, LIU X, WEN X P. Genetic diversity, linkage disequilibrium, and population structure analysis of the tea plant () from an origin center, Guizhou plateau, using genome-wide SNPs developed by genotyping-by-sequencing[J]. BMC Plant biology, 2019, 19: 328.
[10] 牛素贞, 宋勤飞, 安红卫, 黄 政, 刘 霞, 赵德刚, 陈正武. 贵州古茶树种质资源基于形态特征的多样性研究[J]. 浙江农业学报, 2019, 31(10): 1689-1699.
NIU S Z, SONG Q F, AN H W, HUANG Z, LIU X, ZHAO D G, CHEN Z W. Diversity of ancient tea germplasm based on morphological characteristics in Guizhou[J]. Acta Agriculturae Zhejiangensis, 2019, 31(10): 1689-1699. (in Chinese)
[11] 尹 杰, 牛素贞, 宋勤飞, 刘进平, 陈 曦. 黔西南州野生茶树的性状特征及品质分析[J]. 浙江农业学报, 2013, 25(4): 782-786.
YIN J, NIU S Z, SONG Q F, LIU J P, CHEN X. Analysis on the leaf bud characters and biochemical components in the spring shoots of wild tea plant from Qianxinan[J]. Acta Agriculturae Zhejiangensis, 2013, 25(4): 782-786. (in Chinese)
[12] 刘 苇, 邓朝义, 陈 兴, 卢永成, 廖德胜. 大厂茶茶叶中游离氨基酸及挥发性芳香物质分析[J]. 浙江林业科技, 2021, 41(3): 1-14.
LIU W, DENG C Y, CHEN X, LU Y C, LIAO D S. Determination of free amino acid and volatile aromatic compound in[J]. Journal of Zhejiang Forestry Science and Technology, 2021, 41(3): 1-14. (in Chinese)
[13] 张正竹. 茶叶中维生素C含量的测定. 茶叶生物化学实用教程[M]. 北京: 中国农业出版社, 2009: 60-62.
ZHANG Z Z. Determination of vitamin C content in tea. Experimental course of tea biochemistry[M]. Beijing: China Agricultural Press, 2009: 60-62. (in Chinese)
[14] 王艾平, 周丽明. 考马斯亮蓝法测定茶籽多糖中蛋白质含量条件的优化[J]. 河南农业科学, 2014, 43(3): 150-153.
WANG A P, ZHOU L M. Determination of protein content inseed polysaccharides by Coomassie brilliant blue method[J]. Journal of Henan Agricultural Sciences, 2014, 43(3): 150-153. (in Chinese)
[15] 陈 亮, 杨亚军, 虞富莲. 茶树种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2005.
CHEN L, YANG Y J, YU F L. Description specification and data standard of tea germplasm resources[M]. Beijing: China Agricultural Press, 2005. (in Chinese)
[16] 董胜君, 孙永强, 陈建华, 卢彩云, 刘权钢, 刘立新. 野杏无性系表型性状多样性分析及综合评价[J]. 植物遗传资源学报, 2020, 21(5): 1156-1166.
DONG S J, SUN Y Q, CHEN J H, LU C Y, LIU Q G, LIU L X. Phenotypic traits diversity analysis and comprehensive evaluation ofvar.clones[J]. Journal of Plant Genetic Resources, 2020, 21(5): 1156-1166. (in Chinese)
[17] 陈正武, 陈 娟, 龚 雪, 张小琴. 28份贵州茶树种质资源的生化成分多样性分析[J]. 西南农业学报, 2015, 28(4): 1517-1523.
CHEN Z W, CHEN J, GONG X, ZHANG X Q. Diversity analysis for biochemical components of 28 tea germplasm resources in Guizhou[J]. Southwest China Journal of Agricultural Sciences, 2015, 28(4): 1517-1523. (in Chinese)
[18] 段学艺, 胡华健, 王家伦, 曹 雨, 赵华富. 黔西南州大茶树资源现状及其保护与利用[J]. 农技服务, 2011, 28(12): 1736-1737.
DUAN X Y, HU H J, WANG J L, CAO Y, ZHAO H F. Present situation of protection and utilization of tea resources in southwestern Guizhou[J]. Agricultural Technology Services, 2011, 28(12): 1736-1737. (in Chinese)
[19] 陈红霖, 胡亮亮, 杨 勇, 郝曦煜, 李姝彤, 王素华, 王丽侠, 程须珍. 481份国内外绿豆种质农艺性状及豆象抗性鉴定评价及遗传多样性分析[J]. 植物遗传资源学报, 2020, 21(3): 549-559.
CHEN H L, HU L L, YANG Y, HE X Y, LI S T, WANG S H, WANG L X, CHENG X Z. Evaluation and genetic diversity analysis of agronomic traits and bruchid resistance using 481 worldwide mungbean germplasms[J]. Journal of Plant Genetic Resources, 2020, 21(3): 549-559. (in Chinese)
[20] 杨培迪, 刘 振, 赵 洋, 成 杨, 宁 静, 刘 勇, 黄建安, 杨 阳. 基于农艺性状和SSR标记亲缘关系分析的江华苦茶品系评价[J]. 分子植物育种, 2021, 19(7): 2402-2409.
YANG P D, LIU Z, ZHAO Y, CHENG Y, NING J, LIU Y, HUANG J A, YANG Y. Evaluation of Jianghua kucha tea strains based on agronomic and SSR molecular marker relationship analysis[J]. Molecular Plant Breeding, 2021, 19(7): 2402-2409. (in Chinese)
[21] 林蒙嘉. 普白大树茶资源调查[J]. 茶叶, 1983(2): 14-16.
LIN M J. Investigation of Pubai big tree tea resources[J]. Tea, 1983(2): 14-16. (in Chinese)
[22] 李志伟. 基于主成分分析法的茶叶特征性指标分类[J]. 安徽农业科学, 2014, 42(7): 2109-2110, 2160.
LI Z W. PCA-based classification of characteristic indicators of tea[J]. Journal of Anhui Agricultural Sciences, 2014, 42(7): 2109-2110, 2160. (in Chinese)
[23] 余 浩, 唐 敏, 黄升谋. 冲泡条件对绿茶水浸出物含量及感官品质的影响研究[J]. 绿色科技, 2016(24): 137-140.
YU H, TANG M, HUANG S M. Effect of brewing condition on water extracts and sensory quality of tea[J]. Journal of Green Science Technology, 2016(24): 137-140. (in Chinese)
[24] 张 婷, 刘海燕, 邹天才. 贵州8种野生山茶叶片主要化学成分的含量[J]. 贵州农业科学, 2010, 38(11): 78-80.
ZHANG T, LIU H Y, ZOU T C. Main chemical components in leaves of 8 wildspecies in Guizhou[J]. Guizhou Agricultural Science, 2010, 38(11): 78-80. (in Chinese)
[25] 谢志英, 黄立文, 王秀华, 满红平, 赵永祥, 胡文东, 桂丽萍. 云南大叶种茶不同品种儿茶素组分含量分析[J]. 中国农学通报, 2014, 30(19): 146-150
XIE Z Y, HUANG L W, WANG X H, MAN H P, ZHAO Y X, HU W D, GUI L P. The catechins constituent content analysis of different cultivars ofvarin Yunnan[J]. Chinese Agricultural Science Bulletin, 2014, 30(19): 146-150. (in Chinese)
[26] XIA E H, ZHANG H B, SHENG J, LI K, ZHANG Q J, KIM C, ZHANG Y, LIU Y, ZHU T, LI W, HUANG H, TONG Y, HONG N, SHI C, SHI C, JIANG J J, MAO S Y, JIAO J Y, ZHANG D, ZHAO Y, ZHAO Y J, ZHANG L P, LIU Y L, LIU B Y, YU Y, SHAO S F, NI D J, EICHLER E E, GAO L Z. The tea tree genome provides insights into tea flavor and independent evolution of caffeine biosynthesis[J]. Molecular Plant, 2017, 10(6): 866-877.
[27] JIN J Q, LIU Y F, MA C L, MA J Q, HAO W J, XU Y X, YAO M Z, CHEN L. A novel F3'5'H allele with 14 bp deletion is associated with high catechin index trait of wild tea plants and has potential use in enhancing tea quality[J]. Journal of Agricultural and Food Chemistry, 2018, 66(40): 10470-10478.
[28] 阮宇成, 程启坤. 茶儿茶素的组成与绿茶品质的关系[J]. 园艺学报, 1964(3): 287-300.
RUAN Y C, CHENG Q K. Relationship between the composition of tea catechins and the quality of green tea[J]. Journal of Horticulture, 1964(3): 287-300. (in Chinese)
[29] 张豪杰, 郝心愿, 周 超, 王 璐, 王新超, 杨亚军, 曾建明, 孙冷雪, 戴居会, 向 俊, 罗 鸿, 王朝阳, 张贤贵, 刘 涛. 富硒区茶树鲜叶中硒累积与土壤因子的相关性分析[J]. 茶叶科学, 2020, 40(4): 465-477.
ZHANG H J, HAO X Y, ZHOU C, WANG L, WANG X C, YANG Y J, ZENG J M, SUN L X, DAI J H, XIANG J, LUO H, WANG C Y, ZHANG X G, LIU T. Correlation analysis between selenium accumulation in tea leaves and soil factors in selenium-rich areas[J]. Journal of Tea Science, 2020, 40(4): 465-477. (in Chinese)
[30] 陈涛林. 广西元宝山一种特异茶饮植物的系统学鉴定与综合评价研究[D]. 长沙: 湖南农业大学, 2019.
CHEN T L. Systematic identification and comprehensive evaluation of a specific tea plant in Yuanbaoshan, Guangxi[D]. Changsha: Hunan Agricultural University, 2019. (in Chinese)
[31] 胡 月, 陈洪宇, 扈月豪, 罗义菊, 蓝增全, 石 明. 南涧县古茶树形态多样性及相关性分析[J/OL]. 分子植物育种: 1-18[2022-11-20]. http://kns.cnki.net/kcms/detail/46.1068.s. 2022 0119.1901.004.html.
HU Y, CHEN H Y, HU Y H, LUO Y J, LAN Z Q, SHI M. Analysis on morphological diversity and correlation of ancient tea trees in Nanjian county[J/OL]. Molecular Plant Breeding: 1-18[2022-11-20]. http://kns.cnki.net/kcms/detail/46. 1068.s.20220119.1901.004.html. (in Chinese)
[32] 陈洪宇, 陶燕蓝, 罗义菊, 扈月豪, 蓝增全. 西南地区10种茶组植物表型多样性分析[J/OL]. 分子植物育种: 1-17[2022-04-10]. http://kns.cnki.net/kcms/detail/46.1068.s. 20220221.1401.002.html.
CHEN H Y, TAO Y L, LUO Y J, HU Y H, LAN Z Q. Analysis of phenotypic diversity of 10 tea plants in Southwest China[J/OL]. Molecular Plant Breeding: 1-17[2022-04-10]. http://kns.cnki.net/kcms/detail/46.1068.s.20220221.1401. 002.html. (in Chinese)
[33] 张 莹, 曹玉芬, 霍宏亮, 徐家玉, 田路明, 董星光, 齐 丹, 张小双, 刘 超, 王立东. 基于枝条和叶片表型性状的梨种质资源多样性[J]. 中国农业科学, 2018, 51(17): 3353-3369.
ZHANG Y, CAO Y F, HUO H L, XU J Y, TIAN L M, DONG X G, QI D, ZHANG X S, LIU C, WANG L D. Diversity of pear germplasm resources based on twig and leaf phenotypic traits[J]. China Agriculture Science, 2018, 51(17): 3353-3369. (in Chinese)
Identification and Evaluation on Leaf Bud Traits and Biochemical Components of Dominant Strains of
LI Caiyun1, SONG Qinfei1, NIU Suzhen1*, ZHAO Zhifei1, HE Yingqin1, LI Fang1, MA Anquan2
1. College of Tea Science, Guizhou University, Guiyang, Guizhou 550025, China; 2. Tai’an Bailong Ancient Tea Professional Cooperative, Pu’an, Guizhou 561500, China
In order to scientifically and reasonably select new lines of high-quality, seven dominant strains ofwere used as the materials in this study, ‘Fuding Dabaicha’ as the check, to comprehensively analyze and systematically evaluate the leaf bud traits, basic quality traits, catechin components and amino acid components. The leaf bud traits of the seven lines were medium leaf or large leaf species with slightly elongated yellow-green bud leaves of shallow and blunt leaf teeth, without pilose or less pilose, in elliptic or long elliptic, with hard and crisp texture, and glossy leaf thickness.. Biochemical analysis showed that the water extracts, free amino acids and ECG contents of the seven lines were high, indicating that they were excellent tea germplasm resources.The variation range of water extract content, caffeine content, tea polyphenols content, amino acid content, crude fiber content, vitamin C content, protein content and water-soluble carbohydrate content was 47.02%–48.70%, 2.91%–3.42%, 19.03%–21.07%, 4.07%–4.84%, 7.43%–8.38%, 208.00–332.00 mg/100 g, 33.82–35.07 mg/100 g, 3.23%–3.85%, the variation range of phenol-ammonia ratio was 3.98–5.04. The content of main catechin was in the order of C>ECG>EC>EGCG>EGC, the content of C and ECG was high, which was significantly higher than that of CK, and the variation range was 5.25%–9.02% and 6.11%–7.64% , respectively. The contents of EGCG and EGC were low, which were significantly lower than that of CK, and the variation range was 0.08%–1.44% and 0.05%–0.31%, respectively. A total of 19 amino acids were detected in the seven lines. There were some differences in the types and contents of amino acids among different components. The five amino acids with high content were theanine>glutamic acid>arginine>aspartic acid>serine, accounting for more than 82.14 % of the total amino acids. There was a certain correlation between morphological traits andbochemical components.The results of principal component analysis based on 16 biochemical components showed that 16 biochemical components were simplified to 5 factors, and the cumulative contribution rate was 95.68%, which contained most of the star information of all traits.Comprehensive Evaluation based on biochemical components showed that L17, L22and L06ranked the top three among the seven lines. In this study, dominant lines L06, L17and L20with potential characteristics of suitable green tea and lines L12, L13, L21and L22with potential characteristics both for black and green tea production were identified.
; leaf bud traits; biochemical components;identification
S571.1
A
10.3969/j.issn.1000-2561.2022.12.019
2022-01-17;
2022-04-10
国家重点研发计划项目(No. 2021YFD1200203-1,No. 2021YFD1100307);贵州省教育厅重点领域项目(黔教合KY[2021]042)。
李彩云(1995—),女,硕士研究生,研究方向:茶树种质资源与遗传改良。*通信作者(Corresponding author):牛素贞(NIU Suzhen),E-mail: niusuzhen@163.com。
