旱季降雨格局变化对益智生长和碳氮代谢的影响
2023-01-16周小慧李勤奋程汉亭
胡 雯,周小慧,李勤奋,程汉亭*
旱季降雨格局变化对益智生长和碳氮代谢的影响
胡 雯1,2,3,周小慧2,3,李勤奋2,3,程汉亭2,3*
1. 海南大学生态与环境学院,海南海口 570228;2. 中国热带农业科学院环境与植物保护研究所/海南省热带生态循环农业重点实验室,海南海口 571101;3. 农业农村部热带农业绿色低碳重点实验室,海南海口 571101
益智是我国四大南药之一,其间作在橡胶林下成为胶园最重要的农林复合模式。项目组前期研究发现,旱季降雨不足是橡胶林下益智生长和光合的主要限制因子。然而,目前对益智如何应对旱季降雨格局变化的认识非常有限。为研究旱季降雨格局变化对益智生长和碳氮代谢的影响,在实验中模拟3个降雨量梯度:减雨量(W-)10 mm/月、自然雨量(W)25 mm/月、增雨量(W+)40 mm/月,并设置5 d和10 d的降雨间隔期,通过双因素实验分析益智植株的生长参数、碳水化合物和氮化合物水平以及与碳和氮代谢相关的关键酶活性。结果表明:(1)益智的地上生物量和地下生物量随着降雨量的减少而下降。在所有降水水平下,5 d处理下的生物量均显著高于10 d(<0.05),降雨量减少和降雨间隔期延长均抑制植株生物量的增加。在W-处理下,10 d的叶片含水率比5 d显著增加3.63%;(2)降雨减少改变了碳代谢酶的活性,抑制了AMY和INV活性,促进了SPS活性,但这种促进作用随着降雨间隔期延长变得不显著。降雨量减少促进了非结构碳水化合物(可溶性糖和淀粉)积累;(3)降雨减少降低了益智叶片全N含量,5 d处理组中W+和W-的全氮含量比10 d显著增加8.33%和9.81%,降雨量变化改变了N代谢组分;(4)同时GS/GOGAT循环活性以及NR和NiR酶活性增加,这加速了硝态氮还原和铵态氮同化。在W处理下,5 d处理组中的硝态氮和铵态氮比10 d显著降低了57.54%和152.98%。因此,在旱季降雨格局变化会影响益智的生物量、碳氮化合物及代谢酶变化,但益智通过生物量分配和叶片C、N代谢调节来适应不同的干旱胁迫,以保证益智植株的正常生长。
碳水化合物;硝酸还原;氨同化;益智;降雨格局
以橡胶为基础的农林复合系统是一项重要的管理实践,橡胶林间作复合系统的主要模式有:橡胶-茶、橡胶-甘蔗、橡胶-菠萝、橡胶-胡椒、橡胶-咖啡、橡胶-肉桂、橡胶-砂仁、橡胶-大叶千斤拔、橡胶-益智等[1-4]。其中,橡胶-益智间作模式是中国海南推广和种植面积最大的农林复合模式,也是橡胶种植户脱贫致富的有效方法,具有较好的经济效益和生态效益[5]。益智()为姜科(Zingiberaceae)山姜属()多年生常绿草本植物,主要分布在中国的海南、广东、广西地区,近年来,在云南、福建亦有少量试种。益智生于林下荫湿处或栽培[6],是我国重点发展的“四大南药”之一,为海南特有的药用植物,也是《中华人民共和国药典》(2010年版)收载的品种,有着1300多年的应用历史。
在未来全球气候变化形势下,全球或局部地区的降水格局(降水量、降水强度及降水的季节分配)将发生改变,使得植物常常遭受短暂的周期性的土壤水分亏缺。土壤水分亏缺是最常见的环境胁迫因子,其限制植物的生长、光合作用、营养吸收和代谢等生理生化进程[7-9]。海南属于典型的热带季风气候,全年雨量充沛,历年平均降雨量达1607 mm,但是降雨分布极不平衡。雨/旱季明显,5—10月为湿季,11月至翌年4月为旱季,降雨主要分布在7、8、9三个月,占全年降雨量的70%以上[10]。而降雨是橡胶-益智农林复合体系土壤水分的唯一来源,其变化必将影响到林下植物生理及代谢进程。程汉亭等[11]研究发现,在旱季的3月,降雨量低造成土壤水分含量不足是益智光合作用的主要限制生态因子。
干旱环境下植物细胞渗透胁迫是主要胁迫形式之一。干旱胁迫会破坏植物细胞渗透稳定性,增加细胞吸收环境水分的难度,严重时造成细胞失水最终引起植物的死亡。许多植物通过调节细胞的渗透势以忍耐和抵御干旱胁迫。比如,植物进化出一系列合成和积累细胞内相容性溶质的能力,以渗透调节的方式来促进蛋白质和细胞结构稳定,维持细胞膨压[12]。大多数相容性溶质是碳、氮代谢化合物,如碳水化合物、氨基酸和蛋白质等[13]。因此,植物的碳、氮代谢与植物适应逆境环境能力密切相关。有研究发现,干旱胁迫能够提高植物多种碳代谢关键酶的活性[14],促进细胞内可溶性糖的积累,以增强植物细胞的渗透调节能力。例如,干旱环境下,植物碳代谢进程中的蔗糖合酶(SS)和蔗糖转化酶(INV)活性通常会增加,进一步促进可溶性糖(葡萄糖和果糖)合成和积累[15]。同时干旱环境下,植物体内氨基酸含量会增加,特别是脯氨酸含量,但是叶片氮含量减少[16]。
本研究旨在研究降雨格局如何影响益智的生长和光合碳氮代谢。因此,本研究调查了植物生长、水分状况、关键化合物的水平以及参与C和N代谢的酶活性。对提高益智的水分管理效率和农林复合系统可持续发展具有重要意义。
1 材料与方法
1.1 材料
实验选择在海南省儋州市宝岛新村农业农村部儋州农业环境科学观测实验站进行。所选的实验样地土壤为砖红壤,橡胶树树龄13 a,平均胸径18.3 cm,树冠平均高度约11 m,冠幅平均7 m,荫蔽度80%~85%,株行距3 m×6 m。前期研究发现,橡胶林下土壤含水量与降雨量变化趋势一致,1—3月土壤含水量最低,分别为11.98%、11.97%、12.00%,此时土壤属于轻度干旱状态;6—9月土壤潮湿,其含水量均高于16%,其中8月土壤含水量达到最高,为18.32%。
通过前期对橡胶-益智农林复合模式不同月份降雨量和土壤含水量(0~20 cm)观测发现,橡胶林下土壤水分在1—3月低于12%,处于轻度干旱状态。1985—2015年气象数据表明,海南儋州市历年1—3月月均降雨量为25 mm。其中属于干旱年份的1—3月月均降雨量仅为10 mm,而模拟增雨量参照偏湿润年份1—3月月均降雨量为40 mm,同时降雨频次也有减少趋势。因此,本试验设置3个降雨梯度(模拟减雨/旱季模拟自然降雨/模拟增雨)和2个模拟降雨间隔期的双因子实验。试验采用完全随机区组设计,降雨梯度包括:模拟增雨(40 mm/月,W+)、模拟自然降雨(25 mm/月,W)和模拟减雨(10 mm/月,W‒)。模拟降雨间隔期包括:5 d/次,每次降雨量为模拟月降雨量的1/6;10 d/次,每次降雨量为模拟月降雨量的1/3;每次降雨选择在18:00—19:00进行。样方设置24 m´2 m,用土工膜包围,向下深度30 cm,避免外界土壤对水分和养分的干扰,样方顶部2 m处用透明塑料膜遮挡,消除自然降雨的干扰。同时定期清除小区内自然萌发的其他植物。
试验共设6个处理,每个处理5次重复。于2018年11月选取益智株高、分蘖数均一致,苗龄1 a的益智幼苗提前一月定值。定值40 d后(2018年12月上旬),控制土壤相对含水量至40%左右。此时进行模拟降雨处理,同时监测每个处理样地的土壤相对含水量。3月下旬取样进行指标测定。
1.2 方法
1.2.1 生长指标测定 连根部整株采取益智样品植株,洗净后置于烘箱内105℃杀青15 min,80℃恒温48 h,分别测定其地上部干质量和地下部干质量,并计算根冠比。
1.2.2 叶片相对含水量测定 叶片相对含水量= [(鲜重-干重)/(吸水饱和的叶片质量-干重)]× 100%。吸水饱和的叶片是指将新鲜叶片放入去离子水中浸泡12 h的叶片。
1.2.3 益智叶片碳代谢产物和相关酶活性测定 碳代谢碳水化合物测定(包括可溶性糖、蔗糖、非结构碳水化合物):可溶性糖采用蒽酮法测定;淀粉采用蒽酮硫酸法测定。非结构性碳水化合物在此定义为可溶性糖和淀粉的总和。
碳代谢相关酶测定:在上午9:00—10:00时,取益智功能叶片(去叶脉)剪碎,迅速用液氮速冻,在‒30℃保存测定酶活性(下同)。蔗糖磷酸合成酶(sucrose phosphate synthase, SPS)活性采用果糖和UDPG比色法;淀粉酶(amylase, AMY)和蔗糖转化酶(sucrose invertase, INV)采用比色法。酶活力以单位酶蛋白中的酶在单位时间内催化底物合成蔗糖的微摩尔数表示,单位μmol/(h·mg)。
1.2.4 益智叶片氮代谢产物和相关酶活性测定 氮形态、可溶性蛋白和游离氨基酸含量测定:分别取冷冻的叶片0.2 g,剪碎,沸水提取30 min,冷却后过滤,过滤液用于测定NO3‒-N和NO2‒-N含量,测定方法采用水杨酸法;叶片NH4+-N含量参照汤章城《现代植物生理学实验指南》测定[17];全氮采用凯氏定氮法测定。可溶性蛋白质含量采用考马斯亮蓝G-250法测定。游离氨基酸和脯氨酸分别用2 mL 10%的乙酸和5 mL 3%的磺基水杨酸提取,采用茚三酮法测定。
氮代谢相关酶测定:采用离体法测定硝酸还原酶(nitrate reductase, NR)活性,酶活力单位μmol/(h·mg);亚硝酸还原酶(nitrite reductase, NiR)采用微量扩散比色法测定;采用可见分光光度法测定谷氨酰胺合成酶(glutamine synthetase, GS),酶活力单位μmol/(h·mg);采用NADH微量速率法的方法测定谷氨酸合酶(glutamate synthase GOGAT),酶活单位μmol/(h·mg),以上酶均使用试剂盒(北京Solarbio科技有限公司)测定。
1.3 数据处理
采用SPSS统计分析软件(SPSS 16.0 for Windows, Chicago, USA)进行数据统计分析,采用单因素方差法(LSD法)和Duncan’s新复极差法作多重分析比较;以降水量和降水间隔为因子,对益智叶片的碳、氮代谢物含量和代谢酶活性采用双因子方差分析(S-N-K法),并比较其交互作用。所有数据进行正态分布(K-S法)和方差齐性检验(Levene test法),若不满足正态分布需进行数据转化(如对数转化)然后再进行方差分析。
2 结果与分析
2.1 降雨格局对益智生长的影响
随着降水量减少,益智的地上生物量和总生物量呈现逐渐下降的趋势(表1),5 d处理组中地上生物量显著高于10 d,与10 d相比,5 d中W+、W、W-处理组分别显著增加63.16%、66.66%和63.05%。与地上生物量相同,5 d中W+、W、W-处理组的总生物量比10 d分别增加60.23%、60.82%和50.68%。在5 d处理组中,益智的地下生物量随着降雨量的减少而降低,但在10 d处理组中这种趋势并不显著,5 d处理下W+和W增加53.37%和48.46%。益智的根/冠比随着降水量的减少而增加,5 d处理组中的根/冠比显著低于10 d。益智叶片的相对含水量随着降水量的减少而降低,与5 d相比,在10 d处理组中W-的叶片相对含水量显著增加3.63%。降水量和降水间隔的相互作用影响了根系生物量、根冠比、总生物量和叶片相对含水量。
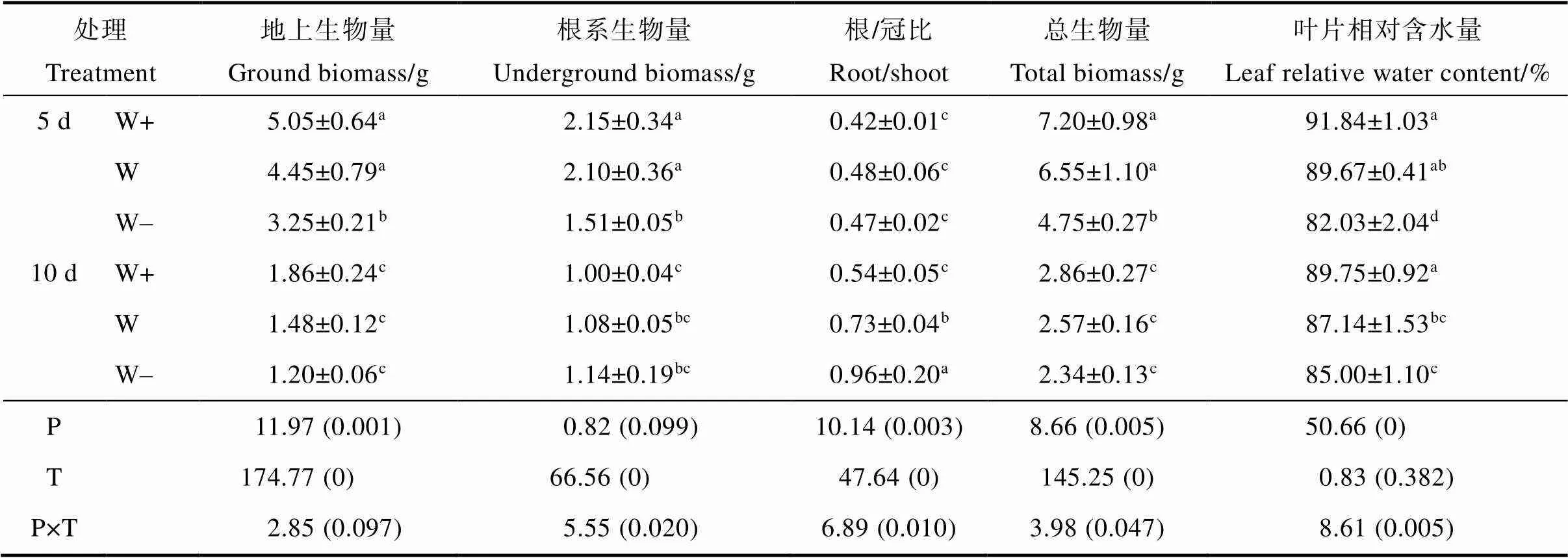
表1 降水模式变化对益智生长状况的影响及其交互作用
注:P表示模拟降雨量;T表示模拟降雨间隔天数;P×T表示降雨量与间隔天数的交互作用。不同小写字母表示处理间差异显著(<0.05)。
Note: P means simulated precipitation; T means the interval days of simulated precipitation; P×T means the interaction of P and T. Different lowercase letters indicate significant difference among treatments (<0.05).
2.2 降雨格局对益智叶片碳代谢和相关酶的影响
叶片的可溶性糖含量、非结构碳水化合物和可溶性糖/淀粉随着降水量减少逐渐增加(表2)。叶片的可溶性糖含量和非结构碳水化合物不受降水间隔的显著影响。10 d和5 d相比,10 d处理组中W+的可溶性糖/淀粉增加了73.74%,W-降低了17.11%。淀粉含量则是随着降水量的减少而降低,但在10 d处理组中该结果并不显著。10 d比5 d处理组的W+的淀粉含量降低了38.03%。降水量和降水间隔的相互作用影响植物淀粉含量和可溶性糖/淀粉。
植物叶片中的AMY和INV酶活性随着降水量的减少而显著降低(图1)。在5 d和10 d处理组中,W+的AMY和INV酶的活性最高。益智叶片的AMY和INV酶活性不受降水间隔的显著影响。5 d处理下,叶片的SPS酶活性随着降水量的减少而增加,但在10 d处理组中这种趋势并不显著(图1)。5 d处理组下的SPS酶活性显著低于10 d,W+、W、W-分别增加了122.20%、63.13%、2.38%。降水量和降水间隔的相互作用影响SPS酶活性。
2.3 降雨格局对益智叶片硝酸还原和铵生产的影响
益智叶片的硝态氮、铵态氮和亚硝酸盐含量均是随着降水量的减少而增加(表3)。5 d处理组的硝态氮含量均低于10 d处理组,5 d处理中的W的硝态氮含量比10 d显著降低了57.54%。与5 d相比,10 d处理组中W的铵态氮含量增加152.98%。亚硝酸盐含量不受降水间隔的显著影响。益智叶片的NiR和NR酶活性随降水量减少而增加(表3),降水量和降水间隔的相互作用对叶片的NiR酶和NR酶活性影响不显著。
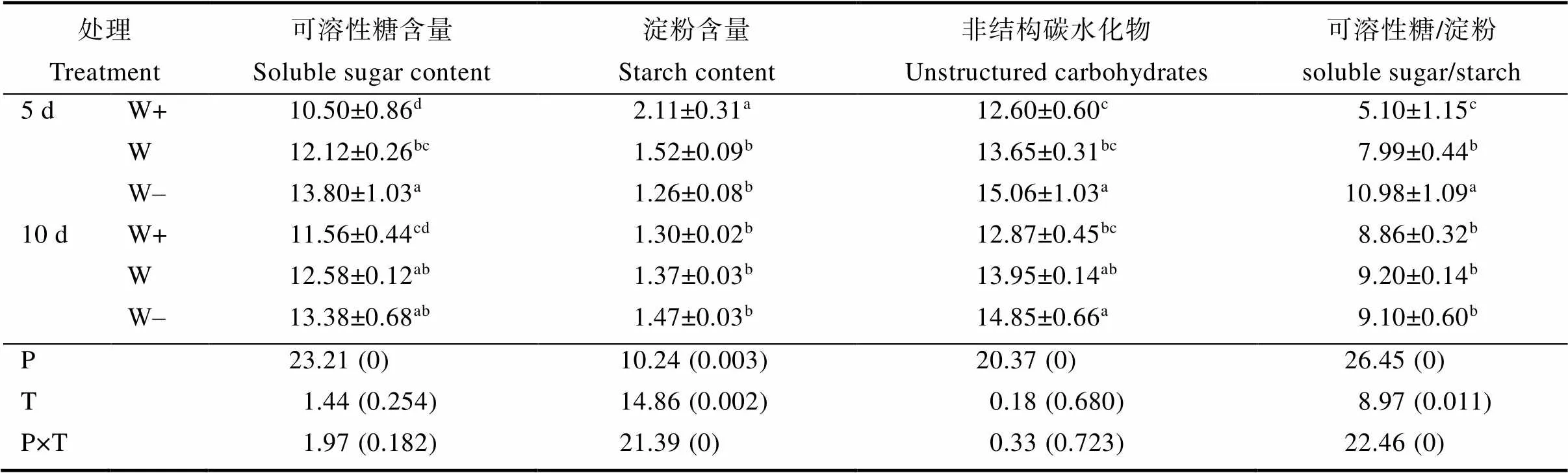
表2 降水模式的变化对益智叶片碳水化合物含量的影响及其交互作用
注:同列数据后不同小写字母表示处理间差异显著(<0.05)。
Note: Different lowercase letters after the same column of data indicate significant difference (<0.05).
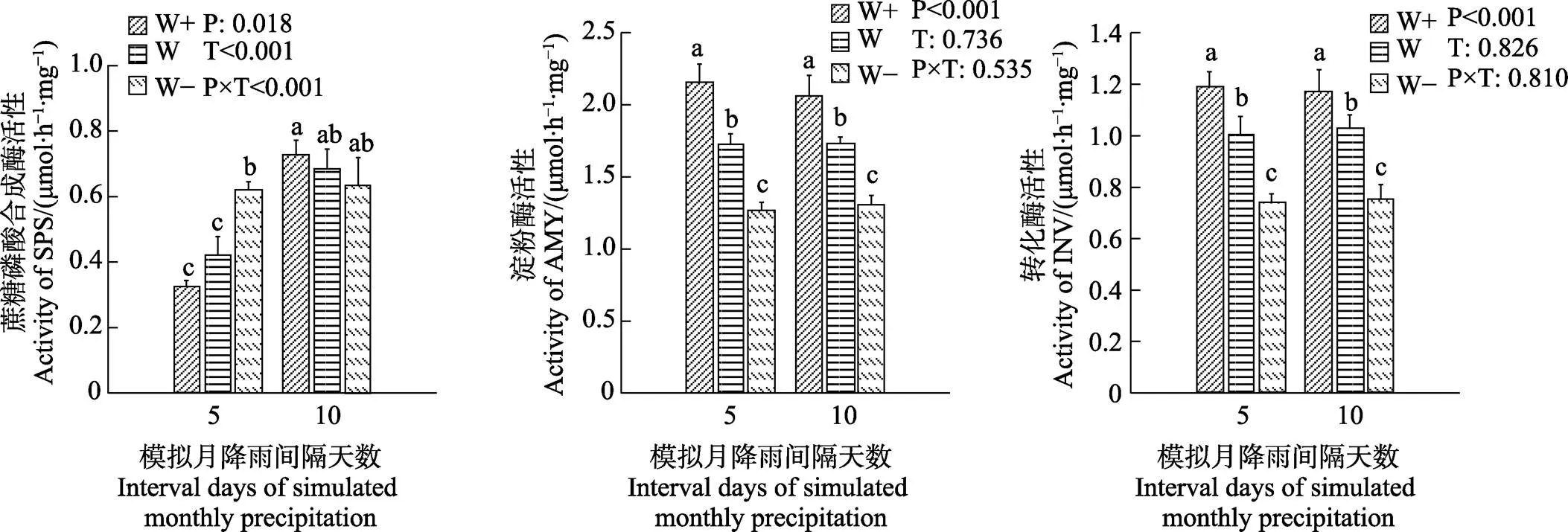
不同小写字母表示处理间差异显著(P<0.05)。
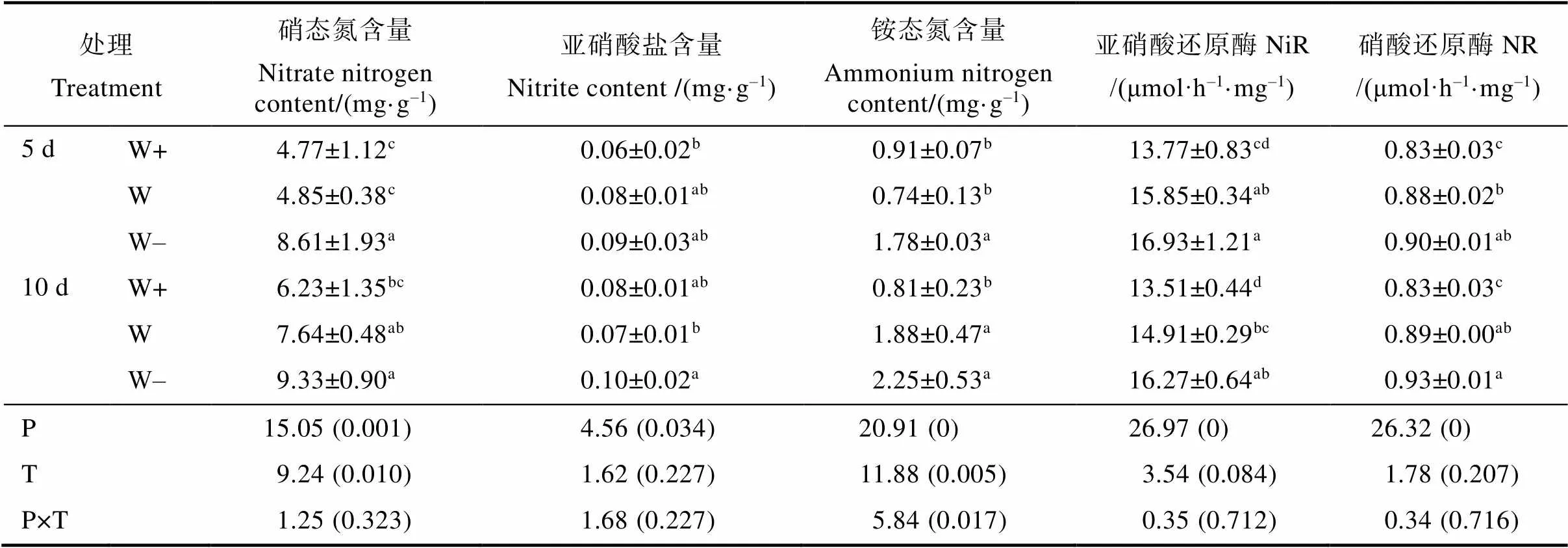
表3 降水模式的变化对益智叶片铵氮和硝氮含量的影响及其交互作用
注:同列数据后不同小写字母表示处理间差异显著(<0.05)。
Note: Different lowercase letters after the same column of data indicate significant difference (<0.05).
2.4 降雨格局对益智叶片铵与同化产物结合的影响
益智叶片的游离氨基酸、脯氨酸含量随着降雨量的减少而增加(表4)。与5 d相比,10 d处理组中W的游离氨基酸含量降低20.31%,而植物叶片的脯氨酸含量不受降水间隔的影响。叶片的蛋白质含量随着降雨量的减少呈现先减后增的趋势,在W-达到最大值,但降水间隔期对其影响不显著。叶片的全氮含量随着降水量的减少而降低(表4),与5 d相比,10 d处理组中W+和W-的全氮含量分别增加8.33%和9.81%。降水量和降水间隔的相互作用影响了植物游离氨基酸和全氮含量。
益智叶片的GOGAT和GS酶活性随着降水量的减少而增加(图2)。在不同降水间隔下GOGAT酶活性差异不显著。与5 d相比,10 d处理组中的W+和W的GS酶活性分别增加8.67%和29.16%。降水量和降水间隔的相互作用影响GS酶的活性。
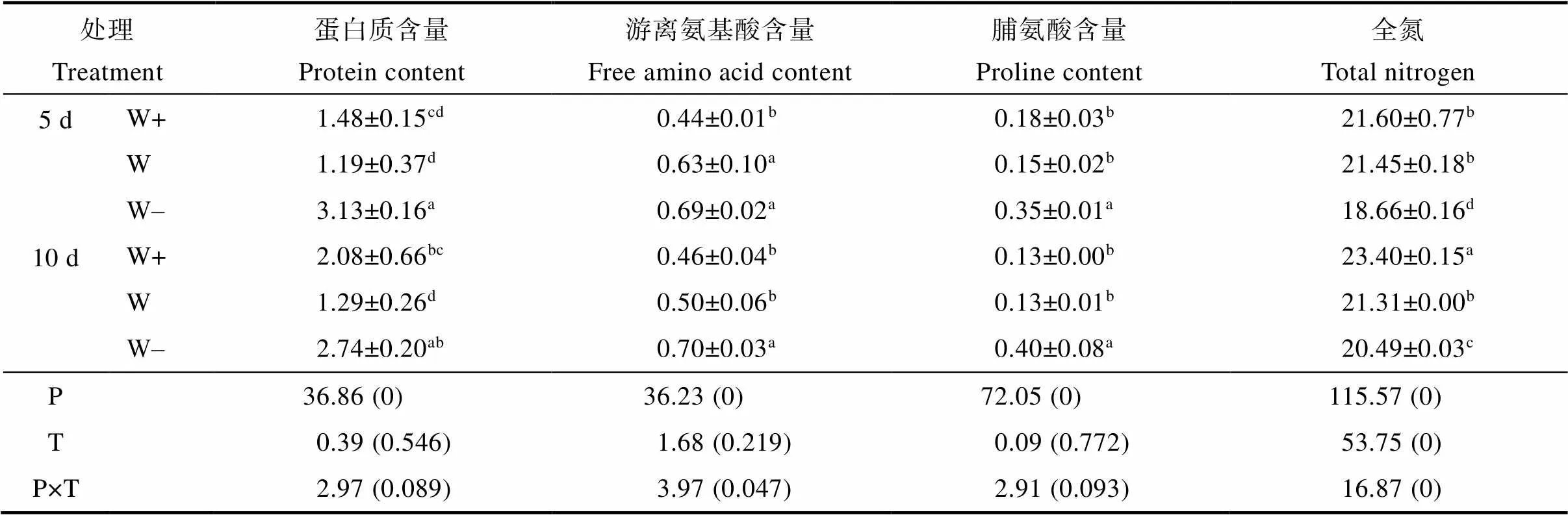
表4 降水模式的变化对益智叶片含氮化合物含量的影响及其交互作用
注:同列数据后不同小写字母表示处理间差异显著(<0.05)。
Note: Different lowercase letters after the same column of data indicate significant difference (<0.05).
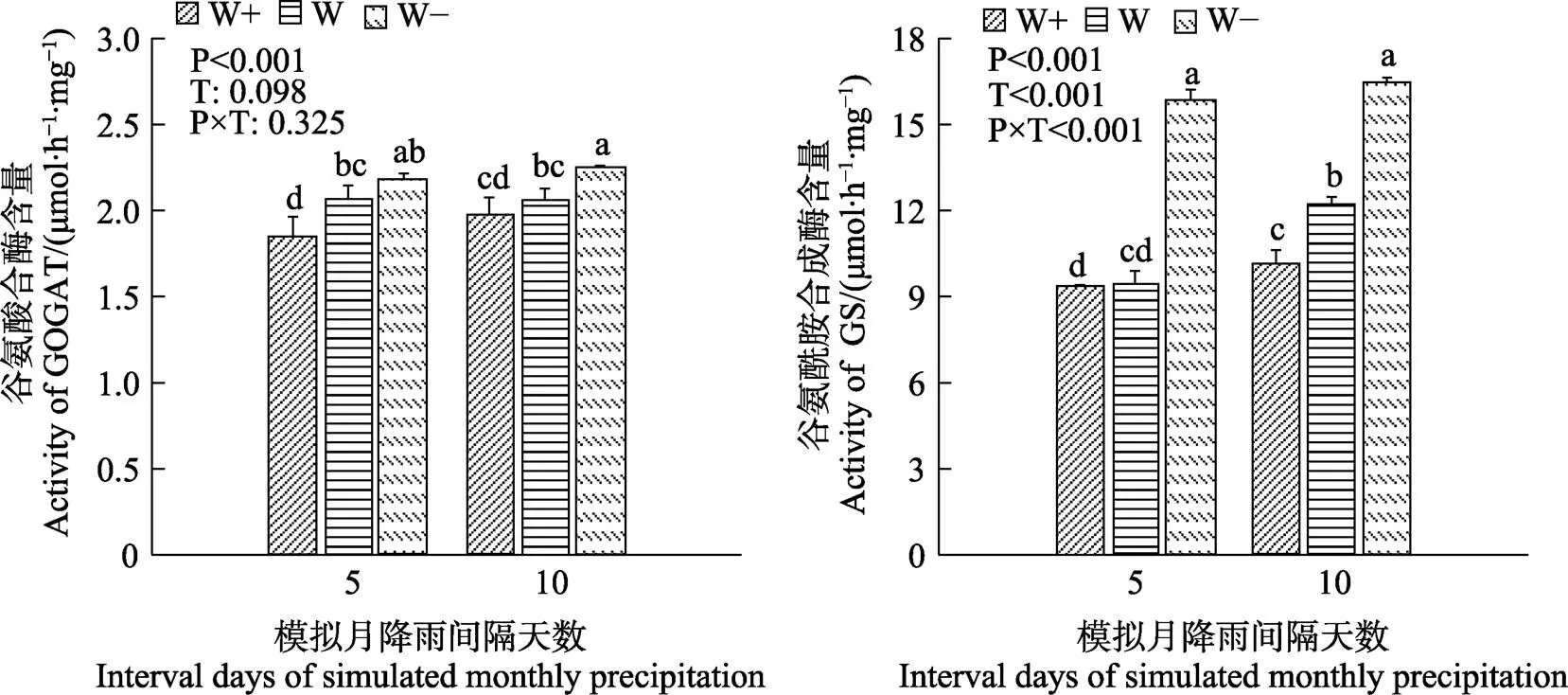
不同小写字母表示处理间差异显著(P<0.05)。
3 讨论
3.1 降雨格局变化对益智生长的影响
由气候变化引起的干旱的频率和强度日益增加,已成为本世纪的一个全球问题[18]。干旱胁迫通常导致植物地上部分的大量减少[19]。本研究中,在5 d的降雨间隔处理下,降雨量减少显著抑制了益智地上部分的生长。同时间隔期延长进一步加剧了这一现象,这与YANG等[20]的研究结果一致。
植物的地下部分也会受到干旱胁迫的影响。在5 d的降雨间隔处理下,随着降雨量减少,益智根系生物量和总生物量显著减少。该研究结果与YIN等[21]的研究发现康定杨的根干物质显著减少一致。但也有研究表明,植物根系生长受到干旱胁迫的影响程度可能与其胁迫强度有关[19, 22]。本研究结果发现,降雨间隔期为10 d的地下生物量均低于5 d,表明降雨间隔期的延长使植物根系受到的干旱胁迫影响更大,但降雨量减少对根的生物量影响并不显著。而根/冠比的增加,作为一种对干旱的适应性状,因为根系生长通常比空中生长更不容易受干旱的影响[23]。这种形态策略有助于植物适应干旱环境,其主要依赖于整个植物叶片C和N化合物组分及代谢[24]。
3.2 降雨格局变化对益智叶片碳代谢的影响
高等植物中,叶绿体中淀粉作为C的储存形式,以支持新陈代谢[25]。干旱条件下淀粉含量的变化取决于光合效率和一些参与淀粉降解的酶。本研究结果表明,降雨减少时益智叶片淀粉含量显著降低,同时淀粉酶活性受到抑制。这与ZEID等[23]的研究结果一致,其发现在中度或重度干旱胁迫下紫花苜蓿叶片淀粉酶活性会下降。表明干旱胁迫可能会降低叶片光合产物的合成。可溶性糖的积累是对干旱胁迫的一种适应性响应[26],并与蔗糖磷酸合成酶和转化酶活性密切相关。本研究结果显示,降雨减少促进了益智叶片可溶性糖的积累,伴随着蔗糖磷酸合成酶活性增加和转化酶活性降低,即可溶性糖的增加可以提高植物叶片渗透压以适应干旱环境。此外,干旱胁迫下紫花苜蓿也发现同样的结果[23]。以上研究表明,非结构碳水化合物(可溶性糖和淀粉)的分配可以调节水势,是益智光合器官对旱季降雨减少的一种响应。
3.3 降雨格局变化对益智叶片氮代谢的影响
氮代谢调控是植物抗旱性的关键,而干旱和氮营养之间是一个非常复杂的网络,几乎影响植物的所有生理过程[27]。本研究结果表明,在W-处理下益智叶片中NO3‒的浓度和NR活性均有显著的提高。同时,降雨间隔期的延长使得NO3‒的浓度略有增加。这主要与NO3‒的供给和从液泡到细胞质的转移率有关,有研究表明,较高的NO3‒和NR活性可促进植物在水分亏缺下正常生长[24]。与此同时,NO2‒浓度在降雨减少时普遍增加,并激发了NiR酶活性,表明益智叶片中NiR的表达程度与NO2‒水平密切相关。NH4+不仅由NO3‒的还原产生,更重要的是,在水分胁迫后激活的光呼吸过程中,甘氨酸的氧化也会产生NH4+[28]。旱季降雨减少增加了益智叶片NH4+的浓度,这与SANCHEZ-RODRIGUEZ等[22]的结果一致。然而,也有研究表明,干旱胁迫对NO3‒的还原和NH4+产生无显著影响[9],这说明,不同物种可能具有不同的NH4+代谢机制。
过量的NH4+对植物细胞有害,因此它必须通过GS/GOGAT循环迅速吸收[29]。本研究结果显示,降雨减少不同程度地增加了GS/GOGAT循环活性,特别是在W-处理下,这是因为较高的NH4+浓度诱导的结果。这与SANCHEZ等[24]发现番茄的GS/GOGAT循环活性增加而表现出显著抗旱性得到的结果一致。因此,干旱胁迫激发了GS和GOGAT酶活性,加速了益智叶片NH4+的同化,一定程度上减轻了NH4+对益智叶片细胞的毒害。叶片氮含量的比例对估算CO2同化能力至关重要,而干旱会降低植物的氮利用率,降低叶片氮含量[30]。在本研究中,随着降雨减少,益智叶片的总氮减少,这是因为干旱促进了益智叶片蛋白质的水解。还原氮主要由氨基酸和蛋白质形成,可溶性蛋白和游离氨基酸还含有若干参与氮代谢和重要渗透物质的转移化合物[16]。本研究还发现,降雨减少导致益智叶片可溶性蛋白、游离氨基酸和脯氨酸大量积累,说明干旱在一定程度上加强了N传递功能和细胞的渗透调节。
4 结论
综上所述,旱季降雨减少会影响益智的生长,降低益智地上和地下生物量;由于关键酶的平衡调节,非结构碳水化合物(可溶性糖和淀粉)含量在降雨减少时增加,以调节叶片细胞水势应对逆境胁迫;同时,干旱胁迫加速了益智叶片的N代谢,减轻了NH4+对益智叶片细胞的毒害,增强了细胞的渗透调节。益智植株生物量分配和光合器官生理策略将保证益智植株适应旱季降雨格局的变化,实现正常生长和光合作用。
[1] PENOT E, OLLIVIER I. Rubber tree intercropping with food-crops, perennial, fruit and tree crops: everal examples in Asia, Africa and America[J]. Bois Et Forets Des Tropiques, 2009, 15(301): 67-82.
[2] 程汉亭, 李勤奋, 刘景坤, 严廷良, 张俏燕, 王进闯. 蚯蚓粪-土复合基质对益智幼苗生长和叶绿素荧光特征的影响[J]. 热带作物学报, 2017, 38(12): 2256-2262.
CHENG H T, LI Q F, LIU J K, YAN T L, ZHANG Q Y, WANG J C. Effects of vermicompost-soil complex media on growth and chlorophyll fluorescence characters ofMiq seedlings[J]. Chinese Journal of Tropical Crops, 2017, 38(12): 2256-2262. (in Chinese)
[3] 程汉亭, 沈奕德, 范志伟, 黄乔乔, 李晓霞, 黄东东. 橡胶‒益智复合生态系统综合评价研究[J]. 热带农业科学, 2014, 34(10): 7-11.
CHENG H T, SHEN Y D, FAN Z W, HUANG Q Q, LI X X, HUANG D D. Comprehensive evaluation of the rubber-agro-forestry ecosystem[J]. Chinese Journal of Tropical Agriculture, 2014, 34(10): 7-11. (in Chinese)
[4] 庞家平, 陈明勇, 唐建维, 郭贤明, 曾 荣. 橡胶-大叶千斤拔复合生态系统中的植物生长与土壤水分养分动态[J]. 山地学报, 2009, 27(4): 433-441.
PANG J P, CHEN M Y, TANG J W, GUO X M, ZENG R. The dynamics of plant growth and soil moisture and nutrient in the rubber plantation and rubber-agro-forestry system in Xishuangbanna, southwest China[J]. Journal of Mountain Science, 2009, 27(4): 433-441. (in Chinese)
[5] 程汉亭, 王进闯, 侯宪文, 李勤奋, 邹雨坤, 李光义, 王定美. 胶价低迷背景下昌江县民营橡胶产业的发展现状和对策[J]. 热带农业科学, 2015, 35(5): 78-81.
CHENG H T, WANG J C, HOU X W, LI Q F, ZOU Y K, LI G Y, WANG D M. Development status of the private rubber industry in Changjiang under a situation of rubber price downturn[J]. Chinese Journal of Tropical Agriculture, 2015, 35(5): 78-81. (in Chinese)
[6] 程汉亭, 刘景坤, 严廷良, 张俏燕, 王进闯. 不同采收期对药用植物-益智种子质量的影响研究[J]. 热带作物学报, 2017, 38(10): 1840-1845.
CHENG H T, LIU J K, YAN T L, ZHANG Q Y, WANG J C. Effects of different picking on seed quality of medicinal plantsMiq[J]. Chinese Journal of Tropical Crops, 2017, 38(10): 1840-1845. (in Chinese)
[7] ROBRED A, PEREZ L U, MIRANDA A, J, LACUESTA M., MENA P A, MUNOZ R A. Elevated CO2reduces the drought effect on nitrogen metabolism in barley plants during drought and subsequent recovery[J]. Environmental and Experimental Botany, 2011, 71(3): 399-408.
[8] YI X P, ZHANG Y L, YAO H S, LUO H H, GOU L, CHOW W S, ZHANGW F. Different strategies of acclimation of photosynthesis, electron transport and antioxidative activity in leaves of two cotton species to water deficit[J]. Functional Plant Biology, 2016, 43(5): 448-460.
[9] LIU C G, WANG Y J, PAN K W, ZHU T, LI W, ZHANG L. Carbon and nitrogen metabolism in leaves and roots of dwarf bamboo (Yi) subjected to drought for two consecutive years during sprouting period[J]. Journal of Plant Growth Regulation, 2014, 33(2): 243-255.
[10] 吴志祥, 谢贵水, 杨 川, 周兆德, 王 旭. 海南岛西部不同林龄橡胶林土壤养分特征[J]. 海南大学学报, 2011, 29(1): 42-48.
WU Z X, XIE G S, YANG C, ZHOU Z D, WANG X. Soil nutrient characteristics of rubber plantations at different age stages in western area of Hainan island[J]. Natural Science Journal of Hainan University, 2011, 29(1): 42-48. (in Chinese)
[11] 程汉亭, 李勤奋, 刘景坤, 严廷良, 张俏燕, 王进闯. 橡胶林下益智光合特性的季节动态变化[J]. 植物生态学报, 2018, 42(5): 585-594.
CHENG H T, LI Q F, LIU J K, YAN T L, ZHANG Q Y, WANG J C. Seasonal changes of photosynthetic characteristics ofgrowing under[J]. Chinese Journal of Plant Ecology, 2018, 42(5): 585-594. (in Chinese)
[12] UPADYHYAYA H, SAHOO L, PANDA S K. Molecular physiology of osmotic stress in plants[J]. Molecular Stress Physiology of Plants, 2013: 179-192.
[13] WANG H, ZHANG M, GUO R, SHI D, LIU B, LIN X, YANG C. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (L.)[J]. BMC Plant Biology, 2012, 12(1): 1-11.
[14] YANG J C, ZHANG J H, WANG Z Q, ZHU Q. Activities of starch hydrolytic enzymes and sucrose-phosphate synthase in the stems of rice subjected to water stress during grain filling[J]. Journal of Experimental Botany, 2011, 52(364): 2169-2179.
[15] PARVAIZ A, SATYAWATI S. Salt stress and phyto-biochemical responses of plants-a review[J]. Plant Soil and Environment. 2008, 54(3): 89-99.
[16] GUPTA N, THIND S K, BAINS N S. Glycine betaine application modifies biochemical attributes of osmotic adjustment in drought stressed wheat[J]. Plant Growth Regulation.2014, 72(3): 221-228.
[17] 汤章城. 现代植物生理学实验指南[M]. 北京: 科学出版社, 1999.
TANG Z C. Guide to modern plant physiology experiment[M]. Beijing: China Science Publishing, 1999.
[18] MCDDOWELL N G. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality[J]. Plant Physiology, 2011, 155(3): 1051-1059.
[19] INCLAN R, GIMENO B S, DIZENGREMEL P, SANCHEZ M. Compensation processes of Aleppo pine (Mill.) to ozone exposure and drought stress[J]. Environmental Pollution, 2005, 137(3): 517-524.
[20] YANG F, MIAO L F. Adaptive responses to progressive drought stress in two poplar species originating from different altitudes[J]. Silva Fennica, 2010, 44(1): 23-37.
[21] YIN C, PENG Y, ZANG R, ZHU Y, LI C. Adaptive responses ofto drought stress[J]. Physiologia Plantarum, 2005, 123(4): 445-451.
[22] ERICE G, LOUAHLIA S, IRIGOYEN J J, SANCHEZ D M, AVICE J C. Biomass partitioning, morphology and water status of four alfalfa genotypes submitted to progressive drought and subsequent recovery[J]. Journal of Plant Physiology, 2010, 167(2): 114-120.
[23] ZEID I M, SHEDEED Z A. Response of alfalfa to putrescine treatment under drought stress[J]. Biologia Plantarum, 2006, 50(4): 635-640.
[24] SANCHEZ R E, RUBIO W M, RIOS J J, BLASCO B, ROSALES M A, MELGAREJO R, ROMERO L, RUIZ J M. Ammonia production and assimilation: its importance as a tolerance mechanism during moderate water deficit in tomato plants[J]. Journal of Plant Physiology, 2011, 168(8): 816-823.
[25] ZEEMAN S C, SMITH S M, SMITH A M. The breakdown of starch in leaves[J]. New Phytologist, 2004, 163(2): 247-261.
[26] SICHER R C, TIMLIN D, BAILEY B. Responses of growth and primary metabolism of water-stressed barley roots to rehydration[J]. Journal of Plant Physiology, 2012, 169(7): 686-695.
[27] LAWLOR D W. Carbon and nitrogen assimilation in relation to yield: mechanisms are the key to understanding production systems[J]. Journal of Experimental Botany, 2002, 53(370): 773-787.
[28] WANG H, WU Z, HAN J, YANG C. Comparison of ion balance and nitrogen metabolism in old and young leaves of alkali-stressed rice plants[J]. PLoS One, 2012, 7(5): 37817-37827.
[29] THOMAS F M, HILKER C. Nitrate reduction in leaves and roots of young pedunculate oaks () growing on different nitrate concentrations[J]. Environmental and Experimental Botany, 2000, 43(1): 19-32.
[30] SINCLAIR T R, PINTER P J, KIMBALL B A, ADAMSEN F J, LAMOTYE R L, WALL G W, HUNSAKER D J, ADAM N, BROOKS T J, GARCIA R L, THOMPSON T, LEAVITT S, MATTHIAS A. Leaf nitrogen concentration of wheat subjected to elevated [CO2] and either water or N deficits[J]. Agriculture, Ecosystems and Environment, 2000, 79(1): 53-60.
Effects of Precipitation Pattern Change on the Growth Carbon and Nitrogen Metabolism ofDuring Dry Season
HU Wen1,2,3, ZHOU Xiaohui2,3, LI Qinfen2,3, CHENG Hanting2,3*
1. College of Ecology and Environment, Hainan University, Haikou, Hainan 570228, China; 2. Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences / Hainan Key Laboratory of Tropical Eco-Circular Agriculture, Haikou, Hainan 571101, China; 3. Key Laboratory of Low Carbon Green Agriculture in Tropical China, Ministry of Agriculture and Rural Affairs, Haikou, Hainan 571101, China
is one of the four southern medicines in China, and its cultivation under rubber forest has become the most important agroforestry mode in rubber plantation. The study previously found that insufficient precipitation is the main limiting factor for the growth and photosynthesis of.during the dry season. However, our knowledge is still limited in understanding the response ofto precipitation pattern change during the dry season. Three precipitation gradients including decreased precipitation (W-)10 mm/ month, natural precipitation (W) 25 mm/ month and increased precipitation (W+) 40 mm/ month, and set 5 days (5 d) and 10 days (10 d) of interval between precipitation were simulated in the study. The leaf water content in W- plant of 10 d was significantly increased by 3.63% compared with 5 d. In addition, we analyzed plant growth parameters, carbohydrate and N compound levels and the activity of key enzymes related to C and N metabolism through two factors experiment of precipitation and precipitation interval. The aboveground biomass and underground biomass of.decreased with the decrease of precipitation. The total biomass of 5 d was significantly higher than that of 10 d at all precipitation levels (<0.05). The plant growth was further inhibited by precipitation and extended precipitation interval. The decreased precipitation changed the activity of carbon metabolism enzymes, inhibited the AMY and INV activity, and promoted the SPS activity. However, this promoting effect became trivial with extended precipitation interval. The accumulation of non-structural carbohydrates (soluble sugar and starch) was promoted by decreased precipitation and activity change of C metabolic enzymes. The decreased precipitation lowered the total N content of.leaves, and the total N content of W+ and W- in 5 d was significantly increased by 8.33% and 9.81% compared with that of 10 d. The N metabolic components changed in plant leaves with decreased precipitation. Meanwhile, the GS/GOGAT, NR and NiR activity increased with the decreased precipitation, which accelerated nitrate nitrogen (NO3‒) reduction and ammonium (NH4+) assimilation. The NO3‒and NH4+in W plant in 5 d were significantly decreased by 57.54% and 152.98 compared with that of 10 d. Therefore, the change of precipitation pattern could affect the change of biomass carbon and nitrogen compounds and metabolic enzymes of the plant during the dry season. In summary, drought stress was adapted to ensure normal growth ofthrough biomass allocation and C and N metabolism regulation.
carbohydrate; ammonium assimilation; ammonium assimilation;; precipitation patterns
S567.5
A
10.3969/j.issn.1000-2561.2022.12.022
2022-02-16;
2022-04-25
海南省基础与应用基础研究计划(自然科学领域)高层次人才项目(No. 2019RC281);国家自然科学基金项目(No. 31800535)。
胡 雯(1997—),女,硕士研究生,研究方向:资源利用与植物保护。*通信作者(Corresponding author):程汉亭(CHENG Hanting),E-mail:chenghanting@163.com。
