苹果树腐烂病菌丝氨酸蛋白酶基因VM1G_08720的功能分析
2023-01-12刁雨菲熊雄郑金柱靳纪洋于成明刘会香
刁雨菲,熊雄,郑金柱,靳纪洋,于成明*,刘会香*
苹果树腐烂病菌丝氨酸蛋白酶基因的功能分析
刁雨菲1,熊雄2,郑金柱3,靳纪洋2,于成明1*,刘会香1*
1. 山东农业大学植物保护学院/山东省林业有害生物防控工程技术研究中心, 山东 泰安 271018 山东农业大学林学院, 山东 泰安 271018 徂徕山林场, 山东 泰安 271027
苹果是我国种植面积最广的温带水果之一,由苹果黑腐皮壳菌()引起的苹果树腐烂病(canker of apple)是危害我国苹果树最为严重的病害之一。明确病原菌生长发育、响应环境胁迫及致病作用,解析病原菌的适应性及致病分子机制,有利于提出病害新的防控措施。本研究应用Double-joint PCR以及gap-repair方法获得基因的缺失突变体和回补菌株。利用十字交叉法和伤口接种等方法,研究了基因对病菌生长发育和致病性的影响。通过在PDA培养基中分别添加400 μg/mL的荧光增白剂、200 μg/mL的刚果红、0.01%的SDS三种细胞壁干扰剂和0.5 M的NaCl、0.5 M山梨醇两种渗透胁迫因子,研究了该基因对病菌细胞壁完整性的维持及渗透胁迫的影响。通过设置不同pH的培养基,研究了基因在不同pH下对病菌生长发育的影响。研究结果显示,与野生型sdau11-175相比,基因缺失突变体在菌落颜色、菌丝致密度、生长速率无显著差异,分生孢子器产生数量减少;在含有200 μg/mL刚果红、0.01%SDS的培养基上,抑制率显著降低,在含有0.5 M NaCl的培养基上的抑制率显著增加,而在含有400 μg/mL荧光增白剂和0.5 M山梨醇的培养基上,无显著差异;在pH为4-10时,生长速率显著降低,而在pH为11-12时,无显著性差异;在苹果果实和苹果枝条上的病斑大小无显著性差异,回补菌株均恢复到野生型水平。表明基因在病菌分生孢子器的产生、酸碱度的响应、维持细胞壁完整性以及渗透胁迫方面发挥重要作用。
苹果树腐烂病菌;基因; 丝氨酸蛋白酶; 表型分析
苹果是我国种植面积最广的温带水果之一,由苹果黑腐皮壳菌(Miyabe et Yamada)引起的苹果树腐烂病(canker of apple)是分布广泛、危害严重、防治困难的一种枝干性病害,该病害主要危害主干和大枝,造成苹果树皮腐烂,树势衰弱,严重时可造成整株死亡[1, 2]。对苹果腐烂病菌适应性及致病分子机制进行研究有利于提出病害新的防控措施。
丝氨酸蛋白酶(S)是蛋白酶中最庞大的家族,以丝氨酸为活性中心蛋白水解酶,丝氨酸蛋白酶在治疗心血管疾病、胚胎发育、组织重建、细胞分化、血管形成和病原侵入等过程中都发挥着重要的作用[3,4]。在植物病原真菌中,研究发现,烟曲霉()中的丝氨酸蛋白酶基因ALP2缺失后,菌丝生长速度减慢,分生孢子产生数量减少,分生孢子囊直径减少[5]。在稻瘟病菌()中,丝氨酸蛋白酶基因和在cAMP诱导的孢子萌发和附着胞形成阶段均上调表达[6],基因的缺失突变体表现为气生菌丝减少,分生孢子产生数量减少,致病性下降,而基因的缺失在生长发育和致病性上与野生型相比无显著差异[7]。苹果腐烂病菌中丝氨酸蛋白酶基因位于10号染色体上,全长1888bp,编码606个氨基酸,登录号为KUI73074.1,具有Peptidases_S8_S53超家族结构域,该基因对病菌生长发育及致病作用调控功能尚未研究报道。
本研究以基因为目的基因,利用Double-joint PCR技术构建敲除载体,通过PEG介导原生质体转化法获得敲除突变体,利用gap-repair技术获得回补菌株;通过对野生型菌株、基因缺失突变体、回补菌株的生长发育、对胁迫因子响应、致病性等方面的比较分析,研究结果为明确基因在病菌生长发育、响应环境胁迫及致病等方面功能提供理论依据。
1 材料与方法
1.1 实验材料
苹果树腐烂病菌野生型菌株sdau11-175由山东农业大学植物保护学院林木病原与寄主分子互作研究室提供。菌株保存在20%的甘油中,苹果树2年生枝条采自山东农业大学植保实验站,苹果果实购于市场上生长一致的富士苹果
1.2 基因VM1G_08720 缺失突变体及回补菌株的获得
从苹果腐烂病菌基因组数据库[8]NCBI(http://www.ncbi.nlm.nih.gov)中获得基因序列,通过替换基因开放阅读框(ORF),构建基因敲除突变体,以野生型菌株sdau11-175 DNA为模板,分别扩增基因的上游片段、HPH片段以及下游片段,所用引物见表1(其中08720-SY-R和08720-XY-F小写部分,分别与HPH-F、HPH-R两端序列反向互补)。应用Double-joint PCR方法构建敲除载体[9],利用PEG介导法将得到的敲除载体转化到sdau11-175的原生质体中[10]。用3对引物(表1)验证是否敲除成功[11]。用gap repair的方法构建回补载体,以sdau11-175为模板,用表1的回补引物扩增基因片段,后与pFL2质粒相连,将得到的回补载体转到敲除突变体的原生质体中,筛选得到回补菌株[12]。

表1 本研究所使用的引物
1.3 基因VM1G_08720 对病菌生长发育的影响
将野生型菌株sdau11-175、基因缺失突变体和回补菌株接种到PDA培养基上,25℃黑暗培养3d后测定菌落直径大小,观察菌落形状、颜色、菌丝生长状况。每个处理重复3次,试验共重复3次,数据处理采用SPSS方法(<0.05)[13]。继续将野生型菌株sdau11-175、基因缺失突变体和回补菌株置于光暗交替的培养基中培养25 d后,观察分生孢子器的产生情况,实验方法同上。
1.4 基因VM1G_08720对病菌细胞壁完整性的影响
在PDA培养基中添加了400 μg/mL的荧光增白剂、200 μg/mL的刚果红、0.01%的SDS三种细胞壁干扰剂,观察突变体对细胞壁干扰剂的敏感性,评价基因对病菌细胞壁完整性的调控作用,上述均在3 d后测定菌落直径大小,每个处理重复3次,试验共重复3次。
1.5 基因VM1G_08720对病菌渗透胁迫的影响
在PDA培养基中添加了0.5 M的NaCl、0.5 M的山梨醇两种渗透胁迫因子,测定基因对渗透胁迫的响应,上述均在3 d后测定菌落直径大小,每个处理重复3次,试验共重复3次。
1.6 基因VM1G_08720对病菌酸碱性适应的影响
将培养3 d的野生型菌株sdau11-175、基因缺失突变体、回补菌株接种到不同pH值(pH为4-12)的PDA平板上,25 ℃培养3 d,测量菌落直径。每个处理重复3次,试验重复3次。
1.7 基因VM1G_08720 对病菌致病力的影响
选取PDA培养基上培养3 d的野生型菌株sdau11-175、基因缺失突变体和回补菌株及生长一致的成熟苹果果实和2年生苹果枝条,采用伤口接种法,接种后的苹果果实和枝条置于25℃下保湿培养7 d,每个处理重复10次,试验共重复3次。
2 结果与分析
2.1 基因VM1G_08720 缺失突变体及回补菌株获得
以sdau11-175基因组DNA为模板,用引物08720-SY-F/R、08720-XY-F/R扩增得到了上游片段、下游片段;以PCB1003为模板,用引物HPH-F/R扩增得到了HPH片段,三片段融合后得到敲除载体,转入原生质体后,用3对引物进行验证,证明敲除成功(图1A)。
以sdau11-175为模板,用引物08720HB-F/R扩增得到了回补片段,转入原生质体后,用引物08720HB-F/R进行PCR验证,证明回补成功,成功获得回补菌株(图1B)。
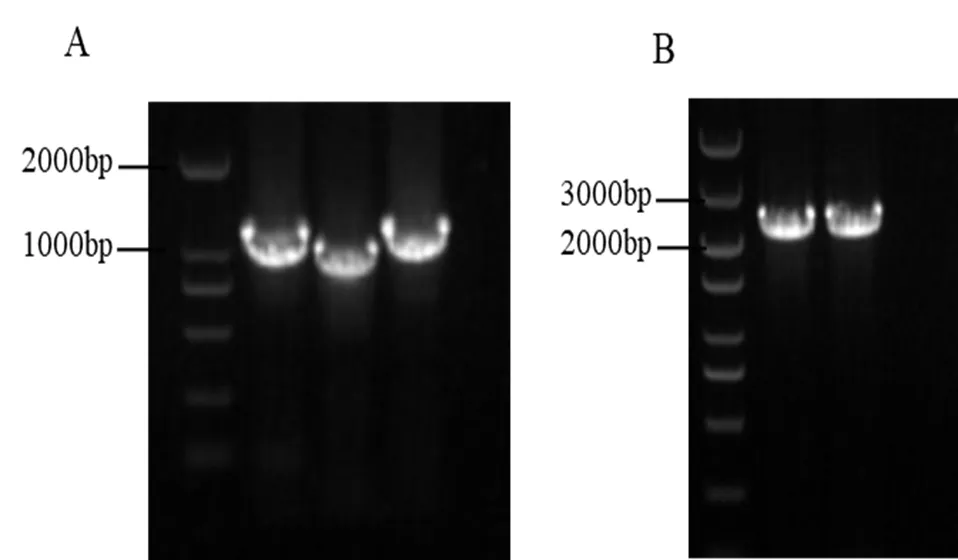
图1 苹果腐烂病菌基因VM1G_08720缺失突变体及回补菌株PCR鉴定
备注:A:3对引物对缺失突变体进行PCR验证;B:回补菌株的PCR鉴定。
Note:A:PCR identification for deletion mutants with three pairs of primers;B:PCR identification for complementation strains.
2.2 基因VM1G_08720对病菌生长发育的影响
在25 ℃生长3 d后,与野生型菌株sdau11-175相比,基因缺失突变体在菌落颜色、菌丝致密度、生长速率无显著差异(图2A、2B)。继续将菌株光暗交替培养25 d后,观察分生孢子器产生情况,与野生型菌株sdau11-175相比,基因缺失突变体分生孢子器产生数量减少(图2C),野生型菌株分生孢子器平均产生数量为120个,而基因缺失突变体分生孢子器平均产生数量为83个,回补菌株分生孢子器产生数量恢复到野生型水平(图2D)。
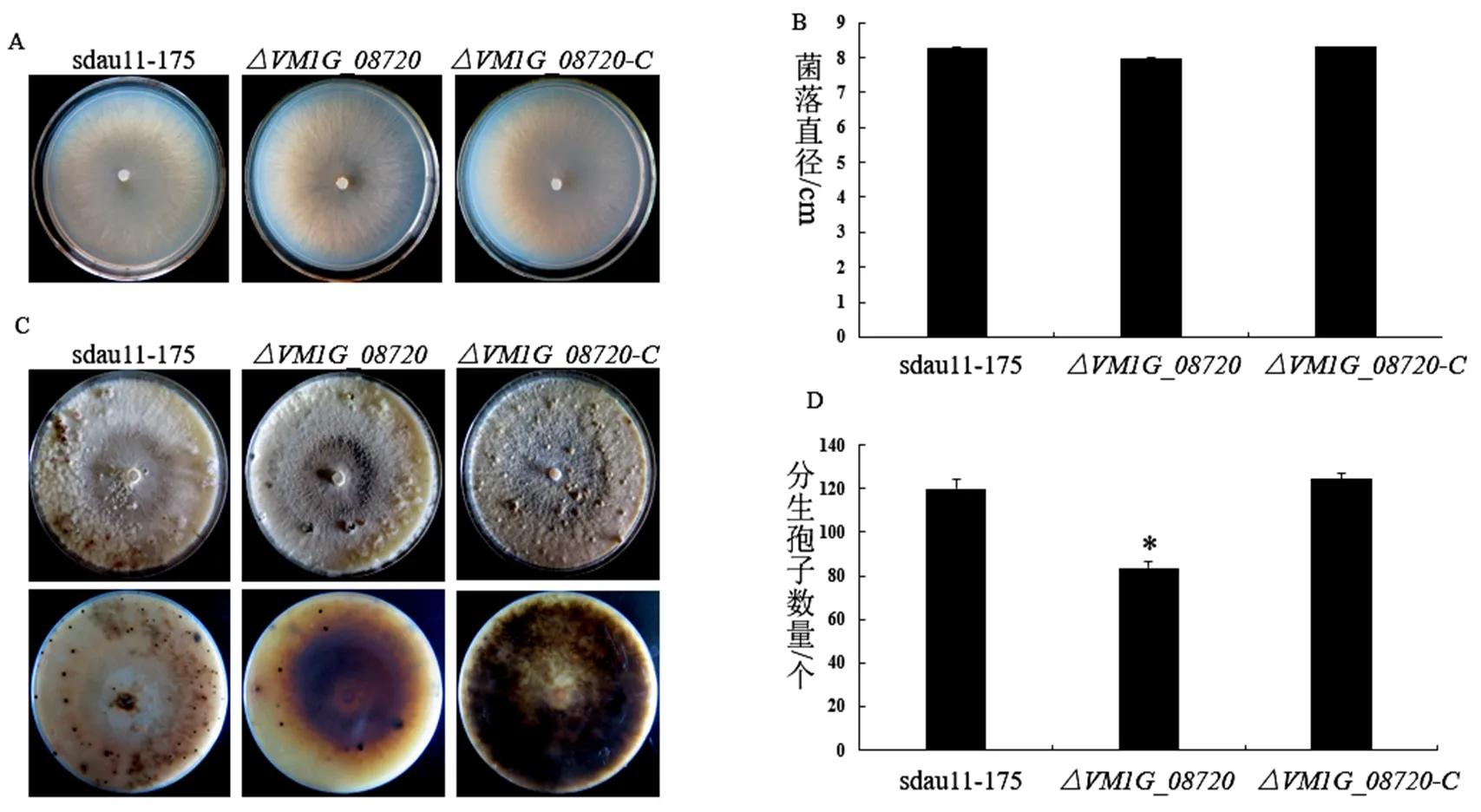
图2 野生型菌株sdau11-175、缺失突变体、回补菌株对病菌生长发育的影响
备注:A:野生型菌株sdau11-175、缺失突变体、回补菌株的菌落形态;B:野生型菌株sdau11-175、缺失突变体、回补菌株的菌落直径大小;C:野生型菌株sdau11-175、缺失突变体、回补菌株分生孢子器产生情况;D:野生型菌株sdau11-175、缺失突变体、回补菌株分生孢子器产生数量统计。
Note: A: Colony morphology of wild-type strains sdau11-175, deletion mutants and complementary strains; B: Colony diameters of wild-type strains sdau11-175, deletion mutants and complementary strains; C: Pycnidia formation of the wild-type sdau11-175, deletion mutants and complementary strains; D: Analysis pycnidial numbers on wild-type sdau11-175, deletion mutants and complementary strains.
2.3 基因VM1G_08720对病菌细胞壁完整性的影响
相对于野生型菌株sdau11-175,在含有200 μg/mL刚果红、0.01%SDS的培养基上,菌丝生长抑制率显著降低,分别降低了50%和18%;而在含有400 μg/mL荧光增白剂的培养基上,无显著差异,回补菌株恢复到野生型水平(图3)。这些结果表明基因对维持细胞壁的完整性有重要作用。
备注:A:野生型菌株sdau11-175、缺失突变体、回补菌株在不同胁迫因子下的菌落生长情况;B:野生型菌株sdau11-175、缺失突变体、回补菌株在不同胁迫因子时的菌落直径大小。
Note: A: Colony growth of wild-type strains sdau11-175, deletion mutants and complementary strains under different stress factors; B: Colony diameter of wild-type strains sdau11-175, deletion mutants and complementary strains under different stress factors.
2.4 基因VM1G_08720对病菌渗透胁迫的影响
相对于野生型菌株sdau11-175、基因缺失突变体在含有0.5 M NaCl的培养基上的菌丝生长抑制率显著增加,增加了15%,而在含有0.5 M山梨醇的培养基上无显著差异,回补菌株恢复到野生型水平(图4)。这些结果表明基因对病菌渗透胁迫有重要作用。
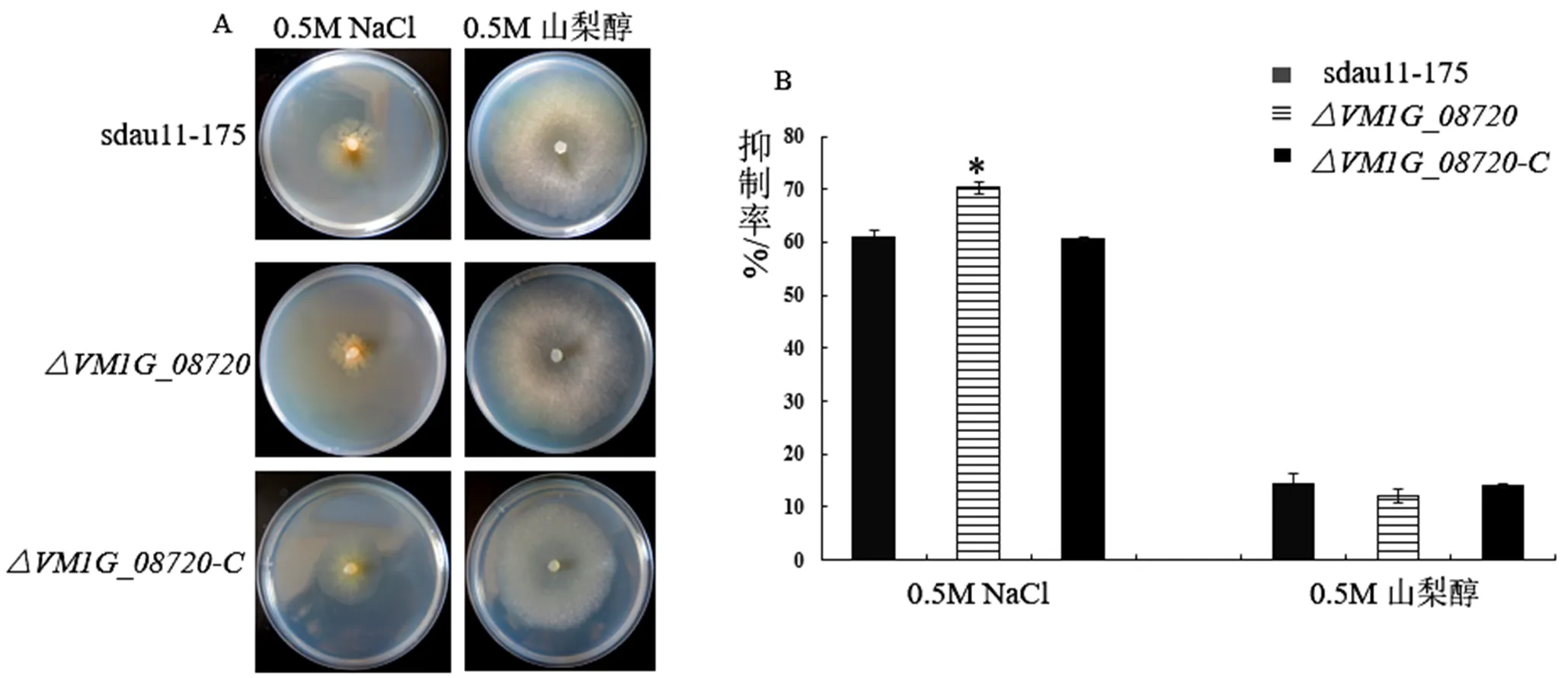
图4 基因VM1G_08720对病菌渗透胁迫的影响
备注:A:野生型菌株sdau11-175、缺失突变体、回补菌株在不同胁迫因子下的菌落生长情况;B:野生型菌株sdau11-175、缺失突变体、回补菌株在不同胁迫因子下的菌落直径大小。
Note: A: Colony growth characters of wild-type strains sdau11-175, deletion mutants and complementary strains under different stress factors; B: Colony diameters of wild-type strains sdau11-175, deletion mutants and complementary strains under different stress factors.
2.5 基因VM1G_08720对病菌酸碱适应性的影响
相野生型菌株sdau11-175、基因缺失突变体及回补菌株在pH值为4-12时均能生长。与野生型菌株sdau11-175相比,在pH为4-10时,基因缺失突变体的生长速率显著降低;而在pH为11-12时,无显著性差异,回补菌株恢复到野生型水平(图5)。
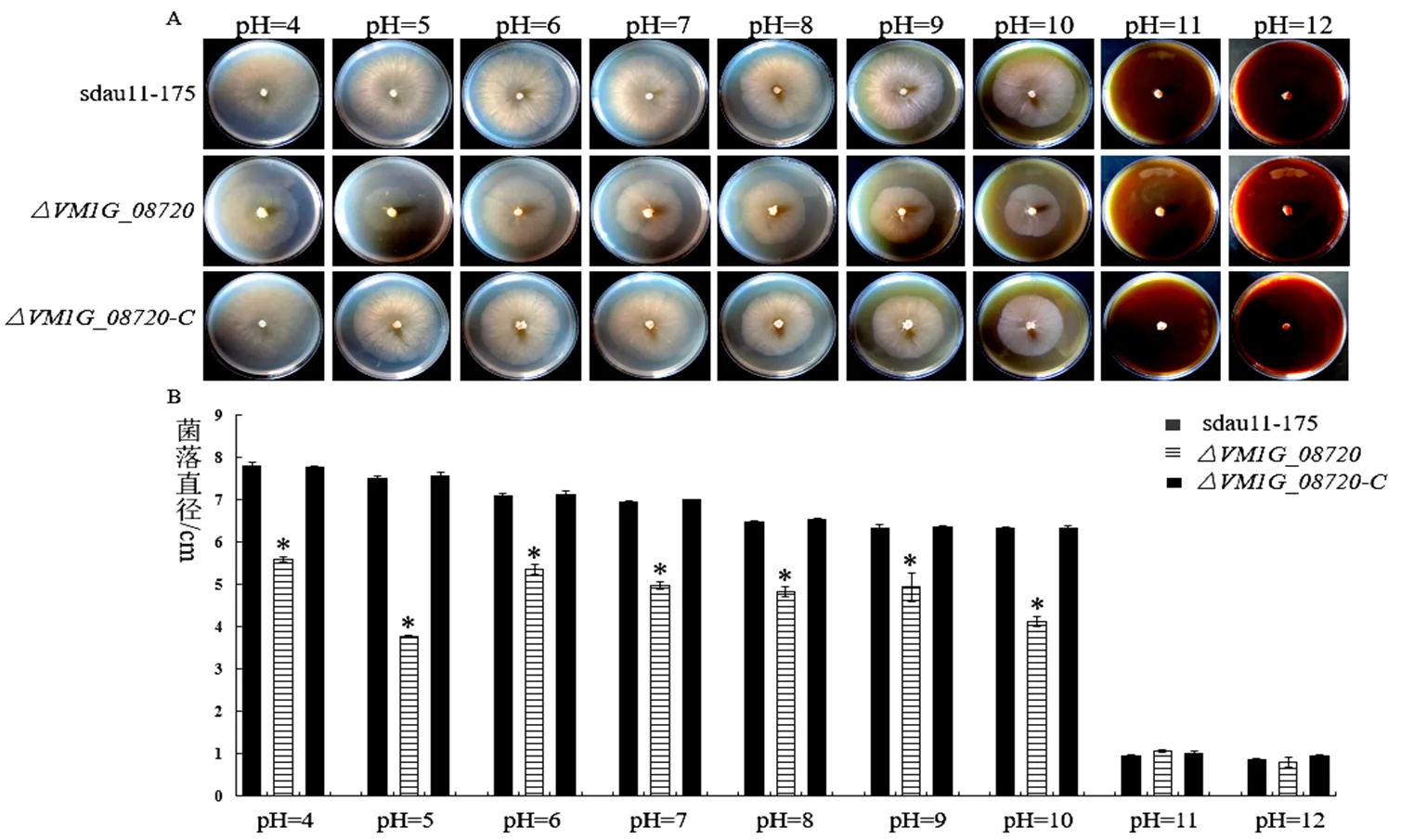
图5 基因VM1G_08720对病菌酸碱适应性的影响
备注:A:野生型菌株sdau11-175、缺失突变体、回补菌株在不同pH条件下的生长特征;B:野生型菌株sdau11-175、缺失突变体、回补菌株在不同pH条件下的菌落直径大小。
Note:A: Growth characters of wild-type strains sdau11-175, deletion mutants and complementary strains under different pH; B: Colony diameter status of wild-type strains sdau11-175, deletion mutants and complementary strains under different pH.
2.6 基因VM1G_08720对病菌致病力的影响
基因缺失突变体及回补菌株在苹果果实和苹果枝条上的病斑大小与野生型菌株sdau11-175无显著性差异(图6)。说明基因对苹果腐烂病菌致病性影响不大,调控作用不明显。
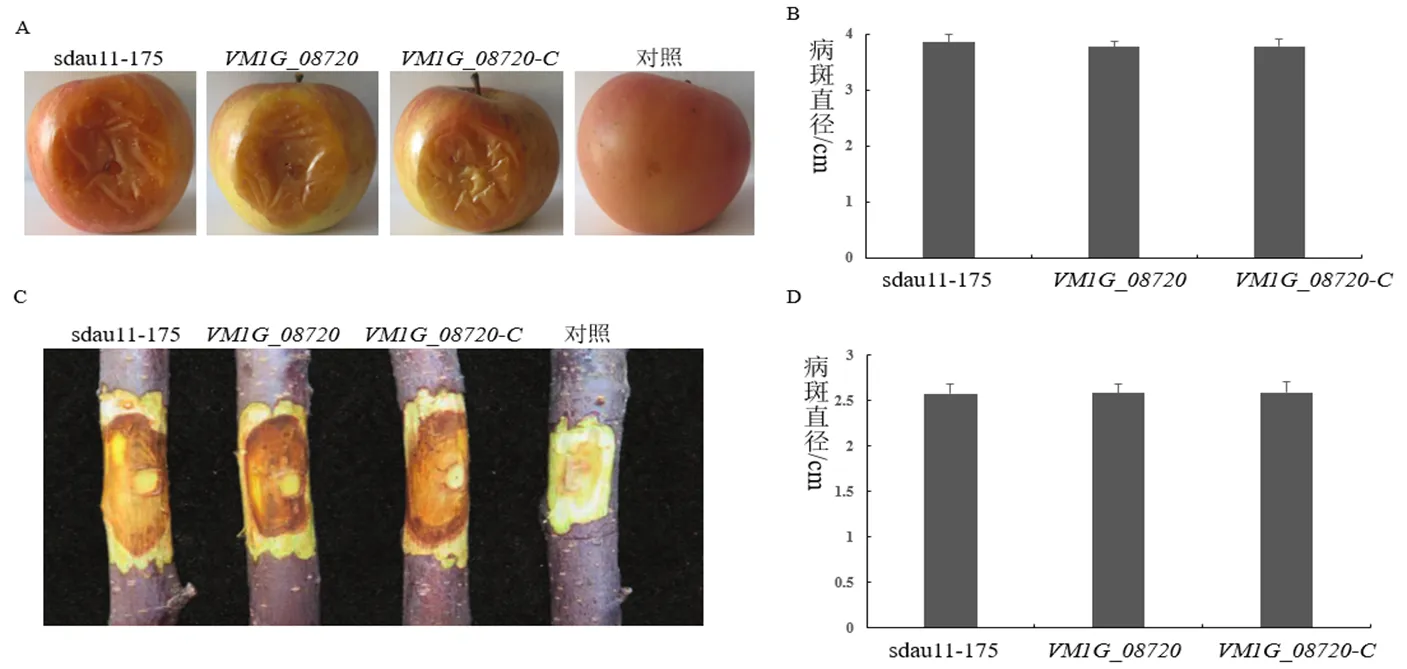
图6 基因VM1G_08720对病菌致病力的影响
备注:A:野生型菌株sdau11-175、缺失突变体、回补菌株接种苹果果实后的症状;B:野生型菌株sdau11-175、缺失突变体、回补菌株接种苹果果实后的病斑大小;C:野生型菌株sdau11-175、缺失突变体、回补菌株接种枝条后的症状;D:野生型菌株sdau11-175、缺失突变体、回补菌株接种枝条后的病斑大小。
Note: A: Symptom of wild-type strains sdau11-175, deletion mutants and complementary strains on apple fruit after inoculation; B: Lesion sizes of wild-type strains sdau11-175, deletion mutants and complementary strains on apple fruit; C: Symptom of wild-type strains sdau11-175, deletion mutants and complementary strains on twigs after inoculation; D: Lesion sizes of wild-type strains sdau11-175, deletion mutants and complementary strains on apple twigs.
3 结论与讨论
3.1 结论
本研究以基因为研究对象,通过Double-joint PCR的方法获得了缺失突变体,利用gap-repair方法获得回补菌株。通过对病菌生长发育、细胞壁完整性、渗透胁迫、酸碱度以及致病性测定,基因缺失突变体对病菌生长无影响,正调控分生孢子器的产生,在受到200 μg/ml刚果红、0.01%SDS、0.5M NaCl胁迫时,生长受到抑制;在受到400 μg/ml CFW、0.5 M山梨醇胁迫时,与野生型菌株相比无显著差异,在pH为4-10时,与野生型菌株相比生长速率显著降低;在pH为11-12时,与野生型菌株相比无显著性差异,在苹果果实和苹果枝条上的病斑大小与野生型菌株无显著性差异,其回补菌株均恢复原有功能。结果表明基因在病菌发育、维持细胞壁完整性、渗透胁迫以及酸碱适应响应方面发挥重要作用;对病菌生长、致病性调控作用不显著。
3.2 讨论
丝氨酸蛋白酶在病菌生长发育方面有着重要的作用,在烟曲霉()中,丝氨酸蛋白酶基因缺失后导致无性生长速度轻微减缓,分生孢子的产生减少,这表明丝氨酸蛋白酶在烟曲霉生长发育中发挥重要作用[5],水稻稻瘟病菌()中的丝氨酸蛋白酶基因可能存在冗余现象,丝氨酸蛋白酶基因缺失后能够产生附着胞,但分生孢子产生数量减少,而另一个丝氨酸蛋白酶基因缺失后对菌丝生长及分生孢子的产生均无影响[7]。本研究中发现,当苹果腐烂病菌丝氨酸蛋白酶基因缺失后,突变体菌丝生长无影响,但分生孢子器产生数量减少。在稻瘟病菌中,丝氨酸蛋白酶基因的缺失能够正调控细胞壁的完整性,但不参与渗透胁迫调节,本研究中基因的缺失正调控细胞壁完整性,这与稻瘟病菌研究结果相同,基因的缺失负调控渗透胁迫,这与稻瘟病菌中的研究结果不相同。烟曲霉的丝氨酸蛋白酶基因2在pH为4.5-11之间有着广泛的活性,这与本试验结果一致。本研究基因缺失突变体与野生型菌株相比,致病性无显著性差异,表明基因在致病性方面调控作用不明显,上述结果表明基因可能与其它丝氨酸蛋白酶基因互作共同调控病菌生长发育、致病性等,今后尚需对该基因参与的调控网络继续深入研究,以解析其在病菌生长发育、环境胁迫及致病性调控中的分子作用机制。
[1] 李保华,练森,王彩霞,等.从病菌自剪锯口木质部侵染再谈苹果树腐烂病的防治[J].中国果树,2017(2):88-90
[2] Wang C, Li C, Li B,. Toxins produced byvar.and their relationship with pathogenicity [J]. Toxins, 2014,6:1139-1154
[3] 何智,陈政良.补体系统丝氨酸蛋白酶的结构与功能[J].国外医学免疫学分册,2004,27(6):316-319
[4] Maxwell MK, Enrico DC. Molecular markers of serine protease evolution [J]. Embo Journal, 2001,20(12):3036-3045
[5] Shen HD, Tam MF, Chou H,. The importance of serine proteinases as aeroallergens associated with asthma [J]. Int Arch Allergy Immunol, 1999,119(4):259-264
[6] Oh Y, Donofrio N, Pan H,. Transcriptome analysis reveals new insight into appressorium formation and function in the rice blast fungus[J]. Genome Biol, 2008,9(5):R85
[7] Donofrio NM, Oh Y, Lundy R,. Global gene expression during nitrogen starvation in the rice blast fungus,[J]. Fungal Genet Biol, 2006,43(9):605-617
[8] Yin ZY, Liu HQ, Li ZP,. Genome sequence ofcanker pathogens uncovers a potental adaptation of colonization of woody bark [J]. New Phytologist, 2015,208(4):1202-1216
[9] Yu JH, Hamari Z, Han KH,. Double-joint PCR: a PCR-based molecular tool for gene manipulations in filamentous fungi [J]. Fungal Genetics and Biology, 2004,41(11):973-981
[10] 陈亮,伦莹莹,孙庚午,等.苹果腐烂病菌原生质体制备与再生条件优化[J].山东农业科学,2014,46(8):109-112
[11] Li ZP, Yin ZY, Fan YY,. Candidate effector proteins of the necrotrophic apple canker pathogencan suppress BAX-induced PCD [J]. Frontiers in Plant Science, 2015,6:579
[12] Zhou XY, Zhang HF, Li GT,. The Cyclase-associated protein Cap1 is important for proper regulation of infection-related morphogenesis in[J]. Plos Pathog, 2012,8(9):503-512
[13] 贾晓曼,王怡霖,孙庚午,等.苹果腐烂病菌基因敲除突变体获得及表型分析[J].核农学报,2018,32(3):448-454
Functional Analysis of Serine Protease Genein
DIAO Yu-fei1, XIONG Xiong2, ZHENG Jin-zhu3, JIN Ji-yang2, YU Cheng-ming1*, LIU Hui-xiang1*
1.,/,271018,2./,271018,3.271027,
Apple is one of the most widely planted temperate fruits in China.canker of apple caused byis one of the most serious diseases of apple tree in China. Therefore, strengthen the scientific prevention and control of, and clarify the adaptability and pathogenic effect of pathogens to the environment is the key problem for diease. In this paper, Double-joint PCR and gap-repair methods were used to obtain deletion mutants and complement strains of the gene. The effect of the geneon nutritional growth and pathogenicity was investigated used the cross-over method and the wound inoculation. The role on the maintenance of cell wall integrity and the regulation of osmotic stress of the genewere investigated by adding 400 μg/mL calcofluor white, 200 μg/mL Congo red, 0.01% SDS, and 0.5 M NaCl, 0.5 M sorbitol. The effect of the geneon the hyphal growth of the pathogen under different pH was investigated by setting the medium for different gradient. The results showed that the genedeletion mutant was no significant differences in colony color, mycelium density, growth rate compared with wild-type sdau11-175. The numbers of pycnidial production were reduced. The inhibition rate was significantly reduced on medium containing 200 μg/mL of congo red and 0.01% SDS, and the inhibition increased significantly on medium containing 0.5 M NaCl, and there was no significant difference in the culture medium containing 400 μg/mL calcofluor white and 0.5 M sorbitol. Growth rate was significantly reduced at pH 4-10, with no significant difference at pH 11-12. There was no significant difference in diseased spot sizes between mutant and wild strain on apple fruit and apple branches. All the complement strains were recovered to wild-type levels. It indicates that the geneplays an important role in pycnidial production, pH response, maintenance of cell wall integrity, and osmotic stress.
;gene; serine proteinase; phenotype analysis
S436.611.1+1
A
1000-2324(2022)05-0735-07
2022-06-25
2022-06-05
国家自然基金项目(31770684);山东省自然科学基金项目(ZR2017MC042)
刁雨菲(1996-),女,博士研究生,专业方向林果病原与寄主分子互作机制研究. E-mail:1031151284@qq.com
通讯作者:Author for correspondence. E-mail:ycm2006.apple@163.com; hxliu@sdau.edu.cn
