滩羊肉嫩度形成中Ca2+的信号转导作用
2023-01-12姬琛尤丽琴刘吉娟罗瑞明
姬琛,尤丽琴,刘吉娟,罗瑞明*
(1 宁夏大学农学院 银川750021 2 宁夏大学食品与葡萄酒学院 银川 750021)
嫩度是衡量肉品品质的重要指标,贮藏过程中肉品嫩度的变化主要是由于内源酶引起的结构蛋白质的水解[1]。钙蛋白酶(Calpains)是公认的结构蛋白水解酶,钙蛋白酶-1 参与大多数肌原纤维蛋白的降解过程,引起肌肉嫩度约70%的差异。细胞内钙蛋白酶-1 的激活依赖于Ca2+浓度[2],其被激活后可降解多种细胞骨架蛋白,如伴肌动蛋白、肌钙蛋白T 和肌间线蛋白等。细胞凋亡酶(Caspases)被认为在细胞骨架蛋白的降解中发挥重要作用[3-4]。Huang 等[3]和Ouali 等[4]研究表明细胞凋亡效应酶-3 有助于弱化Z 盘和I 盘之间的连接,从而影响肉的嫩度。细胞凋亡酶的激活依赖于内源性细胞凋亡引起的级联反应,线粒体Ca2+超载是引起内源性细胞凋亡的重要原因。因此,宰后贮藏期间Ca2+的信号转导作用对肉品嫩度形成具有重要意义。
蛋白质组学分析为揭示肌肉转化为肉所经历的分子修饰提供了强有力的基础[6]。Zuo 等[7]采用蛋白质组学对不同保水性的牦牛肉进行分析,发现肌球蛋白轻链、热休克蛋白27 和磷酸丙糖异构酶可作为牦牛肉持水能力的生物标志物。Della等[8]采用蛋白质组学研究宰后成熟过程中马不同部位肌肉蛋白质的降解情况,发现半腱肌的水解程度更剧烈。LÓPEZ-PEDROUSO 等[9]基于SWATH-MS 研究发现糖异生、糖酵解及三羧酸循环是影响牛肉嫩度的重要途径。罗辉等[10]发现秦川牛肉贮藏过程中蛋白质变化引起的能量代谢物变化,对肉品质形成具有一定贡献。李升升[11]研究发现钙蛋白酶抑制蛋白、SLC25A4、ZYX、LMOD1等可作为牦牛平滑肌嫩度的指示蛋白。然而,现有研究多为肉品嫩度生物标志物鉴定,蛋白质水平变化与肉品嫩度的关系,影响肉品嫩度形成的主要途径等,缺乏关键信号因子Ca2+在嫩度形成中发挥信号转导作用的研究。
本试验以6 月龄滩羊背最长肌为研究对象,测定其4 ℃贮藏0,96,192 h 时剪切力、线粒体Ca2+水平、钙蛋白酶-1 活性及细胞凋亡效应酶-3活性的变化情况,基于同位素标记相对和绝对定量技术(iTRAQ)结合多维液相色谱-串联质谱(LC-MS/MS)技术研究贮藏过程中Ca2+信号通路及凋亡调节信号转导通路蛋白质表达量变化,确定Ca2+在滩羊肉嫩度形成方面的信号转导作用,为进一步完善宰后肉品嫩度形成理论,改善滩羊肉嫩度提供参考。
1 材料与方法
1.1 材料与试剂
滩羊背最长肌,宁夏盐池县大夏牧场食品有限公司提供。
尿素、二硫苏糖醇、碘乙酰胺、IPG buffer、甲酸,美国GE Healthcare 公司;十二烷基磺酸钠、三羧基氨基甲烷、三氯乙酸、过硫酸铵、四甲基乙二胺,美国Amresco 公司;三乙二胺碳酸盐和考马斯亮蓝G-250,美国Sigma 公司;胰蛋白酶,美国Promega 公司;乙腈、水,美国赛默飞世尔公司;甘露醇、EDTA、硝酸、浓盐酸、浓硫酸(分析纯),国药集团;Elisa 试剂盒,上海科兴生物科技有限公司。未注明试剂均为色谱纯级。
1.2 仪器与设备
Sciex Triple TOF 5600 质谱仪、Eksigent nano LC-1D 液相系统,美国SCIEX 公司;Image Scanner III 扫描仪,美国GE Healthcare 公司;iCE 3400 AAS 原子吸收光谱仪,美国赛默飞世尔公司;VCX130 超声波细胞粉碎机,美国Sonics 公司;C-LM40 嫩度仪,北京布拉德科技发展有限公司5910R 冷冻离心机,德国Eppendorf 公司;真空冷冻干燥机,小垂直板电泳槽,美国伯乐公司。。
1.3 方法
1.3.1 样品采集 采集宰后4 ℃,风速3 m/s,相对湿度80%,成熟0,96,192 h 的滩羊背最长肌200 g,用于测定剪切力。采集不同成熟时间样本各5 g,封装于灭菌冷冻管中,置于-80 ℃冰箱备用,用于测定钙蛋白酶-1 及细胞凋亡效应酶-3 活性。采集不同成熟时间样本200 mg,封装于灭菌冷冻管中,置于-80 ℃冰箱备用,用于蛋白质组学分析。
1.3.2 蛋白质组学分析 参照李子欣等[12]的方法提取纯化滩羊肉中蛋白质,置于-80 ℃冰箱备用。参照尤丽琴等[13]的方法检索并筛选差异蛋白质。差异蛋白质的生物信息学分析通过京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)通路注释完成。
1.3.3 线粒体Ca2+含量的测定 参照王琳琳[14]的方法分离线粒体,测定其Ca2+含量。
1.3.4 钙蛋白酶-1 及细胞凋亡效应酶-3 活性测定 采用Elisa 试剂盒,根据操作说明书分别测定钙蛋白酶-1 活性及细胞凋亡效应酶-3 活性。
1.3.5 剪切力的测定 剪切力的测定采用Di Luca 等[15]的方法并稍作修改。剔除可见脂肪与结缔组织,将样品修整为5 cm×5 cm×5 cm 的块状,置于自封袋中,于80 ℃水浴锅中煮制。待中心温度到达70 ℃后取出样品,室温放置12 h 后用取样器平行于肌纤维方向取直径为1.27 cm 的肉柱,以1 mm/s 垂直于样品剪切肉柱,测定其剪切力。
1.4 统计分析
每个指标重复测定3 次,结果以平均值±标准偏差(Standard Deviation,S.D.)表示。采用SPSS 软件对数据进行方差分析、邓肯多元方差检验以及Pearson 相关性分析。P <0.05 的变量被认为不同成熟时间之间存在显著差异。
2 结果与分析
2.1 剪切力、线粒体Ca2+含量、钙蛋白酶-1 活性及细胞凋亡效应酶-3 活性变化
表1 所示不同贮藏时间滩羊背最长肌剪切力、线粒体Ca2+含量、钙蛋白酶-1 活性及细胞凋亡效应酶-3 活性变化情况。贮藏0 h 时,滩羊背最长肌剪切力为4.79 kgf;贮藏96 h 时,滩羊背最长肌剪切力显著降低至3.17 kgf(P<0.05);贮藏192 h时,滩羊背最长肌剪切力显著升至3.74 kgf(P<0.05),说明适度的贮藏有助于改善滩羊肉嫩度,这与李文博等[16]的结论一致。0~96 h 内剪切力降低,可能是由于构成肌肉结构的蛋白质在贮藏期间发生降解[17]。

表1 不同贮藏时间滩羊肉剪切力、线粒体钙离子水平、钙蛋白酶-1 活性及细胞凋亡效应酶-3 活性变化Table 1 Changes in shear force,content of mitochondria Ca2+,and activities of calpain-1 and caspase-3
线粒体Ca2+含量贮藏0 h 为2.23 mg/L,96 h显著上升至2.52 mg/L(P<0.05),192 h 时又降至2.20 mg/L(P<0.05),表明0~96 h 内滩羊肌细胞线粒体发生Ca2+超载,与王琳琳[14]的研究结果一致。Ca2+主要储存于内质网,正常的离子稳态是维持细胞生命活动的基本条件,线粒体摄取生理性Ca2+信号调节有氧代谢,而宰后Ca2+超载会引起细胞程序性死亡,即细胞凋亡过程[4]。同时,Ca2+浓度可调节细胞骨架蛋白水解酶钙蛋白酶活性。
钙蛋白酶-1 活性在整个贮藏期呈显著降低趋势(P<0.05),96 h 时显著降至初始值的14%,192 h 时显著降至初始值的6%。大量研究表明,Ca2+浓度升高促使钙蛋白酶活性增强,Ca2+浓度进一步提高导致构象变化而表现蛋白水解酶活性,是宰后贮藏初期细胞质中主要的蛋白水解酶,在肌原纤维蛋白降解及肉品嫩度形成中发挥重要作用[4]。钙蛋白酶-1 最适pH7.0 左右[22],因此,除Ca2+外宰后肌肉pH 值的变化也影响钙蛋白酶-1 活性,其活性下降可能与宰后初期细胞环境酸化有关。
细胞凋亡效应酶-3 活性于贮藏期内呈先升高后降低的趋势,96 h 时显著上升至初始值的130%(P<0.05),192 h 时活性为初始值的116%(P<0.05),表明线粒体Ca2+超载可能引起细胞凋亡级联反应,使细胞凋亡效应酶-3 被激活,且直至宰后192 h 其活性仍显著高于初始活性。黄明等[23]证实细胞凋亡效应酶-3 可体外降解结构蛋白,参与肉品宰后的嫩化。
2.2 剪切力与线粒体Ca2+水平、钙蛋白酶-1 活性及细胞凋亡效应酶-3 活性的相关性分析
表2 所示为剪切力与线粒体Ca2+水平、钙蛋白酶-1 活性及细胞凋亡效应酶-3 活性的相关性分析。滩羊背最长肌剪切力与Ca2+水平呈显著负相关(P<0.05),与钙蛋白酶-1 活性呈极显著正相关(P<0.01),与细胞凋亡效应酶-3 活性呈极显著负相关(P<0.01);钙蛋白酶-1 活性与细胞凋亡效应酶-3 活性呈极显著负相关(P<0.01),与Ca2+水平与无明显相关性;细胞凋亡效应酶-3 活性与Ca2+水平呈显著正相关(P<0.05)。钙蛋白酶-1 活性不受Ca2+水平影响,可能是因为钙蛋白酶-1 活性受细胞环境影响因素如pH 值、ATP 含量等较大。细胞凋亡效应酶-3 活性受细胞凋亡因子的调控,线粒体Ca2+超载是促使CYTC 释放至胞浆中的重要原因,在细胞凋亡效应酶-3 激活过程中具有正向调控作用。钙蛋白酶-1 与细胞凋亡效应酶-3 均为水解肌原纤维的蛋白质,肌原纤维蛋白质降解由肌原纤维解装配成肌丝启始,大量研究表明钙蛋白酶参与这一解装配过程,因此,钙蛋白酶-1 可能参与成熟初期肌原纤维蛋白的降解,而细胞凋亡效应酶-3 在整个贮藏过程中均可水解肌原纤维。
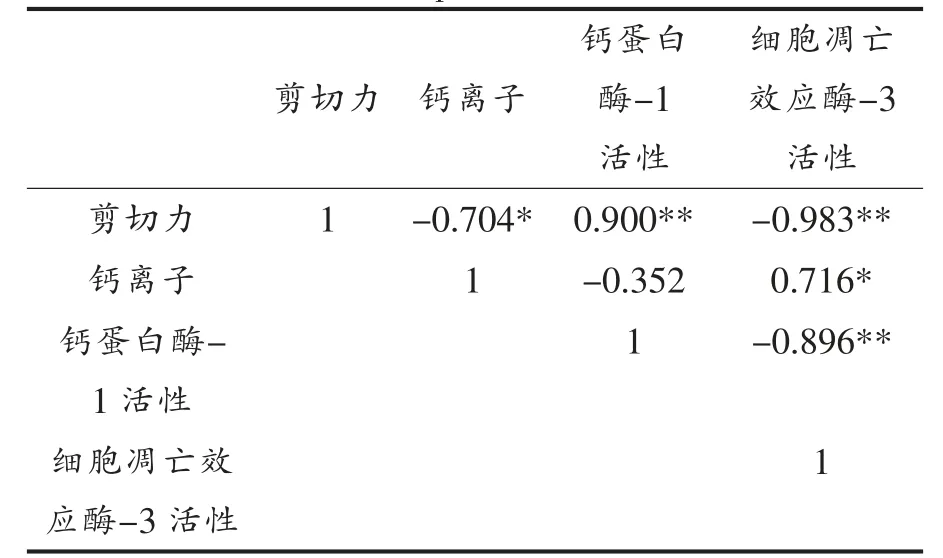
表2 不同贮藏时间滩羊肉剪切力与线粒体钙离子水平、钙蛋白酶-1 及细胞凋亡效应酶-3 活性相关性分析Table 2 Correlations between shear force and content of mitochondria Ca2+ and activities of calpain-1 and caspase-3
2.3 Ca2+信号通路及凋亡调节信号转导通路关键蛋白质的变化
为明确Ca2+在宰后的信号转导作用,通过蛋白质组学分析鉴定Ca2+信号通路及凋亡相关信号转导通路蛋白质表达的变化情况,结果显示:贮藏96 h 与0 h 对比,鉴定出7 个显著差异蛋白质富集于Ca2+信号通路,其中,5 个上调表达差异蛋白质,2 个下调表达差异蛋白;192 h 与96 h 对比,鉴定出3 个显著差异蛋白质富集于Ca2+信号通路,均为下调表达差异蛋白质(表3)。其中,ATP2A1即Ca2+-ATPase,主要催化ATP 的水解,并将Ca2+由细胞质转移至内质网钙库,参与调节收缩/舒张周期[24],其上调表达可能是因为动物屠宰后细胞环境及能量水平的改变导致细胞内离子平衡被打破,引起大量离子交换,ATP2A1 试图将Ca2+逆浓度梯度泵入钙库。VDAC1 与VDAC2 为电压依赖性阴离子通道蛋白1 与2,是线粒体通透性转换孔(mPTP)的构成部分[25],调控mPTP 的开放,其表达量上调可能导致线粒体内膜不完整并引起mPTP 不可逆开放,线粒体Ca2+超载可能由此引起。SLC25A4 参与线粒体ADP/ATP 的转移,PPP3CA 为钙依赖性酶,在细胞内Ca2+介导的信号转导中起重要作用。MYLK2 是一种钙/钙调蛋白依赖的酶,与肌肉收缩相关[26]。不同贮藏期Ca2+信号通路的变化表明,屠宰后初期滩羊肌细胞内离子稳态失衡,Ca2+刺激下游一系列蛋白质表达的变化,引起不同时期肌细胞收缩、肌纤维降解等现象,从而调节动物屠宰后肌肉的嫩度。
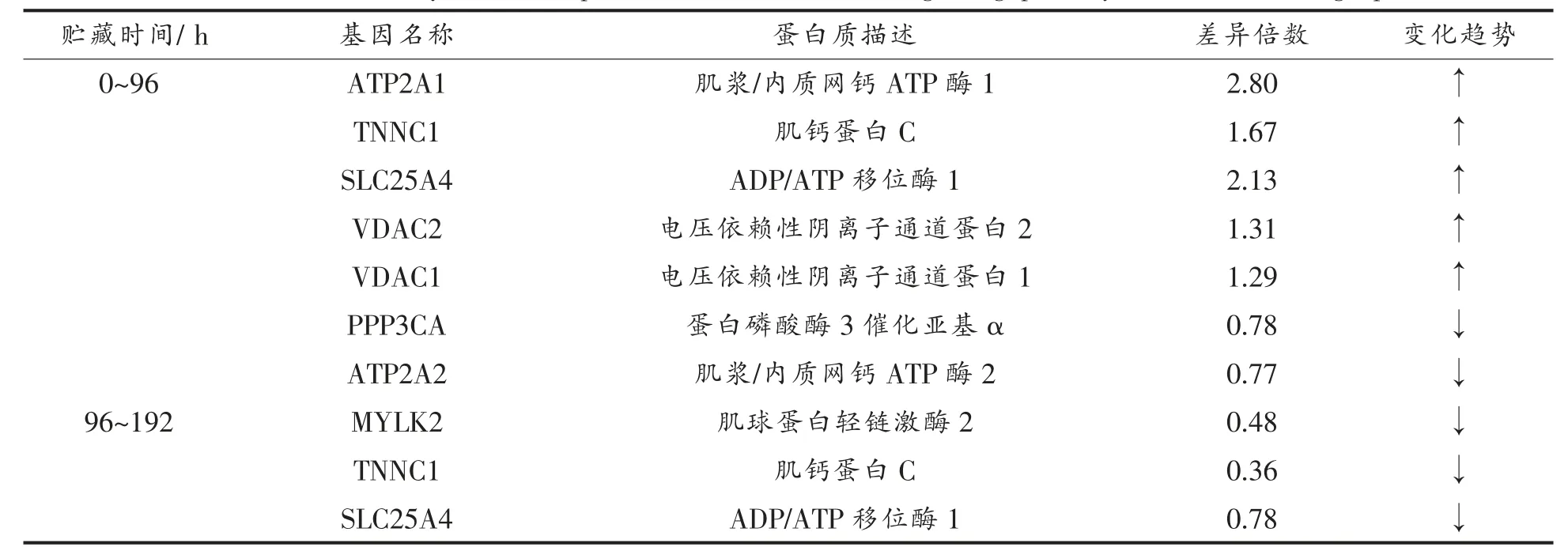
表3 不同贮藏期Ca2+信号通路中富集的差异表达蛋白质Table 3 The differentially abundant proteins enriched in Ca2+ signaling pathway at different storage periods
表4 所示凋亡调节信号转导通路相关蛋白质的变化。0~96 h 内30 个显著差异表达蛋白质富集于凋亡调节信号转导通路,其中,23 个上调表达的差异蛋白质,7 个下调表达的差异蛋白质。96~192 h 内15 个显著差异表达蛋白质富集于凋亡调节信号转导通路,其中6 个上调表达的差异蛋白质,9 个下调表达的差异蛋白质。凋亡信号调节转导通路富集的差异蛋白质表明,滩羊屠宰后受缺氧缺血信号的刺激,线粒体发生不可逆的损伤,线粒体复合体Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ表达量上调,试图通过将NADH 转换为NAD+与H+调节细胞内离子稳态,降低合成反应所引起的ATP 消耗,由此引起H+积累[27],这可能是导致96 h 内钙蛋白酶-1 活性降低的重要原因之一。王霖[28]研究表明胞质或线粒体Ca2+升高可能会破坏线粒体电子传递体,这与本研究结论一致。CYTC 在线粒体嵴突上与其它氧化酶排列成呼吸链,作为电子传递体之一参与有氧呼吸,在能量代谢调节中扮演重要角色,也是线粒体凋亡途径的重要调控因子。CYTC 从线粒体释放到胞浆中,会导致电子呼吸链的电子传递受阻,引发细胞能量供应减少。VDAC1 与VDAC2 作为mPTP 蛋白,其上调表达可能为CYTC 释放至胞浆中提供了渠道,由此启动凋亡级联反应[29],激活细胞凋亡效应酶-3。Wang 等[30]研究表明,CYTC宰后初期从牦牛肌细胞线粒体释放至胞浆中,最终激活细胞凋亡效应酶-3,并影响肌肉嫩度,而线粒体Ca2+超载是触发CYTC 释放的重要原因。CYTC 释放及细胞色素C 氧化酶亚基II(COX2)下调表达,可能是引起宰后电子传递链解偶联的主要原因。胞浆中CYTC 表达量上调引起的电子传递受阻,可能与宰后能量供给方式转变有关。

表4 不同贮藏期凋亡调节信号转导通路中富集的显著差异表达蛋白质Table 4 The differentially abundant proteins enriched in apoptosis signal transduction pathways at different storage periods
不同贮藏时间显著差异蛋白质KEGG 数据库映射的细胞凋亡通路(KEGG#04210)见图1。贮藏初期,CYTC 由线粒体释放至胞浆,引起胞浆中CYTC 表达量升高,由此可能导致凋亡级联反应。细胞凋亡效应酶-3 活性于96 h 时升高,与胞浆中CYTC 表达量升高有关。Ca2+由内质网释放引起钙蛋白酶表达量下调,而Ca2+浓度持续上升会引发钙蛋白酶-1 自溶[31],这可能是导致其表达量下调的主要原因。肌动蛋白于贮藏初期表达量下调可能与肌动球蛋白复合体形成有关[32]。真核翻译起始因子2 亚基1 负责真核生物翻译起始,其下调表达可能是因为宰后细胞生命活动与活体有区别所致,也可能是因为动物屠宰后蛋白质的持续降解。大量研究表明,动物屠宰后初期胞浆中CYTC表达量上升,本研究结果符合这一规律。贮藏96 h后,释放至胞浆的CYTC 减少。王琳琳[14]研究表明牦牛宰后72 h 胞浆中CYTC 表达量显著下降约60%。本研究中CYTC 变化趋势与其一致。肌动蛋白表达量上调可能与热休克蛋白引起应激防护作用有关。
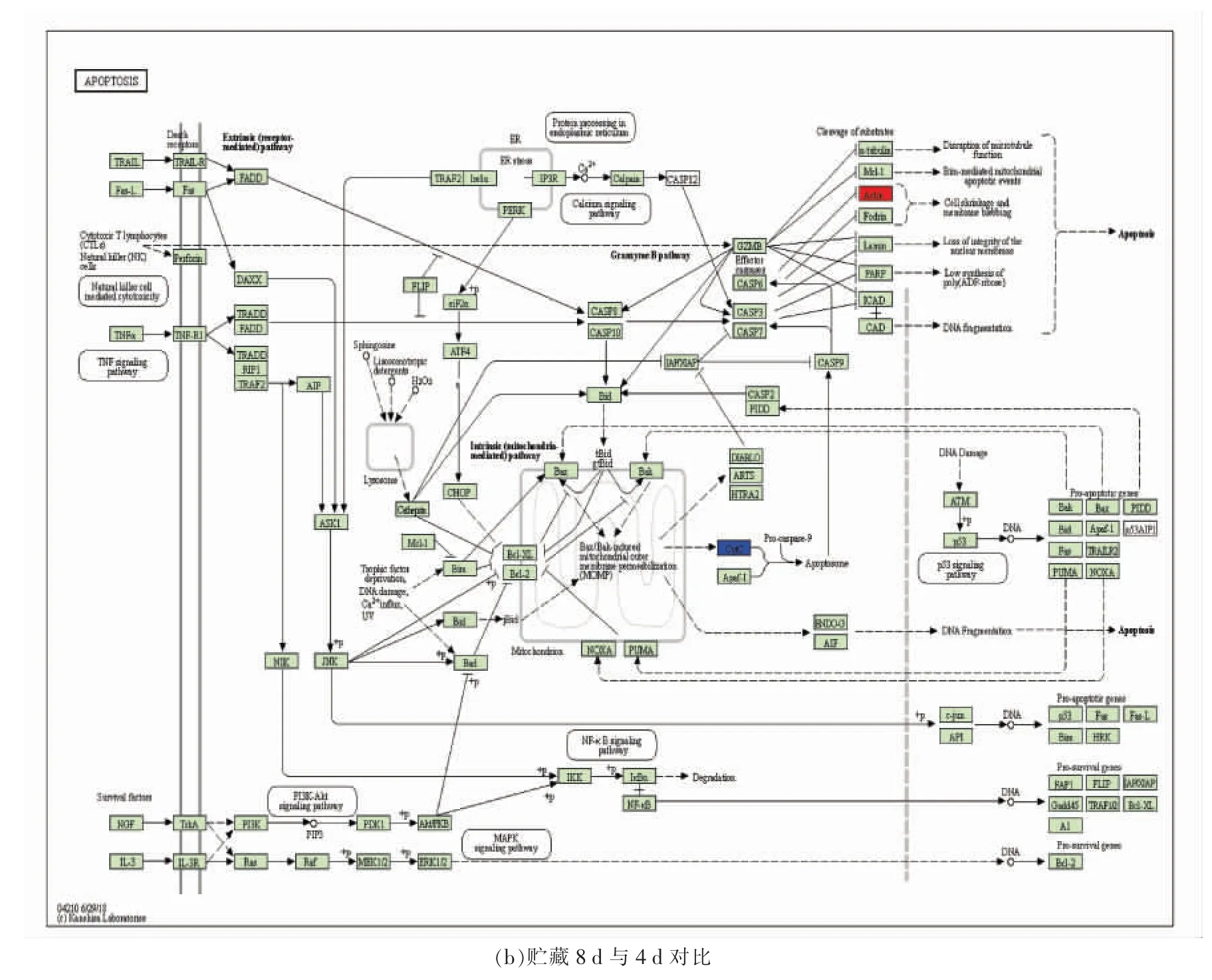
图1 不同贮藏时间滩羊肉中差异蛋白质映射的细胞凋亡通路(KEGG#04210)Fig.1 Apoptosis of differential abundant proteins mapping in Tan Sheep meat at different storage period(KEGG#04210)
2.4 Ca2+的信号转导作用及其在嫩度形成中的贡献
为进一步阐明Ca2+的信号转导作用,基于以上关键通路中富集到的差异蛋白质,以KEGG pathway 为载体,绘制Ca2+信号转导核心网络于图2。由图2 可知,SERCA 利用ATP 水解的能量驱动Ca2+跨膜运输,mPTP 由多个电压依赖性阴离子通道蛋白构成,贮藏初期上调表达为Ca2+流入线粒体提供渠道,导致线粒体Ca2+超载、功能紊乱,使线粒体复合体表达量上升,NADH 转化为NAD++H+,细胞内出现H+积累,而COX2 表达量下调促使电子传递链解偶联,导致宰后细胞能量供给方式转变;同时,mPTP 蛋白质异常表达促使CYTC 释放至胞浆中,可能引起细胞凋亡级联反应,因此激活细胞凋亡效应酶-3,水解结构蛋白质,影响肉的嫩度。Ca2+可间接导致肌动蛋白表达量下调引起细胞收缩,从而影响肉的嫩度。另外,由Ca2+信号转导核心网络图可知,钙蛋白酶与细胞凋亡效应酶-3 可能在水解蛋白质方面具有协同作用[33]。Huang等[34]研究发现,当牛骨骼肌中注射Ca2+时,细胞凋亡效应酶-3 活性呈下降趋势,可能是由于calpain的直接切割导致细胞凋亡效应酶-3 失活。这一结果充分证明Ca2+在嫩度形成过程中重要的信号转导作用,SERCA、CYTC、VDAC、COX2 等多个蛋白质在其信号传递过程中发挥重要作用。

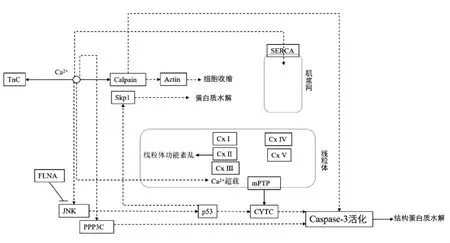
图2 滩羊肉宰后嫩度形成中Ca2+信号转导核心网络Fig.2 Core network of calcium signal transduction in postmortem tenderization of Tan sheep meat
3 结论
滩羊肉4℃贮藏期间剪切力先降低后升高,96 h 时降至最低。线粒体Ca2+含量于宰后96 h 内显著升高,随后降低,钙蛋白酶-1 活性在贮藏期内持续降低,细胞凋亡效应酶-3 活性于0~96 h 显著上升,96~192 h 有所下降。Pearson 相关性分析表明滩羊背最长肌剪切力与Ca2+水平呈显著负相关,与钙蛋白酶-1 活性呈极显著正相关,与细胞凋亡效应酶-3 活性呈极显著负相关。蛋白质组学分析表明,0~96 h 内5 个显著上调差异蛋白质与2 个显著下调差异蛋白质富集于Ca2+信号通路,96~192 h 内3 个显著下调的差异蛋白质富集于Ca2+信号通路;0~96 h 内30 个显著差异表达蛋白质富集于凋亡调节信号转导通路,96~192 h 内15个显著差异表达蛋白质富集于凋亡调节信号转导通路。SERCA 利用ATP 水解的能量驱动Ca2+跨膜运输,mPTP 蛋白质VDAC 贮藏初期上调表达为Ca2+流入线粒体提供渠道,导致线粒体Ca2+超载、线粒体功能紊乱,致线粒体复合体表达量上升;同时,mPTP 蛋白质异常表达促使CYTC 释放至胞浆中,最终激活细胞凋亡效应酶-3,水解结构蛋白质影响肉的嫩度。另外,Ca2+可间接促使肌动蛋白表达量下调引起细胞收缩,从而影响肉的嫩度。本研究结论证实了Ca2+在肉品嫩度形成过程中重要的信号转导作用。
