Three New Species of Diploderma Hallowell,1861 (Reptilia: Squamata:Agamidae) from the Shaluli Mountains in Western Sichuan,China
2023-01-05BoCAIMeihuaZHANGJingLISimengDUFengXIEMianHOUHuamingZHOUandJianpingJIANG
Bo CAI ,Meihua ZHANG, Jing LI ,Simeng DU ,Feng XIE ,Mian HOU,Huaming ZHOU and Jianping JIANG
1 CAS Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization, Ecological Restoration and Biodiversity Conservation Key Laboratory of Sichuan Province, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041,Sichuan, China
2 Ganzi Institute of Forestry Research, Kangding 626001, Sichuan, China
3 College of Continuing (Online) Education, Sichuan Normal University, Chengdu 610068, Sichuan, China
4 University of the Chinese Academy of Sciences, Beijing 100049, China
5 Mangkang Biodiversity and Ecological Station, Tibet Ecological Safety Monitor Network, Changdu 854500, Tibet, China
Abstract In recent years,taxonomic research of the genus Diploderma has made great progress,but there are still areas lacking relevant investigations.During 2020–2021,we carried out survey of reptile diversity in and around the Shaluli Mountains and collected some specimens of Diploderma.Of which,based on systematics inferred from mitochondrial ND2 gene sequence and morphological data,three new species were recognized:Diploderma daochengense sp.nov.from the Shuiluo River and Muli River valleys in southeastern Daocheng County,D.xinlongense sp.nov.from the upper-middle valleys of the Yalong River in Xinlong County,and D.kangdingense sp.nov.from the Yalong River middle valley and Liqiu River valley.Specifically,these three new species formed three monophyletic lineages with strongly supported and nested into the subclade I of the Bayesian and Maximum Likelihood trees.The uncorrected mean genetic distance between these new species and their closely related species was 4.3% (D.xinlongense sp.nov.vs.D.panchi),3.4% (D.daochengense sp.nov.vs.D.yulongense) and 5.5% (D.kangdingense sp.nov.vs.D.bowoense),higher than many pairs of species of Diploderma.With respect to the morphology,D.daochengense sp.nov.can be easily diagnosed from D.yulongense by having smooth upper edge of the dorsolateral stripes,cyan or dark olive green to yellow gular spots in both sexes and light yellow or white ventrolateral body in males;D.xinlongense sp.nov.can be easily diagnosed from D.panchi by having cyan gular spot in both sexes;D.kangdingense sp.nov.can be easily diagnosed from D.bowoense by having more T4S 20–25 (average 23),more F4S 16–20 (average 17) and yellow to light yellow ventrolateral body,yellow gular spot present in males.To date,the number of species of the genus Diploderma has increased to 40,of which 37 species have been known in China.
Keywords Ganzi,taxonomy,Hengduan Mountains,Mountain Dragons,RGB color model
1.Introduction
Species diversity is an important part of biodiversity,and species taxonomy is fundamental to biodiversity researches,further contributing to systematics,biogeography,evolution,ecology and conservation biology (Chiarucciet al.,2011;Jetzet al.,2019).The Mountain Lizard genusDiplodermaHallowell,1861 currently contains 37 described species,and is widely distributed throughout China,Myanmar,Laos,Thailand,Vietnam and Japan (Uetzet al.,2022;Caiet al.,2022;Wanget al.,2022),of which,34 species are known in China (Wanget al.,2021;Caiet al.,2022;Wanget al.,2022).
The Shaluli Mountains are lied in the eastern part of the Qinghai-Tibet Plateau and in the middle of Hengduan Mountain,and is surrounded by the Jinsha and Yalong Rivers.To our knowledge,this mountain range has a high level of biodiversity,to date,just in the genusDiploderma,this region is known to harbor 13 species,i.e.,D.panchi,D.bowoense,D.angustelinea,D.panlong,D.swild,D.dymondi,D.flavilabre,D.batangense,D.formosgulae,D.aorun,D.qilin,D.yulongenseandD.brevicauda(Wanget al.,2021;Caiet al.,2022).Most of these species were found near the banks of the Jinsha River,but branches of Jinsha River and the middle,upper and interior regions of Yalong River still lack investigation.So the species diversity in this region might be underestimated.
In 2020 and 2021,we conducted a series of field surveys in and around the Shaluli Mountains,and collected a series of specimens ofDiplodermaspecies.Based on molecular phylogenetic analyses and morphological comparisions,these specimens contained three undescribed species.Herein,we describe them as three new species.
2.Materials and Methods

Figure 1 Distributions of Diploderma species in and around the Shaluli Mountains.The stars represent the holotype localities of new species described in this paper (8: D.daochengense sp.nov.,9: D.xinlongense sp.nov.,11: D.kangdingense sp.nov.);the big circles represent the holotype localities of the remaining recognized congeners (1: D.flavilabre;2: D.batangense,3: D.formosgulae,4: D.aorun,5: D.qilin,6: D.brevicauda,7: D.yulongense,10: D.panchi,12: D.bowoense,13: D.angustelinea,14: D.panlong,15: D.swild,16: D.dymondi);the small circles represent the known localities of new species,their sister species and morphologically similar species;and the dotted oval line represents the approximate known range of that species.

Table 1 Samples used in the molecular study with new species indicated in bold and arranged in top-down order within a species.

Continued Table 1

Continued Table 1

Continued Table 1

Continued Table 1
2.1.SamplingA total of 28 specimens were collected from in and around the Shaluli Mountains of Sichuan Province,China (Figure 1,Table 1).After taking photographs of these live animals to document their color patterns,they were euthanized by MS-222.Liver tissue samples were taken and preserved in 95% ethanol for molecular analyses.Other tissue samples were stored in 99% ethanol and specimens were preserved in 75% ethanol.All specimens were deposited at the Museum of Herpetology,Chengdu Institute of Biology (CIB),Chinese Academy of Sciences (CAS).Field collection of specimens was authorized by the Forestry and Grassland Bureau of Ganzi Tibetan Autonomous Prefecture,Sichuan Province,China.The animal care and use protocols have been reviewed and approved by Animal Ethical and Welfare Committee of Chengdu Institute of Biology,CAS (Number: 2017-AR-JJP-03).
2.2.DNA extraction and sequences of ND2Total genomic DNA was extracted from the liver tissue of every specimen collected in this study using the QIAamp DNA Mini Kit(QIAGEN,Hilden,Germany) following the manufacturer’s recommended protocols.The mitochondrial gene NADH dehydrogenase subunit 2 (ND2) was amplified and sequenced.The primer sequences (Jap_70F: CCACCAAACAACTA CACCTA,Jap_1559R: GGATTAATGCCCTCTGGATT)were retrieved from Wanget al.(2019).The PCR amplifications were performed in a 30 μL reaction volume with the following conditions: an initial denaturing step at 95 °C for 4 min;36 cycles of denaturing at 95 °C for 40 s,annealing at 55 °C and extending at 55°C for 70 s,and a final extension step of 72 °C for 10 min.PCR products were sequenced with both forward and reverse primers as used in PCR.Sequencing was conducted using an ABI3730 automated DNA sequencer at Sangon Biotechnologies Co.,Ltd.(Shanghai,China).New sequences were uploaded to GenBank (see Table 1).
2.3.Molecular phylogenetic analysesThere were 21 novel sequences ofND2obtained in this work,and a total of 163ND2sequences of 33Diplodermaspecies were downloaded from GenBank for phylogenetic analyses (Table 1).In addition,corresponding sequences ofAcanthosaura lepidogaster(AF128499),Calotes versicolor(KC875820) andPseudocalotes kakhienensis(MK001435) were downloaded and used as outgroups according to Wanget al.(2022).
Sequences were assembled and aligned using BioEdit v.7.0.9.0 (Hall 1999) with default settings and were further revised manually if necessary.Non-sequenced fragments were treated as missing data.Phylogenetic analyses were conducted using Bayesian inference (BI) and maximum likelihood (ML)methods implemented in MrBayes v.3.2 (Ronquistet al.,2012)and PhyML v.3.0 (Guindonet al.,2010),respectively.Before the phylogenetic analyses,the best evolutionary model was conducted and chosen under the Bayesian inference criteria(BIC) using jModelTest v.2.1.3 (Darribaet al.,2012).The analyses selected the GTR+F +I+G4 model for the mitochondrial gene.In the BI analyses,the parameters for each partition were unlinked,and branch lengths were allowed to vary proportionately across partitions.Two runs each with four Markov chains were simultaneously run for 80 million generations with sampling every 1000 generations.The first 25% of trees were removed as the “burn-in” stage,which was followed by calculations of Bayesian posterior probabilities at stationarity,and the 50% majority-rule consensus of the postburn-in trees sampled was obtained.For the ML tree,branch supports were drawn from 10000 nonparametric bootstrap replicates.Finally,uncorrected genetic pairwise distances (p-distances) were obtained for the coding region ofND2using MEGA 6.0.(Tamuraet al.,2013).
2.4.Morphological analysesMeasurements were taken mainly following Zhaoet al.(1999) to the nearest 1 mm using a steel ruler for snout-vent length (SVL) and tail length(TAL),and to nearest 0.1 mm using a digital calliper for other relatively short measurements.Totally,seven measurements were measured,snout-vent length (SVL): distance from the snout tip to anterior edge of the vent;tail length (TAL): distance from the anterior edge of the vent to the tip of tail;head length(HL): distance from the tip of snout to the right angle of the jaw;snout-eye length (SEL): distance from the tip of snout to the anterior margin of the eye;foreleg length (FLL): distance from the armpit to the tip of finger IV,excluding the claw,measured as the limb straightened;hindleg length (HLL): distance from the groin to the tip of toe IV,excluding the claw,measured with the limb straightened;trunk length (TRL): distance from the armpit to the groin.
Definitions of morphological characters and the counting methods also mainly followed Zhaoet al.(1999) and Wanget al.(2021) as follows,supralabial scale count (SL): number of enlarged,modified labial scales from rostral to the corner of the mouth;infralabial scale count (IL): number of enlarged,modified labial scales from mental to the corner of the mouth;suborbital scale rows (SOR): number of longitudinal rows of scales between supralabials and inferior-most edge of orbit circle,excluding fine ciliary scales in the orbit;middorsal crest scale count (MD): number of modified crest scales longitudinally from the first nuchal crest to the scale above the cloaca;finger IV subdigital lamellae count (F4S): number of subdigital lamellae scale from the base between finger III and IV to the tip of finger IV,excluding the claw;toe IV subdigital lamellae count (T4S): number of subdigital lamellae scales from the base between toe III and IV to the tip of toe IV,excluding the claw;radial stripes below eyes (RSBE): absent or present;gular fold state (GF): absent or present;gulat pouch state (GP): absent or present;gular spot state (GS): absent or present;gular spot color (GSC);tympanum state (TS): absent or present;skin fold under nuchal crest (SFNC): absent or present;nuchal crest state(NC): crest scale strongly erected or not;skin fold under dorsal crest (SFDC): absent or present;shape of dorsolateral stripes(SDS): defined as the shape of dorsolateral stripe in males,either smooth edged or jagged;ventral scale state (VSS): absent or present;hindlimbs adpressed forward (HAF): the hindlimbs adpressed forward to reaching the area.
Coloration descriptions were using terminology and codes in RGB (red,green,blue) color model,which is widely used in various fields (Ibraheemet al.,2012).Data of coloration and ornamentation were also collected from life specimens both in low light or stressed state,including the following (Wanget al.,2021): inner-lip coloration (ILC);coloration of the oral cavity(CO): defined as the background coloration of the anterior roof and sides of the mouth,excluding the posterior palate and deep throat;coloration of the tongue (CTG): defined as the coloration of the tongue;coloration of the dorsolateral stripes(CDS): defined as the background coloration of the dorsolateral stripes;ventrolateral body coloration (VLBC);spots coloration of ventrolateral body (SCVB);suborbital scale color (SOC): color of scales between supralabials and inferior-most edge of orbit circle,excluding fine ciliary scales in the orbit;coloration of the supralabial (CSL) and ventral body coloration (VBC).
3.Results
3.1.Molecular phylogenetic analysesThe final alignment contain 1032 bp ofND2,and two consistent topologies of phylogenetic trees are obtained from BI and ML analyses,showing that all analyzed species ofDiplodermaare strongly clustered into one monophyletic clade,and further five subclades are recognized (Figure 2).
The newly collected samples from the Shuiluo River and Muli River valleys population,the upper-middle valleys of the Yalong River population,and the Yalong River middle valley and Liqiu River valley population are clustered into three distinct lineages,respectively,and are well supported.Specifically,the Shuiluo River and Muli River valleys population is sister toD.yulongense,the upper-middle valleys of the Yalong River population is sister toD.panchi,and the Yalong River middle valley and Liqiu River valley population is sister toD.bowoense.
Because all the three newly recognized lineages belong to the subclade I,genetic distance is just calculated among those species within this subclade.The uncorrected mean genetic distance range is 2.5%–19.6%,while the genetic distance between the upper-middle valleys of the Yalong River population and other congeners is at least 4.3% (the upper-middle valleys of the Yalong River population versusD.panchi),that between the Shuiluo River and Muli River valleys population and other congeners is at least 3.4% (the Shuiluo River and Muli River valleys population versusD.yulongense),and that between the Yalong River middle valley and Liqiu River valley population and other congeners is at least 5.5% (the Yalong River middle valley and Liqiu River valley population versusD.bowoense),which are much higher than the shortest genetic distance ofDiplodermaspecies (2.5% betweenD.velaandD.drukdaypo(Table 2).
Moreover,the uncorrected mean genetic distance among samples of the upper-middle valleys of the Yalong River population is 1.4% (range from 1.1% to 1.7%),that among the Shuiluo River and Muli River valleys population samples is approximately 0.5% (range from 0.3%–0.7%),and that among the Yalong River middle valley and Liqiu River valley population is 0.4% (range from 0.2% to 0.6%).
The results of molecular phylogenetic analyses above indicate that the genetic differentiations among the three lineages newly recognized in this work and others have reach specific level.

Figure 2 Phylogenetic tree of the genus Diploderma inferred from the mitochondrial ND2,Bayesian posterior probability/ML bootstrap support was denoted above node.
3.2.Morphological analysesBased on their measurements,the Shuiluo River and Muli River valleys population,the uppermiddle valley of the Yalong River population,and the Yalong River middle valley and Liqiu River valley population have their own unique morphological characteristics (Figure 3,Table 3).These three populations showed different morphological characteristics from nearby species (Table 4),similar to the results of molecular phylogenetic analyses above.
The upper-middle valleys of the Yalong River population has longer tail (TAL/SVL 206.8%–251.6% [average 226.8%] in males,and 169.5%–208.5% [average 189.7%] in females),T4S 14–18(average 16) and F4S 20–25 (average 23),cyan or dark olive green gular spots present in both sexes in life,fourth toe with claw reaching eye when hindlimbs adpressed forward.It can be differentiated from nearby speciesD.yulongenseby a suite of morphological characters,especially gular coloration and ornamentation in both sexes: light yellow or white ventrolateral body versus pale green to chartreuse in males;white to grey or light yellow spots present on each side of ventrolateral versus pale greenish yellow and chartreuse,and cyan or dark olive green to yellow gular spots present versus chartreuse to opaline green in both sexes in life.
The upper-middle valleys of the Yalong River population has shorter tails (TAL/SVL 146.7%–152.1% (average 149.4%) in males,and 144.8%–146.6% (average 145.9%) in females);T4S 16–21(average 19) and F4S 13–16 (average 14);radial stripes below eyes present;gular spot always cyan surrounded by yellow in live males,and cyan or light cyan surrounded by white in live females.It can be differentiated from nearby species ofD.panchiby a suite of morphological characters: T4S 16–21 (average 19)versus 20–24 (average 22);gular spot cyan versus distinct gular spots absent but mosaic light sulphur yellow patterns present in females.
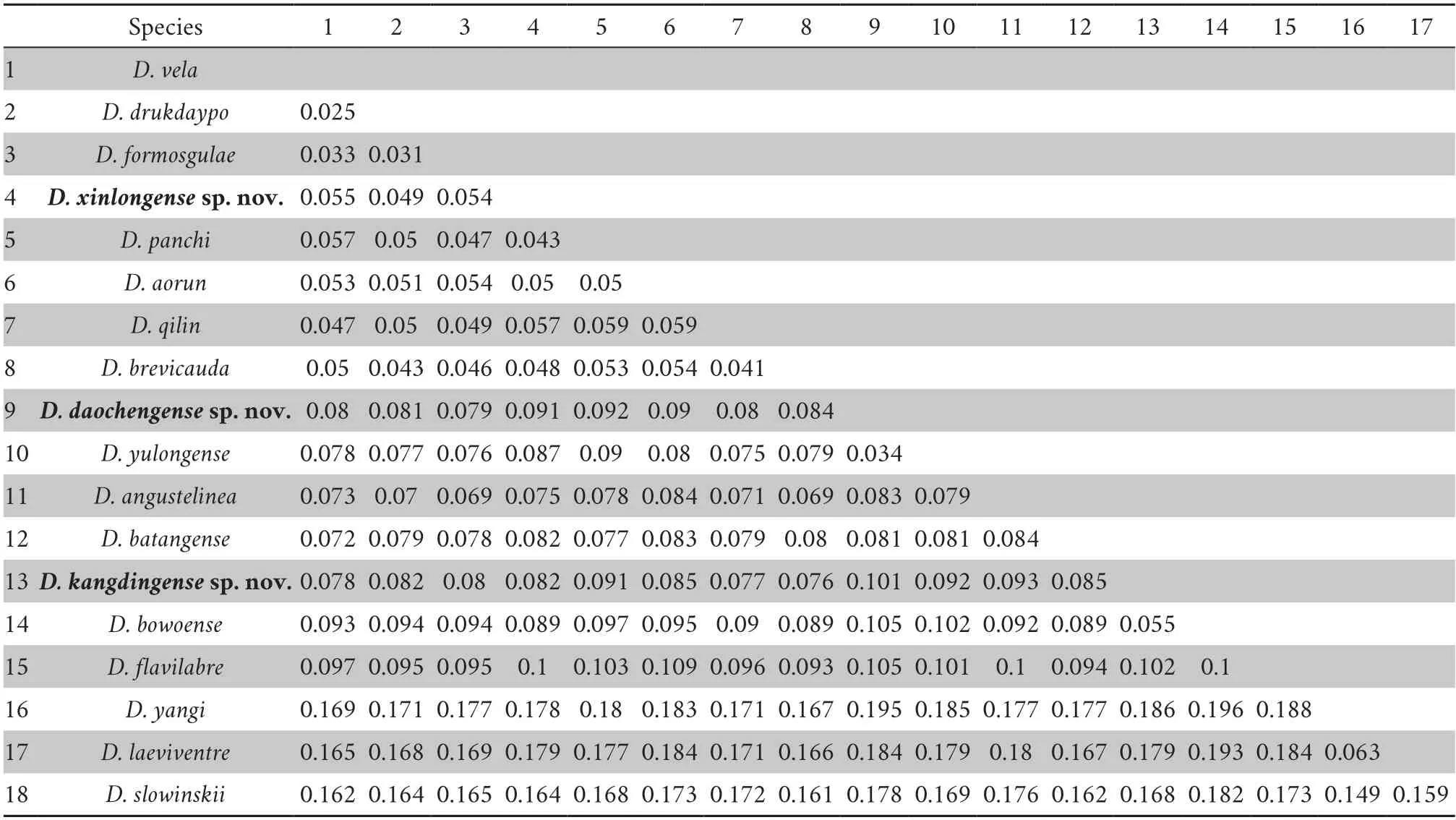
Table 2 Uncorrected genetic distances of the sampled ND2 coding region (1032bp) among the three new lineages (new species) and other species within the sub-clade I in the genus Diploderma (Figure 2).
The Yalong River middle valley and Liqiu River valley populations have longer tails (TAL/SVL 204.8%–227.1% [average 213.2%] in males,and 156.1%–192.5% [average 181.2%] in females);T4S 20–25 (average 23) and F4S 16–20 (average 17);radial stripes below eyes absent;ventrolateral body coloration yellow to light yellow,with yellow or white spots in males,light grey to white smoke with white or wheat spots in females.It can be differentiated from nearby species ofD.bowoenseby a suite of morphological characters: longer trunk length in males (TRL≥ 46.8% SVL versus ≤ 45.3%);ventrolateral body yellow to light yellow,with yellow or white spots in males versus light grey with white spots in males;yellow gular spot present in males versus light chrome orange gular spots present in males and dark spectrum yellow in females.
Both the molecular phylogenetic relationships and the uncorrected pairwise distance inferred fromND2,and the integration of the differences from morphological data,suggest that the three new lineages from the three populations collected from the aforementioned localities represent three new species,they are described here.
3.3.Taxonomic accounts
3.3.1. Diploderma daochengense sp.nov.(Tables 3–4;Figure 3)ChresonymsJapalura flavicepsZhaoet al.,1999: 111–115 in part.;Diploderma yulongenseShuet al.,2021: 259–264.
Suggested English name:Daocheng Mountain Lizard
Suggested Chinese name:稻城攀蜥(dào chéng pān xī)
HolotypeAdult male,CIB119352 (filed number DC001),collected from Eyatong Township (28.04754478°N,100.3224385°E;at elevation 1916 m a.s.l.),Daocheng County,Ganzi Tibetan Autonomous Prefecture,Sichuan Province,China,collected by Huaming ZHOU and Jing LI on September 2021.
AllotypeAdult female,CIB119353 (filed number DC003),also collected from Eyatong Ethnic Township (28.0469772°N,100.32392371°E;at elevation 1930 m a.s.l.),and the same collecting information as the holotype.

Figure 3 Display of the three new species and their habitats.1: Diploderma daochengense sp.nov.,2: D.xinlongense sp.nov.,3.D.kangdingense sp.nov.;A: dorsolateral overview of holotype,B: ventral head view of holotype,C: dorsolateral overview of allotype,D:ventral head view of allotype;E: habitats (A1,B1,C1 and D1 photographed by Bo CAI;A2,B2,C2,D2,A3,B3,C3 and D3 by Meihua ZHANG and Jianping JIANG;E1,E2 and E3 by Huaming ZHOU).
Other specimens (Pratypes)The specimen DC004 (♀,28.04838729°N,100.3217799°E;at elevation 1925 m a.s.l.) collected from Eyatong Ethnic Township.The specimens 20210904(♀,27.946867°N,100.435083°E;at elevation 1664 m a.s.l.),20210905 (♂,27.953308°N,100.427531°E;at elevation 1652 m a.s.l.) collected from Eya Naxi Ethnic Township,Muli Tibetan Autonomous County,Liangshan Yi Autonomous Prefecture,Sichuan Province,China,by Huaming ZHOU and Jing LI on September 2021;2019ML0037 (♂) and 2019ML0038 (♀)collected from Xiamaidi Township,Muli Tibetan Autonomous County (27.789794°N,101.200958°E;at elevation 1889 m a.s.l.) by Feng XIE,Puyang ZHENG and Mingyang CHENG on August 2019;2019ML0006 (subadult,28.283388°N,100.661666°E;at elevation 2208 m a.s.l.) and 2019ML0012 (subadult,28.287653°N,100.654886°E;at elevation 2158 m a.s.l.) collected from Shuiluo Township,Muli Tibetan Autonomous County by Feng XIE,Puyang ZHENG and Mingyang CHENG on August 2019;ML070706 (♀,28.287653°N,100.654886°E;at elevation 2158 m a.s.l.) and ML070807 (♀,28.290626°N,100.666664°E;at elevation 2180 m a.s.l.) respectively collected from Shuiluo Township by Feng XIE,Puyang ZHENG and Mingyang CHENG on July 2020.
EtymologyThe specific epithet of “Daocheng” refers to a county name of Sichuan,and the new species is found on its southeastern edge.
DiagnosisDiploderma daochengensesp.nov.can be distinguished from otherDiplodermaspecies by the following characters: 1) TAL/SVL 206.8%–251.6% (average 226.8%) in males,and 169.5%–208.5% (average 189.7%) in females;2) TRL/SVL 40.8%–67.3% in males and 45.6%–70.3% in females;3) SEL/HL 39.6%–71.2% in males and 37.5%–68.8% in females;4) T4S 14–18 (average 16) and F4S 20–25 (average 23);5) no bright transverse stripes or bands on dorsal body;6) ventral scales distinctively keeled;7) tympanum concealed;8) transverse gular fold deep,forming a distinct pouch across throat;9) no radial stripes around eyes;10) inner-lip coloration smoky white,and coloration of tongue and oral cavity light flesh color;11)ventrolateral body white to grey or light yellow with yellow,cyan or white spots in males;12) cyan or dark olive green gular spots present in both sexes in life;13) upper edge of the dorsolateral stripes smooth,and yellow (normally),brown(when dimly lit or stressed) in males,gray while white or light yellow in females in life;14) fourth toe with claw reaching eye when hindlimbs adpressed forward;15) well-developed skin folds under nuchal crest present in males only,vertebral crest discontinuous between nuchal and dorsal sections with a distinct gap.

Table 3 Morphological comparisons among the three new lineages (new species) in the genus Diploderma.
Description of holotypeAdult male,body medium in size,SVL 57.6 mm;tail long,TAL 144.9 mm;head longer (18.3 mm)than width 12.6 mm;head depth 10.1 mm;snout-eye length 7.3 mm;foreleg length 23.2 mm;hindleg length 42.3 mm;toe IV length 12.1mm;trunk length 27.8 mm;TAL/SVL 251.6%;FLL/SVL 40.3%;HLL/SVL 73.4%;TRL/SVL 48.2%;SEL/HL 39.6%;HL/SVL 31.8%.

It has 10/8 supralabial scales,10/11 infralabial scales,and 40 middorsal crest scales;finger IV subdigital lamellae 16/16;toe IV subdigital lamellae 24/22;nasal-supralabial scale rows 2/2;suborbital scale rows 3/3;eight stripes around the eye except the subocular regions,including one enlarged stripe extending from the posterior edge of the orbit to the corner of the mouth.Transverse gular fold present,distinct;gular pouch present and distinct in life;gular spot present and grey lines on gular region;tympanum covered with scales;well-developed skin fold under nuchal crest present,vertebral crest discontinuous between nuchal and dorsal sections with gap;scales of ventral,head and body distinctively keeled;the fourth toe with claw reaching the eye not beyond it when hindlimbs adpressed forward.The lower edges of the dorsal stripes strongly serrated;the upper edge is smooth.
Coloration of holotypeThe background of the dorsal and lateral surfaces of the head is iron grey;two dorsal head stripes present above the eyes;subocular and supralabial are smoky white,the middle area is a light grey horizontal short stripes;infralabial is white.Inner-lip is smoky white,oral cavity and tongue are light flesh color.Gular region has cyan gular spot and grey lines;nasal is consistent with surrounding as light grey.Seven dorsal patches present,iron grey;of dorsolateral stripes is yellow (normally),brown (dimly lit or stressed);ventrolateral body is grey with white spots;ventral body is white (normally),antique white with light grey reticulates (dimly lit or stressed);interfemoral color is the same as ventral body in life.
Comparisons Diploderma daochengense sp.nov.is the sister species ofD.yulongense,they are morphologically similar,but has different characters,such as the following: 1) SEL/HL 39.6%–71.2% versus 36.8%–39.0% in males,and 37.5%–68.8% versus 36.7%–38.2% in females;2) the upper edge of the dorsolateral stripes smooth versus strongly jagged in males;3) ventrolateral body light yellow or white versus pale green to chartreuse in males;4) white to grey or light yellow spots present on each side of ventrolateral versus pale greenish yellow and chartreuse,and 5) gular spots cyan or dark olive green to yellow versus chartreuse to opaline green in both sexes in life.
The new species differs from other congeners of the genusDiplodermaas follows: differs fromD.fasciatum and D.zszechwanensisby having the following combination of characters: 1) slightly square green patches non-existent on mid dorsum,2) males having dorsolateral stripes and dorsolateral series of spots;differs fromD.varcoae,D.swild,D.slowinskii,D.panlongandD.dymondiby havingconcealed tympanum(Wanget al.,2020);differs fromD.grahami,D.micangshanense,D.yunnanense,D.chapaense,D.swinhonis,D.makii,D.luei,D.brevipes,D.polygonatum,D.ngoclinense,D.splendidum,D.hamptoniandD.menghaienseby having deep transverse gular fold and forming a distinct pouch across throat;differs fromD.yangi,D.panlong,D.dymondi,D.swild,D.angustelineaandD.iadinumby having a dorsolateral stripe shape strongly jagged versus smooth or feebly jagged;differs fromD.laeviventreandD.drukdaypoby having distinctively keeled ventral scales;differs fromD.drukdaypo,D.flaviceps,D.vela,D.panchiandD.kangdingensesp.nov.by having gular spots in both sexes;differs fromD.brevicaudaby having: 1) inner-lip smoky white versus sulphur yellow;2) the tongue light flesh versus light orange yellow;differs fromD.aorun,D.qilin,D.flavilabre,D.zhaoermii,D.batangense,D.formosgulae,D.bowoense,D.brevicauda,D.angustelinea,D.kangdingensesp.nov.andD.xinlongensesp.nov.byhaving white to grey or light yellow ventrolateral body in live males,dark olive green or cyan gular spots in both sexes,and no dark radial stripes around eyes.
DistributionAt present,the new species is only known to be distributed in 4 locations in Sichuan Province,China: Eyatong Township in Daocheng County,Sichuan Province;Eya Naxi Ethnic Township,Xiamaidi Township and Shuiluo Township in Muli Tibetan Autonomous County.The area between the distribution points exceeds 2300 km2and the farthest straight-line distance between known distribution points is approximately 91 km.The known altitude range of this species is 1652–2608 m.This new species inhabits the deciduous broadleaved forest along the Shuiluo River and Muli River and is active at the edge of forests,in bushes,and sometimes even on roads.
3.3.2.Diploderma xinlongense sp.nov.(Tables 3–4;Figure 3)
Suggested English name:Xinlong Mountain Lizard
Suggested Chinese name:新龙攀蜥 (xīn lóng pān xī)
HolotypeAdult male,CIB119354 (filed number 20210907),collected from Eri village (30.8954°N,100.231797°E;at elevation 3040 m a.s.l.),Wuya Township,Xinlong County,Ganzi Tibetan Autonomous Prefecture,Sichuan Province,China,collected by Huaming ZHOU and Jing LI on September 2021.
AllotypeAdult female,CIB119355 (filed number 20210906),also collected from Eri vallige (30.895089°N,100.230106°E;at elevation 3057 m a.s.l.),Wuya Township,Xinlong County,Ganzi Tibetan Autonomous Prefecture,Sichuan Province,China,by Huaming ZHOU and Jing LI.
EtymologyThe specific epithet of “Xinlong” refers to the Xinlong County where the new species was discovered.
Other specimens (Paratypes)The specimens 20210908 share the same locality as the holotype,the 20210909 (♀,30.525361°N,100.339067°E;at elevation 2961 m a.s.l.),20210910(subadult,30.526836°N,100.337386°E;at elevation 2976 m a.s.l.),and 20210911(♂,30.528414°N,100.3357°E;at elevation 2951 m a.s.l.) were collected from Junba Brindge,Xinlong County;20210912 (♂,30.895036°N,100.228056°E;at elevation 3052 m a.s.l.) collected from Wuya Township,Xinlong County.All the specimens were from Xinlong County,Ganzi Tibetan Autonomous Prefecture,China,and the collector information as the holotype.
Diagnosis Diploderma xinlongense sp.nov.can be mainly distinguished from otherDiplodermaspecies by the following characters: 1) TAL/SVL 146.7%–152.1% (average 149.4%) in males,and 144.8%–146.6% (average 145.9%) in females;2) TRL/SVL 43.4%–45.1% in males and 45.3%–52.6% in females;3) HL/SVL 31.2%–31.8% in males and 29.4%–34.2% in females;4) T4S 16–21 (average 19) and F4S 13–16 (average 14);5) ventral scales distinctively keeled;6) tympanum concealed;7) transverse gular fold deep,forming a pouch across throat;8) radial stripes below eyes present;9) ventrolateral body smoky white to white (one specimen is mosaic light yellow),with light yellow and white spots in males,light grey white or wheat spots in females;10)gular spot always cyan surrounded by yellow in live males while cyan or light cyan surrounded by white in live females;11) inner-lip smoky white,and tongue and oral cavity light flesh color;12) dorsolateral stripes strongly jagged,and yellow in males while sandy brown (normally) or light grey (dimly lit or stressed) in live females;13) skin fold under dorsal crest and nuchal crest are weak;14) fourth toe with claw reaching tympanum when hindlimbs adpressed forward.
Description of holotypeAdult male,body small in size,SVL 45 mm;tail short,TAL 66 mm;head longer 1 (4.3 mm) than width 9.3 mm;head depth 6.6 mm;snout-eye length 5.6 mm;foreleg length 17.8 mm;hindleg length 29.0 mm;toe IV length 7.5 mm;trunk length 19.5 mm;TAL/SVL 146.7%;FLL/SVL 39.5%;HLL/SVL 64.4%;TRL/SVL 43.4%;SEL/HL 39.1%;HL/SVL 31.7%.
It has seven supralabial scales on each side,9/10 infralabial scales,and 38 middorsal crest scales;finger IV subdigital lamellae 14 on each side;toe IV subdigital lamellae 16/17;nasalsupralabial scale rows 1/1;suborbital scale rows 3/3;more than eleven radiated black stripes around the eye,including one enlarged stripe extending from the posterior edge of orbit to the corner of mouth and the anterior edge of tympanic;dorsal head has three stripes;transverse gular fold present,distinct;gular pouch present in life,but indistinct after preservation;gular spot present and dark,short lines on gular region;tympanum covered with scales;skin fold under nuchal crest absent;nuchal crest absent;skin fold under dorsal crest absent;scales of ventral,head and body distinctively keeled;the fourth toe with claw reaching the tympanum not to the jaw joint when hindlimbs adpressed forward;the upper and lower edges of the dorsal stripes strongly serrated.
Coloration of holotypeThe background of the dorsal and lateral surfaces of the head is medium grey;dorsal head having three dark grey stripes;inner-lip is smoky white,oral cavity and tongue are light flesh color;gular spot color is cyan surrounded by yellow in life;nasal coloration is consistent with surrounding as light grey;dorsal patches present,of which the shoulder and caudal base are independent patches,but on the central trunk are joined together.Dorsolateral stripes are yellow;ventrolateral body is white with light yellow spots;ventral body is white,in a dark or stressful environment,there will be a few scattered black spots;interfemoral is unobvious light cyan in life.
ComparisonsDiploderma xinlongensesp.nov.is morphologically similar toD.aorun,but it has some features that can distinguish it from the latter: 1) skin fold under dorsal crest and nuchal crest in males weak versus strongly;2) crest scale not strongly erected versus strongly in both sexes;3)gular spot in males always cyan surrounded by yellow versus cyan surrounded by white;4) TAL/SVL 146.7%–152.1% versus 212.0%–221.1% in males,and 144.8%–146.6% versus 191.4%–207.7%in females.
Diploderma xinlongensesp.nov.is the sister species ofD.panchi,but it can be distinguished from it mainly by the following characters: 1) MD in females 36–41 (average 38) versus 42–46 (average 45);2),T4S 16–21 (average 19) versus 20–24(average 22);3) HL/SVL is 29.4%–34.2% versus 27.0%–28.8% in females;4) gular spot cyan versus distinct gular spots absent but mosaic light sulphur yellow patterns in females.
For other species,Diplodermaxinlongensesp.nov.differs fromD.fasciatum and D.zszechwanensisby having dorsolateral stripes and dorsolateral series of spots in males versus large,slightly square green patch on the mid-dorsum.It differs fromD.varcoae,D.swild,D.slowinskii,D.panlong,D.dymondiby havingconcealed tympanum;differs fromD.grahami,D.micangshanense,D.yunnanense,D.chapaense,D.swinhonis,D.makii,D.luei,D.brevipes,D.polygonatum,D.ngoclinense,D.splendidum,D.hamptoniandD.menghaienseby having a deep transverse gular fold;differs fromD.laeviventreandD.drukdaypoby having distinctively keeled ventral scales;differs fromD.laeviventre,D.panlong,D.dymondi,D.swild,D.yangi,D.
angustelineaandD.iadinumby having a dorsolateral stripe shape strongly jagged versus smooth or feebly jagged;differs fromD.dymondi,D.micangshanense,D.slowinskii,D.swild,D.flaviceps,D.vela,D.drukdaypo,D.varcoae,andD.panlongby having gular spots present in males,differs fromD.kangdingensesp.nov.,D.panchiandD.zhaoermiibyhaving gular spots in females;differs fromD.aorun,D.qilin,D.flavilabre,D.zhaoermii,D.batangense,D.formosgulaeD.bowoense,D.bowoense,D.daochengensesp.nov.andD.kangdingensesp.nov.byhaving always cyan gular spot surrounded by yellow in males while the cyan in females in life.
DistributionTo date,the new species is only known to be distributed in the upper-middle Yalong River,from Eri Village to the area around the Junba Bridge in Xinlong County,Sichuan Province,China.The known altitude range of this species is 2950–3080 m,and the farthest straight-line distance between known distribution points is approximately 43 km.This new species inhabits the dry valley along the upper-middle Yalong River,and is active at the edge of coniferous forests,bushes or rock piles in the valley.
3.3.3.Diploderma kangdingensesp.nov.(Tables 3–4;Figure 3)ChresonymsJapalura flavicepsZhaoet al.,1999: 111–115 in part;Zhao,2003: 84 in part;Caiet al.,2018 in part.
Suggested English name:Kangding Mountain Lizard
Suggested Chinese name:康定攀蜥 (kāng dìng pān xī)
HolotypeAdult male,CIB119356 (filed number 20210917),collected from Pusharong Village (29.417275°N,101.159856°E;at elevation 2409 m a.s.l.),Pusharong Township,Kangding City,Ganzi Tibetan Autonomous Prefecture,Sichuan Province,China,collected by Huaming ZHOU and Jing LI on August,2021.
AllotypeAdult female,CIB119357 (filed number 20210916),also collected from Pusharong Village (29.417272°N,101.165431°E;at elevation 2432m a.s.l.),Pusharong Township,Kangding City,Ganzi Tibetan Autonomous Prefecture,Sichuan Province,China,collected by Huaming ZHOU and Jing LI on August,2021.
EtymologyKangding City is the capital of Ganzi Tibetan Autonomous Prefecture,which is located in the east of Ganzi in Sichuan Province.The specific epithet refers to the Kangding City where the new species was discovered.
Other specimens (Paratypes)The specimens 20210913 (♀,29.420342 N,101.144353 E;at elevation 2421 m a.s.l.),20210914(subadult,share the same locality as 20210913) and 20210915(♀,29.419633 N,101.143456 E;at elevation 2430m a.s.l.) were collected from Bosihe Town of Yajiang County by Huaming ZHOU and Jing LI on August,2021;KD2021080101 (♂),KD2021080102 (♂) and KD2021080104 (♂),KD2021080103(♀),KD2021080105(♀) collected from Pusharong Village(29.419742 N,101.153925 E;at elevation 2430 m a.s.l.),and the collecting information same as the holotype.
DiagnosisDiploderma kangdingensesp.nov.can be mainly distinguished from otherDiplodermaspecies by the following features: 1) TAL/SVL 204.8%–227.1% (average 213.2%) in males,and 156.1%–192.5% (average 181.2%) in females;2) TRL/SVL 46.8%–48.6% in males and 50.1%–56.8% in females;3) T4S 20–25(average 23) and F4S 16–20 (average 17);4) bright transverse stripes on dorsal body,especially in males in life;5) ventral scales distinctively keeled;6) tympanum concealed;7) transverse gular fold deep in both sexes,and males having a distinct pouch across throat;8) inner-lip smoky white,and tongue and oral cavity light flesh color;9) dark stripe patterns on the ventral head;10)gular spot present in males (yellow),and absent in females;11)dorsolateral stripes strongly jagged,and yellow in males while front yellow,rear grey or sandy brown in females;12) radial stripes below eyes absent;13) ventrolateral body yellow to light yellow,with yellow or white spots in males,light grey to smoky white with white or wheat spots in females;14) fourth toe with claw reaching eye when hindlimbs adpressed forward;15)well-developed skin folds under nuchal crest present in males only,vertebral crest discontinuous between nuchal and dorsal sections with gap.
Description of holotypeAdult male,body medium in size,SVL 54 mm;tail long,TAL 113 mm;head longer (17.5 mm) than width 11.7 mm,head depth 9.2 mm;snout-eye length 7.4 mm;foreleg length 25.3 mm;hindleg length 42.0 mm;toe IV length 10.81 mm;trunk length 25.25 mm;TAL/SVL 209.3%;FLL/SVL 46.9%;HLL/SVL 77.8%;TRL/SVL 46.8%;SEL/HL 42.3%;HL/SVL 32.4%.
It has 8/9 supralabial scales,9/10 infralabial scales,and 35 middorsal crest scales;finger IV subdigital lamellae 20/18;toe IV subdigital lamellae 25/25;nasal-supralabials scale rows 1/1;suborbital scale rows 3/3;three stripes around the eye except the subocular regions,including one enlarged stripe extending from the posterior edge of orbit to the corner of mouth and the anterior edge of tympanic,one stripes extended from the postsub-orbit to the occiput,and one stripes extended from the preocular to nasal.Transverse gular fold present,distinct;gular pouch present and distinct in life;gular spot present and grey lines on gular region;tympanum covered with scales;skin fold under nuchal crest absent;nuchal crest present;skin fold under dorsal crest present;scales of ventral,head and body distinctively keeled;fourth toe with claw reaching eye not beyond it when hindlimbs adpressed forward;the upper and lower edges of the dorsal stripes strongly serrated;the lower edge stronger.
Coloration of holotypeThe background of the dorsal and lateral surfaces of the head is seal brown;dorsal head stripes are unobvious;subocular and supralabial are smoky white,the middle area is a light grey horizontal short stripe;infralabial is white.Inner-lip is smoky white,and the oral cavity and tongue are light flesh color.Gular region has yellow spots and grey lines;nasal coloration is consistent with surrounding smoky white.Dorsal patches present,but connected and corrugated;dorsolateral stripes is yellow (normally),brown (dimly lit or stressed);ventrolateral body is yellow with white spots;ventral body is white,and light grey reticulates appear in low light or stressed state;interfemoral color is the same as ventral body in life.
ComparisonsDiploderma kangdingensesp.nov.is morphologically similar toD.qilin,but it has some features that can be distinguished from the latter: 1) presence of dark stripe patterns on the ventral head versus grey reticulated patterns;2) gular having yellow spot in males versus gular having light sulphur yellow spots in both sexes;3) dorsolateral stripes yellow and sandy brown in males,yellow and grey in females versus cream yellow in males,beige in females;4) radial stripes below eyes are absent versus present.
Diploderma kangdingensesp.nov.is the sister species ofD.bowoense,but it can be distinguished from it by the following characters: 1) longer trunk length in males (TRL ≥ 46.8% SVL versus ≤ 45.3%);2) more T4S 20–25 (average 23) versus 18–21(average 20);3) more F4S 16–20 (average 17) versus 14–17(average 15);4) ventrolateral body yellow to light yellow,with yellow or white spots in males versus light grey with white spots in males;5) gular having yellow spot in males versus gular having light chrome orange spots present in males and dark spectrum yellow spots in females;6) dorsolateral stripes yellow and sandy brown in males,yellow and grey in females versus cream in males,sulphur yellow in females.
For other species,Diplodermakangdingensesp.nov.differs fromD.fasciatum and D.zszechwanensisby having dorsolateral stripes and dorsolateral series of spots in males versus large,slightly square green patch on the mid-dorsum.It differs fromD.varcoae,D.swild,D.slowinskii,D.panlong,andD.dymondiby havingconcealed tympanum;differs fromD.grahami,D.micangshanense,D.yunnanense,D.chapaense,D.swinhonis,D.makii,D.luei,D.brevipes,D.polygonatum,D.ngoclinense,D.splendidum,D.hamptoniandD.menghaienseby having deep transverse gular fold and forming a distinct pouch across throat;differs fromD.laeviventreandD.drukdaypoby having distinctively keeled ventral scales;differs fromD.laeviventre,D.panlong,D.dymondi,D.swild,D.yangi,D.angustelinea andD.iadinumby having a dorsolateral stripe shape strongly jagged versus smooth or feebly jagged;differs fromD.panlong,D.swild,D.micangshanense,D.slowinskii,D.swild,D.flaviceps,D.vela,andD.drukdaypoby having gular spots in males;differs fromD.panchiby having no typical radial stripes below eyes,no light sulphur yellow mosaic gular pattern;longer tail length (in females TAL ≥ 156.1% SVL versus ≤ 151.5%.);differs fromD.brevicaudaby having 1) inner-lip smoky white versus sulphur yellow;2)the tongue light flesh color versus light orange yellow;and differs fromD.daochengensesp.nov.,D.xinlongensesp.nov.,D.menghaiense,D.yangi,D.aorun,D.batangense,D.angustelinea,D.qilin,D.iadinum,D.laeviventre,D.flavilabre,andD.yulongenseby having gular spots absent in females;differs fromD.aorun,D.qilin,D.flavilabre,D.zhaoermii,D.batangense,D.formosgulaeandD.bowoensebyhaving yellow ventrolateral and yellow gular spots present in live males,and no dark radial stripes around eyes in both sexes.
DistributionAt present,the new species is only known from the type locality near the middle Yalong River and Liqiu River,a tributary of the Yalong River,in Sichuan Province,China.The known altitude range of this species is 2409–2432 m.This new species inhabits the warm-dry valley along the middle Yalong River and active at the edge of coniferous and broadleaved forest,in bushes or on rock piles in the valley.
4.Discussion
In terms of morphology,the differences between these three new species and otherDiplodermaspecies are mainly reflected in the characters: shape and color of dorsolateral stripe,gular spot coloration,radial stripe state below eyes,ventrolateral body and ventral coloration,nuchal crest state (Wanget al.,2020),and the reaching area of hindlimbs adpressed forward (Zhaoet al.,1999).Coloration patterns and ornamentation in reptiles have taxonomic significance (Wanget al.,2020),especially in the family Agamidae.Our findings further confirm this view.At present,there are a variety of color models to describe color with different color names and parameters.By focusing on the most widely used color model internationally,taxonomic researchers would benefit comparative and interdisciplinary research.The RGB (red,green,blue) color model is a threechannel default mode that has a range of approximately16.78 million colors.It is widely used in various fields,especially in scanning and image processing (Ibraheemet al.,2012),and in spectrophotometers.The RGB colors are easy to query on the computer,and to obtain RGB color cards,and RGB colors easily convert to other color systems.Therefore,we attempted to use this model here.
However,we found that reptile color changes in response to changes in ambient temperature or possibly even stress.Specifically,reptile colors will also vary due to light intensity,audience visual difference,and the parameter setting of the photographs.Therefore,it is hard to accurately describe the color of a species,and there are always some deviations in the color description.Meanwhile,the color of gular spot is not only one type in a species.The gular spot ofDiploderma daochengensesp.nov.has two colors: Cyan or Dark Olive Green,and may appear in both males and females.Moreover,the gular spot contains more than one color in the same individual,such as,the gular spot ofDiploderma xinlongensesp.nov.has two layers of color.Hence,more attention should be given to body color under different temperatures or illumination levels,and body color in the active range should be selected as the basis of species classification in practical studies.In the future,strengthening research on the color variation range of species may promote research on taxonomy and behavioral ecology.
In terms of phylogenetic relationship and geographical distribution,the three new species cluster together with species in the upper Lancang River,the middle and upper Jinsha River,and the middle and upper Yalong River formed the subclade I;the species in the middle and upper reaches of the Nujiang River formed the subclade II;the species in southwestern Yunnan (the lower of the Nujiang River,the lower Lancang River and the Yuanjiang River Valleys) formed the subclade III.The phylogenetic position ofD.iadinum,with a relatively special distribution located in the middle reaches of the Lancang River,formed the subclade IV alone,as the base of the group of subclades IIIIII;those species distributed in the lower Yalong River-Lower Jinsha River,Minjiang River-Qinling Mountains,Dadu River-the middle reaches of the Yangtze River,Taiwan island formed the subclade V,being the sister group to the that of the subclades IIIIIIIV.This distribution patterns is similar to that of Wanget al.(2020).These relationships indicate a strong correlation between phylogenetic relationships and geographic distribution of the genusDiploderma.
However,the internal phylogenetic relationship of the subclade I does not strictly correspond to the river valley.For example,Diploderma xinlongensesp.nov.andD.panchiin the middle reaches of the Yalong River are in the same branch withD.aorunin the middle and upper reaches of the Jinsha River;D.daochengensesp.nov.are distributed in two adjacent tributaries of the Jinsha River;D.kangdingensesp.nov.andD.xinlongensesp.nov.are distributed upstream of their respective sister species.To some extent,this reflects the complexity of the speciation processes of the Qinghai-Tibet Plateau on shaping their phylogenetic relationships and geographical distribution patterns,and reflects the important role of the Shaluli Mountains in the process of species differentiation of theDiploderma.
Recent studies continue to improve our understandings of the taxonomy and phylogeny of the Mountain Lizard genusDiploderma(Caiet al.,2022;Wanget al.,2022).The discoveries of these three new species in and around the Shaluli Mountains provide new insights into the diversity and the distribution patterns of the genusDiploderma,andverify the hypothesis that there might be additional hidden species within this genus in the Hengduan Mountain Region of China (Caiet al.,2022;Wanget al.,2022),further indicating that the Shaluli Mountains and vicinity are one of the regions with the highest concentration of the Mountain Lizard.The taxonomic discovery further highlights the underestimated diversity of the genus and the importance of habitat conservation of the neglected hot-dry valley ecosystems in the Hengduan Mountain Region of China,especially in the Jinsha River and Yalong River valleys.
AcknowledgementsWe thank Feirong JI,Ayi BUDIAN,Peng YAN,Hui LI,Guocheng SHU and Puyang ZHENG for their help in the laboratory work,thank Jiatang Li for his help in the Museum of Herpetology,Chengdu Institute of Biology (CIB).This work is supported by the Second Tibetan Plateau Scientific Expedition and Research Program (STEP 2019QZKK05010503 and 2019QZKK04020202),the Biodiversity Survey and Assessment Project of the Ministry of Ecology and Environment (2019HJ2096001006),and China Biodiversity Observation Networks (Sino BON);Survey of wild vertebrates in Ganzi Prefecture (20yyjs00030).
Appendix
Table S1Morphological data of the type series ofDiploderma daochengensesp.nov.,D.xinlongensesp.nov.andD.kangdingensesp.nov.All morphometric measurements are in the unit of mm.For measurement methods and abbreviations,see the Methods section.The file can be downloaded from the website https://pan.baidu.com/s/1zUzA48_qxYEj2bq_6Cjtag?pwd=2210 (access code: 2210).
猜你喜欢
杂志排行

Asian Herpetological Research的其它文章
- Desertification Drives the Shift in Egg Size-Number Trade-Off in an Agamid Lizard
- Endocast Morphological Variation and Its Driving Forces in Scutiger boulengeri
- Metagenomic Analysis of Mangshan Pit Viper (Protobothrops mangshanensis) Gut Microbiota Reveals Differences among Wild and Captive Individuals Linked to Hibernating Behaviors
- Lineage Diversification and Niche Evolution in the Chinese Cobra Naja atra(Elapidae)
- Phylogenetic Relationships among Chinese Rice Frogs within the Fejervarya limnocharis Species Complex (Amphibia: Dicroglossidae)
- Application of eDNA Metabarcoding for Detecting Anura in North China