短期增温对川西北高寒草甸植物群落结构和叶片性状的影响
2023-01-03吉使阿微赵文学周青平田莉华
吉使阿微, 赵文学, 肖 颖, 周青平,2, 田莉华,2*
(1.西南民族大学青藏高原研究院, 四川 成都 610041; 2.四川若尔盖高寒湿地生态系统国家野外科学观测研究站,四川 成都 610041)
青藏高原分布有我国面积最大的高寒草地,也是全球生物多样性保护的重点区域,对气候变化响应十分敏感。高寒草甸是青藏高原主要的草地类型,总面积约8.2×107hm2,占青藏高原草地面积的50%[1-2]。气候变暖将对全球生物多样性产生重要影响,会显著改变陆地生态系统的结构和功能[3-4]。近年来,青藏高原气候变化呈变暖变湿趋势,这将显著影响高寒草甸的植被群落结构特征[5-6]。同时,植物叶片大小、比叶面积、叶氮和叶磷含量等性状也会随气候变暖而产生一系列响应,进而影响高寒草甸不同植物的生存策略和竞争能力[7]。
在高寒草甸生态系统中,植物群落会通过自身调节来抵御环境增温对其的影响[8]。植物群落的高度和生物量会随着温度升高而增加[9],但增温幅度过大会降低植物群落盖度及地上生物量[10-13]。不同功能群物种对增温的响应程度不同[14]。增温显著增加杂类草的盖度,降低禾本科和莎草科植物的盖度[5]。然而,也有研究发现增温后禾本科和莎草科植物的生物量显著增加[5],而杂类草植物的生物量有所减少[15]。植物群落盖度、高度、多样性和生物量对增温呈现出不同程度的响应,反映了植物群落中不同物种对空间资源的竞争及养分的获取能力的差异[16-17]。
植物叶片性状作为植物个体的关键性状之一,能直观指示植物对外界环境变化的响应和适应策略[18]。温度对植物的生长发育具有十分重要的作用,而叶片性状会随着温度变化而发生不同程度的改变[19]。研究发现,植物可通过调节叶片大小来适应环境变化[20-21]。在高寒草地生态系统中,增温后矮嵩草(Kobresiahumili)和风毛菊(AussureaJaponica)的叶长显著增加,而藏嵩草(KobresiatibeticaMaxim)和高山嵩草(Kobresiapygmaea)的叶长呈降低趋势[6]。温度升高,植物叶面积会呈增加趋势,进而以提高其光合作用和蒸腾速率。植物叶片较高的比叶面积可以增加其养分保持的能力,较低的比叶面积可以增强其对干旱和贫瘠环境的适应能力[22]。韩文轩等[23]研究发现,叶氮和叶磷含量随着温度升高而增加。然而,关于气候变暖背景下,植物叶氮和叶磷含量的变化趋势,尚无一致的结论[24-25]。
本文以位于青藏高原东缘的川西北高寒草甸为研究对象,采用被动式开顶箱(Passive open-top chamber,OTC)模拟增温,研究短期增温处理对高寒草甸植物群落结构以及7种常见物种叶片性状的影响,以期揭示川西北高寒草甸植物群落和叶片性状对增温的响应规律。
1 材料与方法
1.1 研究区概况
本研究在四川若尔盖高寒湿地生态系统国家野外科学观测研究站(102°58′E,32°83′N,3 504 m)开展。该地区属大陆性高原寒温带半湿润季风气候,春秋短促、冬季漫长、无夏且植被生长季短。年均气温为1.5℃,年均降水量约为749 mm,且全年80%降雨集中在5—9月[15]。主要植被类型为高寒草甸,莎草科和禾本科植物为优势种,杂类草为伴生种,豆科植物较少。主要优势种为四川嵩草(KobresiasetschwanensisHand.-Mazz.)、无脉薹草(CarexenervisC. A. Mey.)和垂穗披碱草(ElymusnutansGriseb.),伴生种为蒲公英(TaraxacummongolicumHand.-Mazz.)、蓝玉簪龙胆(GentianaveitchiorumHemsl.)、小花草玉梅(AnemonerivularisBuch.-Ham. var. flore-minore Maxim.)和鹅绒委陵菜(PotentillaanserinaL.)等[26]。
1.2 试验设计
在2018年5月初,选择若尔盖站内地势平坦且植物群落较为均一的典型高寒草甸作为研究对象。在该试验样地内,采用被动式开顶箱(Passive open-top chamber,OTC)设置了增温处理(W),以不增温处理作为对照(CK)。OTC采用聚碳酸酯板(透光率>95%)制作,顶部口径尺寸为100 cm,底部口径150 cm,高度48 cm。在本试验开展的2021年生长季,样地内的地表温度为17.21℃,土壤表层(0~10 cm)温度为10.09℃,土壤湿度为10.41%。增温样地内生长季地表温度较对照样地平均增温幅度为(1.31±0.3)℃,土壤表层温度平均升高了(0.55±0.2)℃[27]。每个小区面积为4 m2(2 m×2 m),小区间间距2 m。采用随机区组设计,增温处理和对照分别设10个重复,共计20个试验小区。
1.3 植被群落调查和分析
2021年8月,采用样方法进行植物群落调查,在每个试验小区,使用50 cm×50 cm的样方框,记录样方内的物种名称(精确到种),测定群落总盖度及各物种分盖度;然后用直尺测量3~5植株的自然高度,计算得到植物群落的重要值、丰富度、均匀度和多样性指数。
植物群落调查完成后,剪除地面全部植物,装入信封袋,65℃烘干至恒重,得到地上生物量。在去除地上植物的样方内,用内径3.5 cm的土钻分别在样方的四角钻取0~10 cm土芯,拣去石块、杂物,用流水冲掉附着物过筛后得到根系,65℃下烘干至恒重,获得地下生物量。
植物群落物种重要值IV、丰富度指数R、均匀度指数E、多样性指数H的计算公式如下:
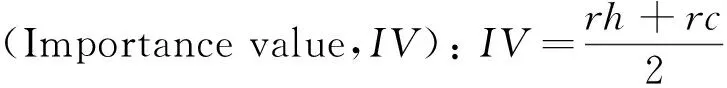
Patrick丰富度指数:R=S
式中,IV为重要值,rh为相对高度,rc为相对盖度;R为丰富度指数;H为多样性Shannon-Wiener指数;E为均匀度Pielou指数;i为样方框内的植物物种;Pi为第i个物种的重要值;S为样方框内的物种数。
1.4 叶片性状采样和测定
每个处理内分别选择7个植物物种进行叶片性状的测定,分别为禾本科(Poaceae)的垂穗披碱草(ElymusnutansGriseb.);莎草科(Cyperaceae)的四川嵩草(KobresiasetschwanensisHand.-Mazz.)和无脉薹草(CarexenervisC. A. Mey.);以及其他科属的蒲公英(TaraxacummongolicumHand.-Mazz.)、蓝玉簪龙胆(GentianaveitchiorumHemsl.)、小花草玉梅(AnemonerivularisBuch.-Ham. var. flore-minore Maxim.)和鹅绒委陵菜(PotentillaanserinaL.)。每个试验小区内分别选取7个物种的5~7株完成营养生长的植物,分别装入保鲜袋冷藏保存,尽快带回实验室。
将植物样品清洗干净后,将叶片从叶柄摘下,使用扫描仪(Epson STD 4800Q)进行叶片图像扫描,叶面积分析软件(WinFOLIA Pro 2020)进行叶面积分析,得到叶宽、叶长、叶周长、叶面积和长宽比,扫描完成后放入信封袋,65℃烘干至恒重,测定干重,计算比叶面积(Specific leaf area,SLA)。将烘干后的叶片样品研磨后过2 mm筛,采用凯氏定氮法测定叶片氮含量,钼锑抗比色法测定叶片磷含量,然后计算叶片氮磷比(N∶P)。
比叶面积(SLA)计算公式如下:
SLA(cm2·g-1)=La/Ld
式中,La为叶面积(cm2),Ld为叶片干重(g)。
1.5 统计分析
采用T检验分析增温处理与对照的植物群落特征和叶属性的差异性。双因素方差分析增温、物种及其交互作用对叶属性的影响。统计分析采用SPSS 26.0软件(SPSS Inc.,Chicago,Illinois,USA)进行,采用OinginLab 2021进行图形绘制。
2 结果与分析
2.1 增温对高寒草甸植物群落结构和生物量的影响
与对照相比,增温处理后丰富度指数显著降低了25.45%(P<0.05),而群落高度、盖度、多样性指数和均匀度指数均同对照无显著差异(表1)。增温处理后,植被地上生物量和地下生物量有增加趋势,但尚未达到统计显著水平。
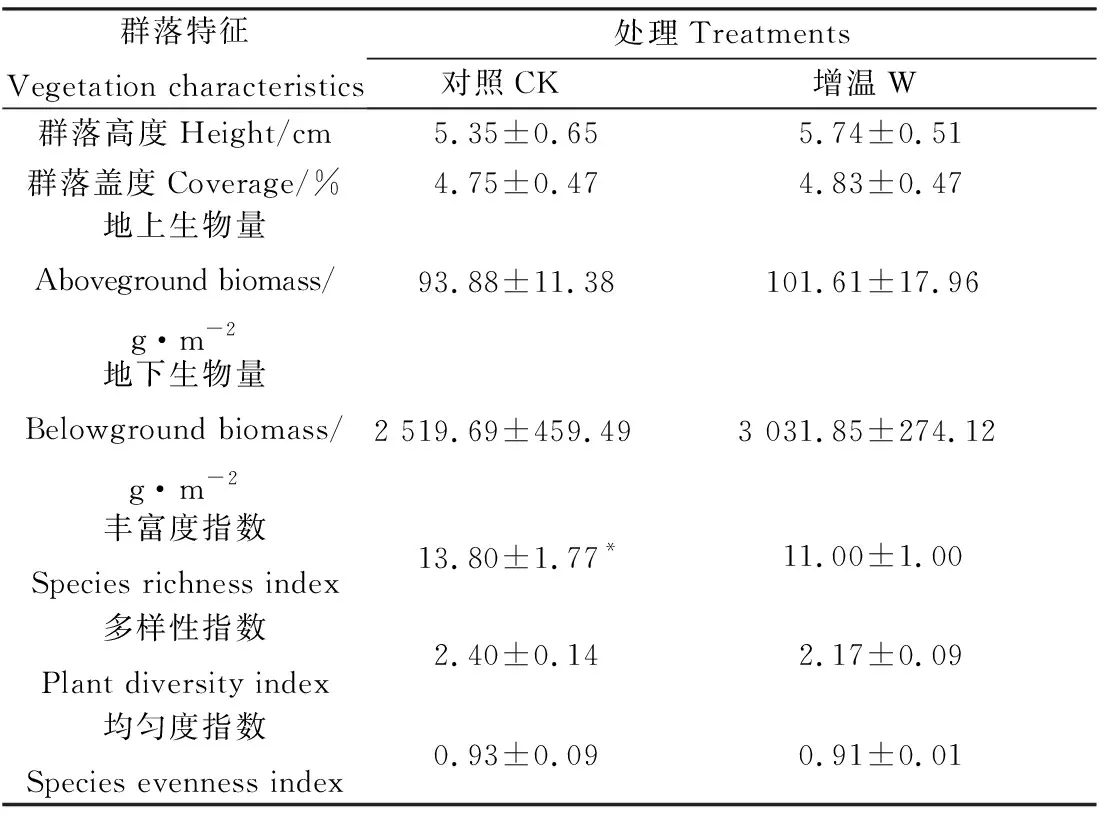
表1 增温对高寒草甸植物群落特征的影响Table 1 Warming effects on plant community characteristics in alpine meadow
2.2 增温对高寒草甸植物功能群的影响
增温处理后,禾本科植物的盖度和重要值均较对照组分别显著增加了329.56%和119.20%(P<0.05)。增温处理下,莎草科和杂类草两个功能群植物的高度、盖度和重要值与对照均无显著差异(表2)。双因素分析结果表明,功能群和增温对植物盖度和重要值均无显著影响,但二者的交互作用会显著影响植物盖度和重要值(P<0.05)(表3)。
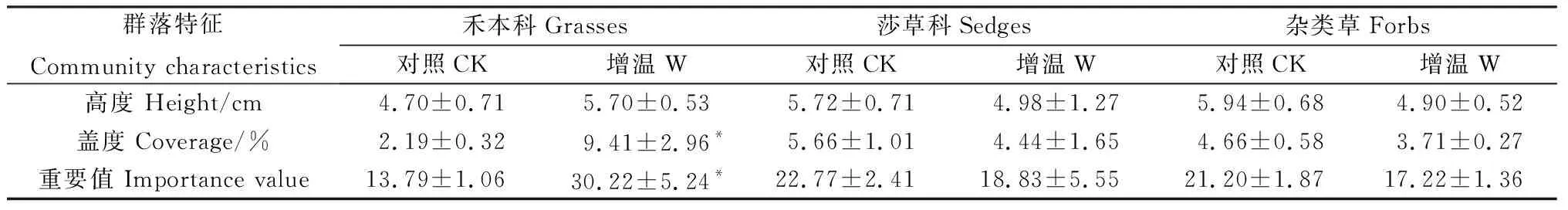
表2 增温对植物功能群特征的影响Table 2 Warming effects on plant functional groups characteristics
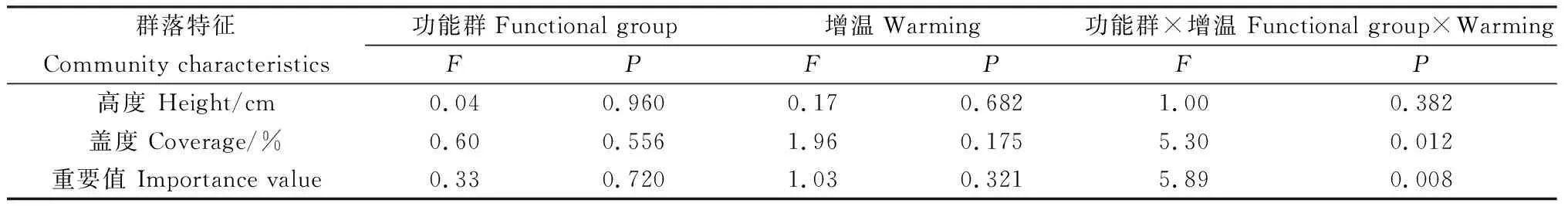
表3 功能群与增温对植物群落特征影响的双因素分析Table 3 Two-way analysis of variance (ANOVA) on the effects of functional groups and warming on the vegetation characteristics
2.3 增温处理对高寒草甸常见植物叶片性状的影响
如图1所示,与对照相比,小花草玉梅和鹅绒委陵菜的叶长分别显著增加25.20%和28.10%(P<0.05);蒲公英和鹅绒委陵菜的叶宽分别显著增加47.30%和44.82%;蒲公英、鹅绒委陵菜的长宽比分别显著增加26.70%和11.62%;鹅绒委陵菜的叶面积显著增加了48.40%(P<0.05)。增温后垂穗披碱草、蒲公英、小花草玉梅和鹅绒委陵菜的叶面积较对照分别显著增加了36.83%,45.14%,30.55%和47.92%;垂穗披碱草、四川嵩草、蓝玉簪龙胆、小花草玉梅和鹅绒委陵菜的比叶面积较对照分别显著增加38.83%,50.20%,11.30%,26.80%和32.14%(P<0.05)。与对照相比,增温处理后的小花草玉梅的长宽比显著降低了21.88%(P<0.05);垂穗披碱草、四川嵩草和无脉薹草的叶长、叶宽、长宽比和叶面积在增温后变化不显著;无脉薹草和蒲公英的比叶面积在增温处理后亦无显著变化。

图1 试验样地的设置和增温装置图Fig.1 Diagram of the experimental design and passive open-top chamber (OTC)
增温处理后,垂穗披碱草和无脉薹草的叶氮含量较对照显著增加了13.72%和17.21%;蒲公英和蓝玉簪龙胆叶磷含量较对照均显著增加了88.63%和94.82%;垂穗披碱草、无脉薹草和鹅绒委陵菜的N∶P较对照亦显著增加(P<0.05)(图2)。增温处理后,四川嵩草、蒲公英、蓝玉簪龙胆、小花草玉梅和鹅绒委陵菜叶氮含量同对照差异不显著;无脉薹草、小花草玉梅和鹅绒委陵菜的叶磷含量同对照无显著差异;四川嵩草和鹅绒委陵菜的N∶P同对照亦无显著差异。与对照相比,增温处理后的垂穗披碱草和四川嵩草的叶磷含量较对照显著降低59.40%和33.70%;增温处理后的蒲公英和蓝玉簪龙胆的N∶P较对照显著降低(P<0.05)(图2)。
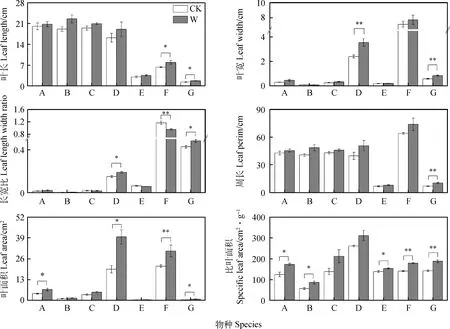
图2 不同植物叶片性状对增温的响应Fig.2 The responses of different plant leaf traits to experimental warmimg注:A,垂穗披碱草Elymus nutans;B,四川嵩草Kobresia setchwanensis;C,无脉薹草Carex evervis;D,蒲公英Taraxacum mongolicum;E,蓝玉簪龙胆Gentiana veitchiorum;F,小花草玉梅Anemone rivularis;G,鹅绒委陵菜Potentilla anserina。图中的误差线表示均值±标准误。下同Note:A, Elymus nutans;B,Kobresia setchwanensis;C, Carex evervis;D, Taraxacum mongolicum;E,Gentiana veitchiorum;F,Anemone rivularis;G, Potentilla anserina. Mean±SE is shown in error bars. The same as below
双因素分析发现,物种对叶宽、叶长、长宽比、周长、叶面积、比叶面积、叶氮含量、叶磷含量和N∶P的影响达到显著水平,增温对叶宽、叶长、周长、叶面积和比叶面积亦有显著影响(P<0.05),对长宽比和叶氮、叶磷和N∶P则无显著影响。物种与增温处理的交互作用对叶片长宽比、叶面积、叶磷和N∶P影响显著(P<0.05),对叶宽、叶长、周长、比叶面积和叶磷含量则无显著影响(表4)。
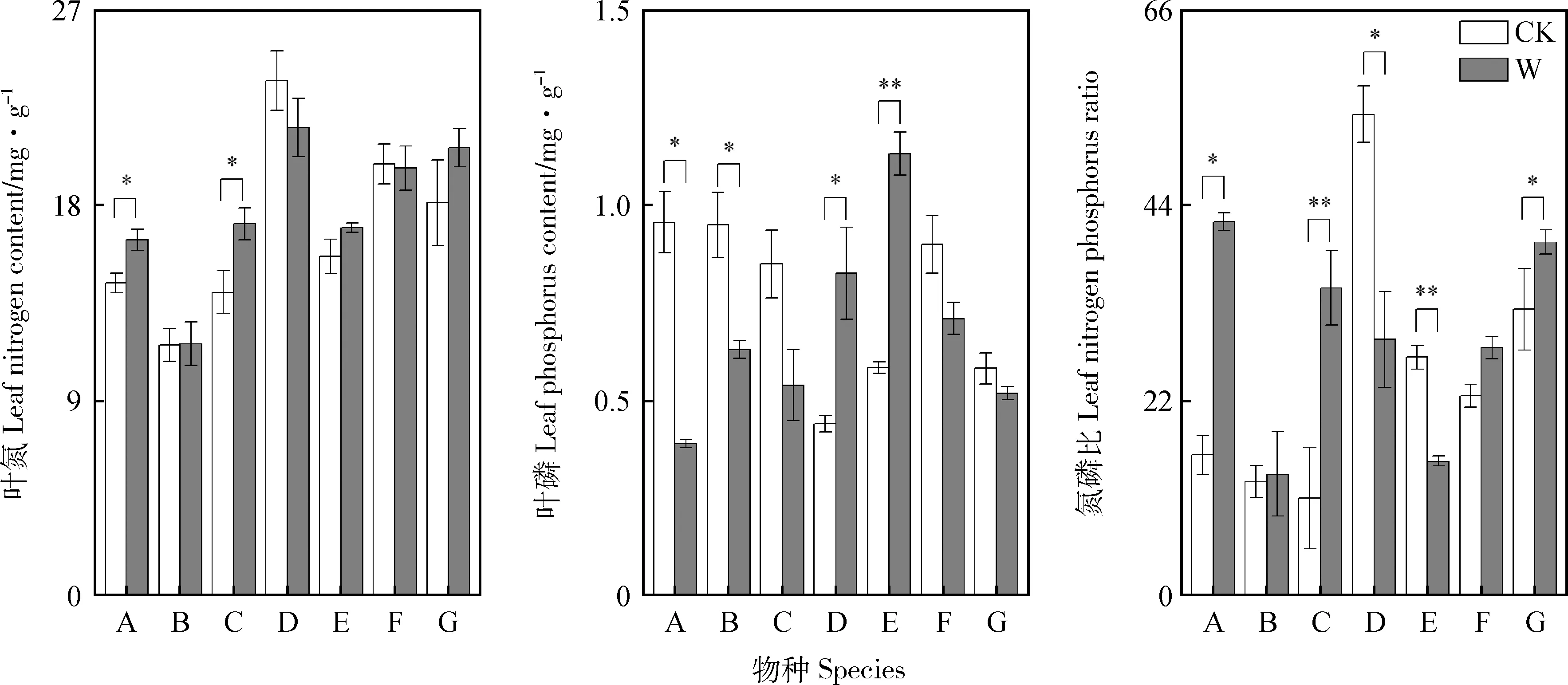
图3 增温对叶氮含量、磷含量和氮磷比的影响Fig.3 Effects of experimental warming on leaf nitrogen content,leaf phosphorus content and leaf nitrogen phosphorus ratio
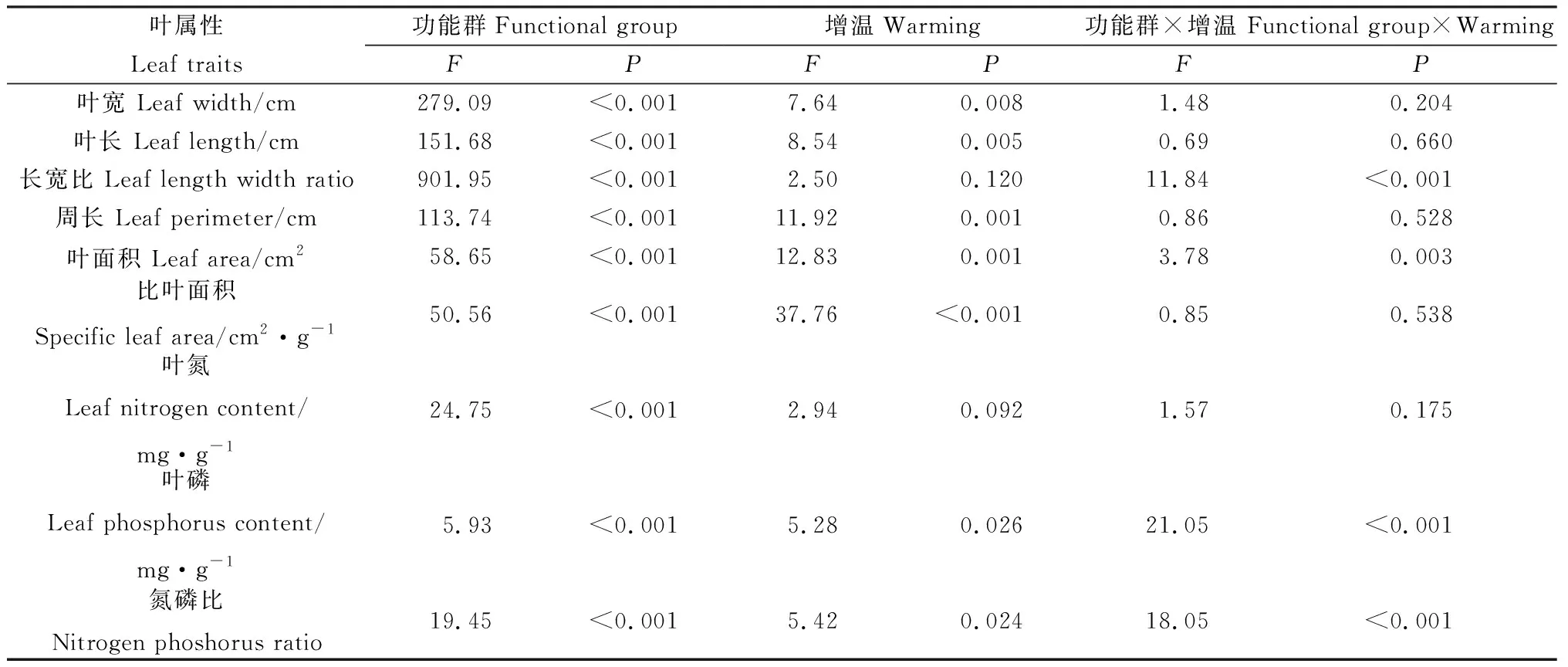
表4 物种与增温对植物叶片性状影响的双因素分析Table 4 Two-way variance analysis of the species and warming on plant leaf traits
3 讨论
3.1 增温对植物群落结构和生物量的影响
本研究发现,增温处理显著降低了植物群落丰富度指数,物种多样性指数和均匀度指数亦呈降低趋势,而植物群落高度、盖度、地上和地下生物量呈增加趋势。短期增温可以降低高寒草甸物种丰富度指数,长期增温更有利于优势物种的生长,但对群落的盖度、物种多样性指数和群落的组成的影响不明显[9,17,28]。本研究发现,增温后禾本科植物群落的盖度和重要值显著增加,而莎草科和杂类草植物的盖度和重要值呈下降趋势,表明增温对不同功能群物种的影响存在显著差异。在云顶山亚高山草甸开展的研究亦发现增温处理下禾本科植物的重要值呈增加趋势[29],说明短期增温有利于禾本科植物的生长[19],这与本研究的结果一致。增温普遍提高了禾本科植物的盖度,说明禾本科植物对增温更加敏感,在短时间气候变暖环境条件下可以更占优势[30-31]。
禾本科植物对暖干气候有较强的耐受性,增温处理下禾本科牧草具有更好的适应性[32]。增温后地表蒸散增高,禾本科植物的光合速率和分蘖能力增强,进而导致其盖度和优势度增加[33]。与禾本科植物相比,嵩草是高寒草甸中占有明显优势的莎草科植物,其根系主要分布在土壤表层,干热环境不利于其生长[17]。前人研究发现,增温对高寒草甸生物量的影响主要受水分条件的限制[34]。在湿润和半湿润区的草地生态系统中,增温可显著提高植物生物量;反之,在干旱的草地生态系统中,增温将加剧植物生长的水分限制,进而显著降低植物生物量[35-36]。本研究所处的川西北高寒草甸受高原寒温带半湿润季风气候调控,水分条件不是草地植物生长的限制因子,因而增温后植被地上、地下生物量均呈增加趋势。
3.2 增温对高寒草甸常见植物叶片性状的影响
在高寒草甸生态系统,土壤可利用性氮、磷含量是草地植被生长的限制因子。温度是调控植物生长和发育的重要环境因子,植物叶片对温度变化的响应极为敏感[37]。对比高寒草甸7种植物叶片对增温的响应,发现植物叶片性状对增温的响应存在明显种间差异[38]。增温后禾本科及莎草科植物叶片大小整体呈增加趋势,而杂类草的叶片大小无一致的响应趋势。本研究中,增温增加了垂穗披碱草、四川嵩草、无脉薹草、蓝玉簪龙胆、小花草玉梅和鹅绒委陵菜的比叶面积。前人研究亦表明,植物比叶面积会随着温度升高而逐渐增大[39],较高的比叶面积利于提高光合速率[40],进而适应增温环境。双因素分析发现,增温对植物叶宽、叶长、周长、比叶面积有显著影响;物种和增温处理的交互作用对长宽比、叶磷含量、N∶P影响显著,这说明增温和物种均会对植物叶片性状产生显著影响。
植物叶片N∶P的大小可以指示植物生长的氮磷限制因子[41]。增温后,垂穗披碱草、无脉薹草、蓝玉簪龙胆和小花草玉梅叶片N∶P显著升高,而蒲公英和蓝玉簪龙胆叶片N∶P显著降低,这表明增温可能会增加垂穗披碱草、无脉薹草和小花草玉梅生长的磷限制,而增加蒲公英和蓝玉簪龙胆生长的氮限制。本研究的植物普遍受到了单一资源磷的强烈限制,而增温加剧了这些物种的磷限制,导致了群落物种丰富度减少,符合“资源平衡假说”[42]。整体而言,同一生境的不同植物叶属性对增温处理表现出了不同的响应[24],证实了植物通过改变叶片性状来适应环境的规律[43]。
4 结论
增温显著降低了高寒草甸植物群落丰富度指数,显著增加了禾本科植物的盖度和重要值,但未显著改变其群落高度、盖度、多样性指数和均匀度指数。增温后,高寒草甸7种常见植物的叶宽、叶长和叶周长和比叶面积总体呈增加趋势,而叶片长宽比无显著变化。增温导致植物的叶磷含量呈降低趋势,而叶氮含量和氮磷比对增温的响应则因物种而异,表明高寒草甸植物对短期增温表现出了不同的适应策略。
