中短链脂肪酸甘油酯和乳酸杆菌对日本沼虾生长和免疫力的影响
2022-12-26熊贻伟傅洪拓
刘 帅, 蒋 丽, 杨 博, 熊贻伟, 傅洪拓*, 刘 波*
(1.上海海洋大学水产科学国家级实验教学示范中心,上海 201306;2.中国水产科学研究院,北京 100141;3.华南理工大学生物科学与工程学院,广东广州 510006;4.中国水产科学研究院淡水渔业研究中心,江苏无锡 214081)
水产业高质量发展对养殖环境提出了新要求,饲料端全面禁抗,养殖端减抗、替抗已成产业界及政府监管机构的共识。2019年,农业农村部发布《药物饲料添加剂退出计划》,对抗生素的退出提出了具体时间表。从抗生素过度使用到无抗生产,产业界急需要寻求新的促生长、提高抗逆性和品质、无药残的环境友好型替代物,中短链脂肪酸(林日添等,2017)、复合寡糖类物质(刘含亮等,2012)、大蒜素(宋文华等,2011)等几大类物质,都有望成为替抗的理想目标。
中短链脂肪酸是指由3~12个碳原子组成的脂肪酸类群,其分子质量小,穿透细胞膜能力强。养殖生产中常用的中链脂肪酸有己癸酸、月桂酸等,短链脂肪酸有甲酸、乙酸等。月桂酸单甘油酯(GML)是中链脂肪酸的一种。Li等(2009)发现,GML能够阻断猴免疫缺陷病毒SIV的传播,并且月桂酸单甘油酯在抑制HIV病毒、细胞巨化病毒、疱疹病毒、感冒病毒等方面的应用已经得到了广泛的验证(黄艺珠等,2020)。此外,GML还具有增加鱼类生长速度(蒋慧琪,2021)、增加蛋鸡产蛋量和改善肉质(Zhao等,2019)等功能,不仅在畜牧(Fortuos等,2019)养殖中发挥巨大作用,也对提高水产(Wang等,2021)动物的免疫起到显著效应。短链脂肪酸(SCFAs),即碳链6个以下的饱和脂肪酸,易被肠道吸收,可促进肠道发育,具有良好的供能、促免疫能力;丁酸作为应用最广泛的短链脂肪酸,在体内具有供能、抗菌功效(Harrison等,2013),常以添加剂的形式在水产(刘宇等,2020)、畜牧(薛永强等,2020)行业发挥动保功能。
乳酸杆菌能降解肠道中的碳水化合物生成乳酸,降低肠道内pH,抑制不耐酸的有害菌的繁殖,达到防病的功效;同时,其分泌的乳酸可以和钙离子结合,生成乳酸钙,促进甲壳类动物的脱壳与生长,因此在水产动物的生长免疫中(Geng等,2011)具有重要调节作用。
日本沼虾(Macrobrachium nipponense)俗名青虾、河虾,生活史13~14个月,雌雄生长差异很大,多代同堂是日本沼虾的繁殖特点,其生长速度雄虾在3 cm后逐渐与雌虾拉开距离,不同的饲养条件下其生长速度迥异,饲料系数参差不齐,且在进行集约化养殖时其产量往往被高频爆发的水产病害所限制,因此,在饲料中全面“禁抗”的大环境下如何通过优化日本沼虾的营养配方达到提高其整体抗逆性能成为当下行业的热点。已有研究证明,中短链脂肪酸联用时对病毒囊膜和细菌胞膜穿透力更强,具有更广谱的抗菌作用,且在酸性条件下,大部分病原微生物均会降低活性甚至失活,因而在酸性条件下中短链脂肪酸的组合抗病原微生物的能力更强(黄艺珠等,2020)。本研究通过添加乳酸杆菌菌液为中短链脂肪酸发挥功效构建酸性环境同时发挥其促免疫作用,探究中短链脂肪酸的联合使用对日本沼虾生长及免疫的效应,为生产实践应用提供理论支撑。
1 材料与方法
1.1 试验设计与饲料配比 试验所用的基础饲料组成见表1,其中,G0为空白对照组,G1为乳酸杆菌组,G2为0.15%月桂酸单甘油酯组,G3为0.15%丁酸甘油酯组,G4为0.15%月桂酸单甘油酯+0.15%丁酸甘油酯组,G5为0.2%月桂酸单甘油酯+0.2%丁酸甘油酯组。G1~G5组饲料以表1为基础,在投喂时将活性1.0×1010CFU/mL的乳酸杆菌菌液通过喷涂的方式以1:4(菌液/mL:饲料/g)的比例拌入基础饲料中形成最终的试验饲料(结果部分G1~G5组代表已经混入乳酸杆菌菌液的试验组),G0组以上述方式和比例拌入蒸馏水。
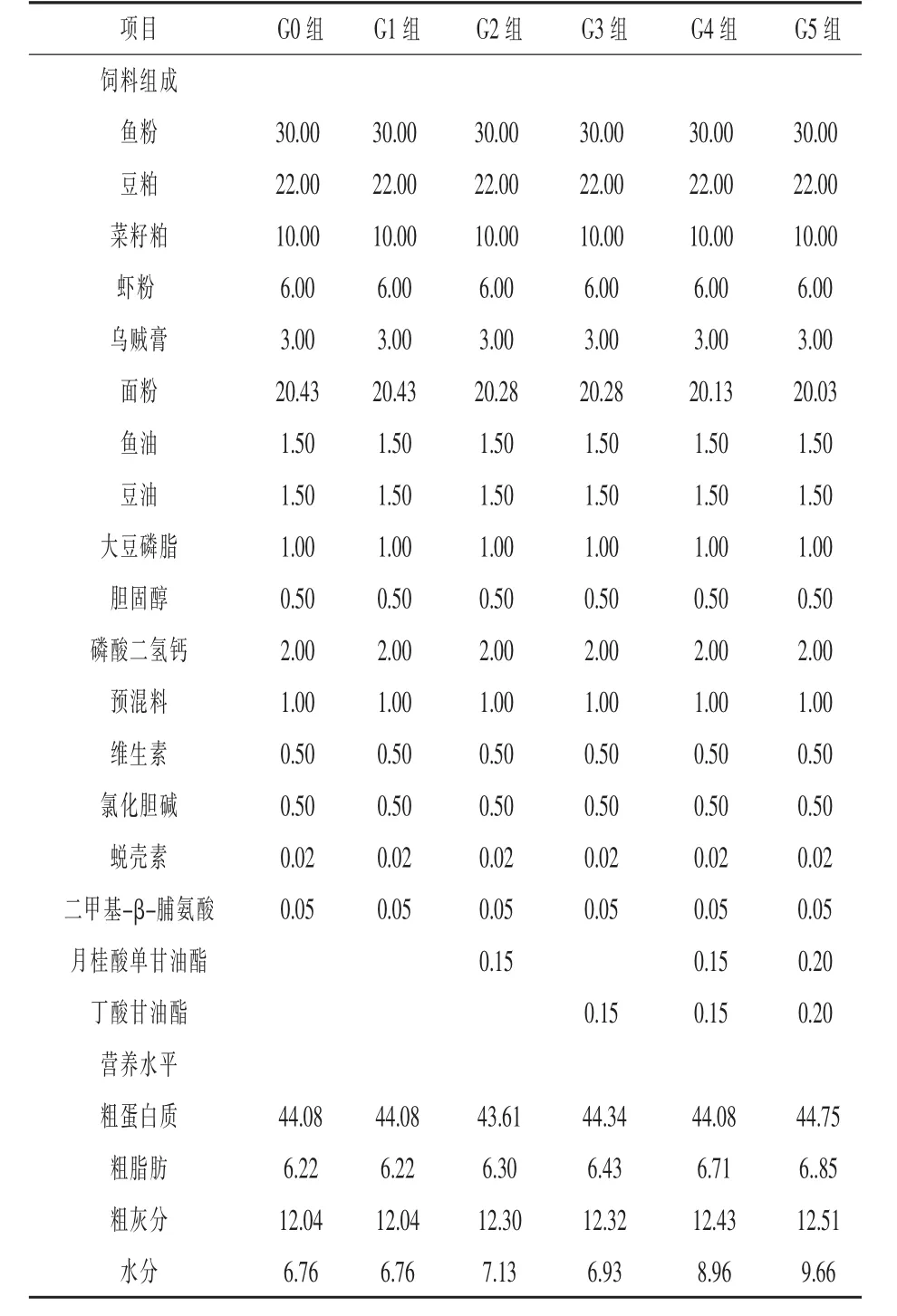
表1 基础饲料组成及营养水平%
所有原料均经粉碎过40目筛后,按表1的配方比例,加适量水后用搅拌机搅拌均匀,然后挤压成粒径为1.2 mm的长条,室温下风干,再用粉碎机破碎成小颗粒饲料,最后在-10℃冰柜中密封保存备用。月桂酸单甘油酯、丁酸甘油酯、乳酸杆菌等由广州佰仕路生物科技有限公司生产,其中,饲料制备过程中添加的月桂酸单甘油酯、丁酸甘油酯干粉的纯度分别为55%、50%。
1.2 试验动物与养殖方法 试验用虾来源于中国水产科学研究院淡水渔业研究中心宜兴大浦基地;虾苗放入300 L暂养缸中进行2周驯化以备待用,驯化期间使用基础日粮饲料;驯化结束后停止投喂1 d,将初始体重(0.401±0.007)g的900尾虾苗(不区分雌雄)按照每个养殖桶50尾,随机分入18个桶(直径1.8 m高1 m),每个处理三个养殖桶。试验期间,保持桶内水深30~40 cm,pH为6.5~7.5,使用气泵对水体不间断供氧使溶氧量维持在5 mg/L以上;每次投喂1 h后清理残饵与粪便,并通过换水(每3 d一次,换水量20%~30%)使氨氮低于0.2 mg/L、亚硝酸盐含量保持在0.1 mg/kg以下;水温18~25℃;每日的7:00~8:00、11:00~12:00、16:00~17:00各投喂1次,投饵率大约为体质量的2%~5%,并根据每周平均摄食状态进行调整,进行为期8周(9月中旬~11月中旬)的养殖试验。
试验第1周出现死虾要及时补充相同数量和体重相当的虾苗;自第2周起,各试验桶的死虾要捞出称重并记录,直至试验结束不再补充虾的数量。
1.3 样品收集 试验结束后,对虾停止投喂24 h后再进行采样以减少采样过程中虾的应激反应。详细采样策略如下:首先收集每个桶中的虾,分别记录数量与体重;其次,每组取18尾虾(每组3个桶,每桶抽取6尾虾),用含有0.5 mL抗凝剂的一次性注射器从虾的第3步足基部抽取大约0.5 mL的淋巴血,样品在4℃冰箱1~2 h后,4℃、3000 r/min离心10 min,收集起上清转入抗凝管中于-80℃保存,每桶3管血样,即每组9管血样,以备相关抗氧化、免疫酶活性的测定;采血后,收集每只虾的肝胰腺于冻存管转入液氮保存运输后转入-80℃冰箱长期保存,保证每桶6个样品,以备相关抗氧化酶活性以及基因表达的测定。考虑样本量以及个体差异等因素(Li等,2014),单个血样或肝胰腺样品是由同一试验桶2或3尾虾的组织混合而成。
1.4 生长性能分析
增重率/%=(W2-W1)/W1×100;
存活率/%=N1/N0×100;
特定生长率/%=(ln W2-ln W1)/T×100;
饵料系数=W3/(W2-W1);
式中:W1、W2和W3分别为养殖前后虾的平均体重及每尾虾吃食量均重,N0和N1分别为养殖前后试验桶内日本沼虾的数量,T为养殖天数。
1.5 抗氧化指标分析 使用南京建成生物工程研究所有限公司的相关试剂盒对虾的血清及肝胰腺组织总超氧化物歧化酶(T-SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-PX)、酸性磷酸酶(ACP)、碱性磷酸酶(AKP)酶活性和一氧化氮(NO)、丙二醛(MDA)活性物质含量进行测定评价,以及对抗超氧阴离子(ASAFR)能力进行测试;血清羟自由基清除能力(HRSA)通过苏州格瑞思生物科技有限公司生产的试剂盒进行测定,其中肝胰腺组织样品为经过冰生理盐水清洗,滤纸吸干,称重,捣碎后移入匀浆器中冰水浴匀浆,并4℃、2500 r/min离心10 min后的上清液。
1.6 荧光定量 使用Trizol法提取虾中肝胰腺的总RNA,使用NanoDrop 2000 spectrophotometer仪器测定OD值并使样本定容到500 ng/mL,采用南京诺威赞生物科技股份有限公司的HiscriptIII RT SuperMix for qRT-PCR试剂盒按20μL体系反转录为cDNA,每个组别8个重复,设置βactin为管家基因,采用ChamQTMUniversal SYBRqPCR Master Mix试剂盒对日本沼虾Dorsal蛋白(Dorsal)、热休克蛋白60(Hsp60)、热休克蛋白70(Hsp70)、Relish蛋 白 (Relish)、Toll样 受 体(Toll)和髓样分化因子Myd88(Myd88)进行荧光定量,定量结果采用2-ΔΔC法(赵姣姣等,2019)进行计算分析;反转录所用引物序列采用前人的研究 (Song等,2020;Luo等,2018;Li等,2018;Gu等,2017)成果并由上海捷瑞生物工程有限公司进行化学合成,引物序列见表2。

表2 目标基因与引物序列
1.7 数据分析 试验数据采用“平均值±标准差”表示,分析软件为SPSS 20.0,试验数据先进行方差齐性检验;方差齐性后再以组别为变量进行单因素方差分析(One-way ANOVA)及Duncan’s多重比较。P<0.05表示差异显著。
2 结果
2.1 中短链甘油酯对日本沼虾生长的影响 由表3可知,在末尾均重方面G1和G4组与空白对照G0组相比,均差异显著(P<0.05);增重率方面,G1组和G4组分别较空白对照G0组提升24.99%和26.69%(P<0.05);而在存活率中各试验组与空白对照组之间均不存在显著性差异(P>0.05);在特定生长率中,试验G1、G4、G5组分别较G0组提升18.92%、18.92%和14.41%(P<0.05);在饲料系数中,G5组显著低于空白对照组28.96%(P<0.05)。
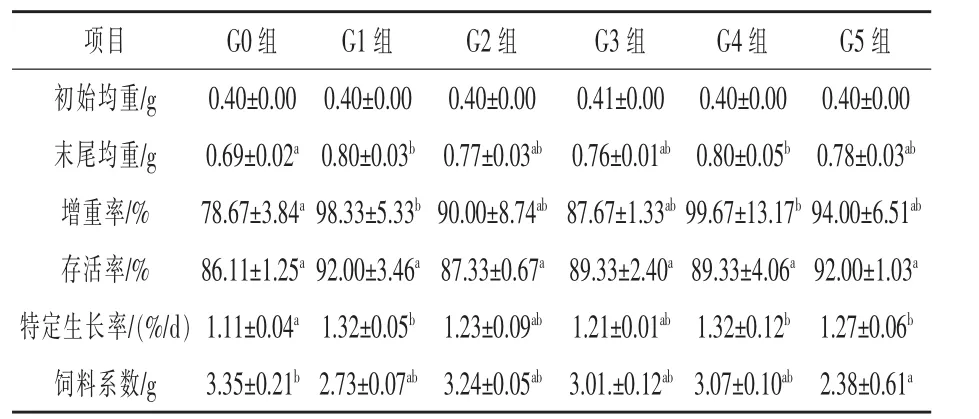
表3 不同添加剂处理下日本沼虾的生长性能表现
2.2 中短链甘油酯和乳酸对日本沼虾非特异性免疫性能的影响
2.2.1 血清抗氧化结果 由表4可知,G1组和G3组分别MDA的含量较G0组提升60.71%和47.61%(P<0.05),G2、G4和G5组与G0组不存在显著性差异(P>0.05);G2、G4、G5组CAT含量分别较G0组提升219.22%、140.94%、431.68%(P<0.05);T-SOD测定结果显示G2、G3、G5组分别较G0组提升50.92%、62.77%、115.36%(P<0.05)。

表4 不同饲料添加剂处理下虾血清中与免疫相关的酶活性变化
2.2.2 肝胰腺组织抗氧化酶活 由表5可知,在GSH-PX酶活性方面,G1、G3、G5组分别较对照G0组提升50.37%、56.09%、50.66%(P<0.05);ACP酶活性方面,G2和G4组分别较G0组提升28.76%和33.31%(P<0.05);NO含量上,G1、G2、G5组分别较G0组提升41.13%、42.74%、43.55%(P<0.05);ASAFR上,G1、G2、G3、G4和G5组较G0组分别提升176.61%、73.90%、230.66%、196.07%和136.81(P<0.05);AKP活性方面,各组之间不存在显著性差异(P>0.05);HRSA活性方面,试验G1~G5组分别较G0组提升34.17%、32.65%、31.44%、25.44%和23.32%(P<0.05);MDA含量方面,试验G1~G5组分别较G0组降低39.28%、53.16%、53.98%、37.84%和41.04%(P<0.05)。

表5 不同饲料添加剂处理下虾肝胰腺抗氧化酶活的变化
2.2.3 肝胰腺免疫抗氧化荧光定量 荧光定量结果如图1,在Myd88基因的表达上,G1与G5组分别较G0组提升14.69%和35.17%(P<0.05);IMD基因的表达上,仅G2较G0组提升47.49%(P<0.05)其余各组与G0无显著性差异(P>0.05);HSP70基因 的 表 达 上,G1、G2、G3、G4和G5分别较G0组提升315.10%、293.03%、331.75%、297.35%和379.43%(P<0.05);在HSP60基因的表达上,G1和G4组分别较对照组G0降低24.46%和40.45%(P<0.05),而G3组和G5组则分别较G0组提升44.30%和69.74%(P<0.05)。在Relish、Dorsal和Toll基因的相对表达量上,各组之间不存在显著性差异(P>0.05)。

图1 不同饲料添加剂处理对免疫抗氧化相关基因相对表达量的影响
3 讨论
3.1 中短链甘油酯和乳酸杆菌对日本青虾生长的影响 中短链脂肪酸甘油酯和乳酸通过改善肠道微生物菌群促进生长,在畜禽上应用已经非常广泛(徐晓燕等,2012)。本研究发现,丁酸甘油酯、月桂酸单甘酯与乳酸杆菌联用的G4组可提高日本沼虾的生长性能。无论是体重、增重率还是特殊生长率,三者联用都与对照差异显著,与畜禽上促进生长的影响一致。由此可看出,中短链脂肪酸在促进日本沼虾的生长方面具有良好的效果,在不改变其他环境条件下,能显著提升其生长速率,在相同的饲喂喂条件下,能显著提高饲料利用效率。中短链脂肪酸甘油酯在传统的营养需求中,与长链脂肪酸不同,是不包含在营养需求中的组分,本研究利用联合补充中短链脂肪酸甘油酯,在改善生长性能方面,获得了良好的效果,说明中短链脂肪酸甘油酯在日本沼虾的营养需求中,是不可或缺的营养组成,其用量不大,却能起到重要的生长促进作用。
中短链脂肪酸以及其衍生物和乳酸杆菌发挥作用的部位是生物体的肠道,即通过促进脂类代谢和调整肠道稳态的方式(Hu等,2006)加快机体的代谢速率,从而实现机体的快速生长与发育。本研究的后期,水温降低到20℃以下,单独加入乳酸杆菌菌液的G1组和三种添加剂联用的G4组仍然在增重率和特定生长率上表现出与G0组的显著差异,这说明即使在非最适温度下养殖日本沼虾,乳酸杆菌依然可以对日本沼虾的生长产生显著促进作用,在一个生长周期内,其促生长的效应显著;乳酸杆菌对虾生长免疫方面的促进作用(Wang等,2019)早已被证实,与常规的益生菌发酵饲料(Abu-Elala等,2020)不同,本试验的乳酸杆菌是在投喂前才与饲料混合,其新鲜度更高,生物活性更好,因而发挥的功效更强。此外,扩散到水体中的乳酸杆菌液呈酸性,因而能够对水体的pH进行调节,对水环境的稳态有一定的维持作用,这也是促进日本沼虾快速增重的一个因素,只添加乳酸杆菌的G1组相较于其他各组增重率最高,达到了98.33%。在增重率上,三者联用添加比例较低的G4组不仅高于添加比例较高的G5组,还高于乳酸杆菌联用单一中短链脂肪酸的G2、G3组,这说明单一中短链脂酸可能会对乳酸杆菌发挥功效起一定制约作用,当两者联合使用且达到一定阈值时,才能发挥其正向的促进作用。
本研究中饲料系数普遍偏高,这可能是由于在试验的后期水温没有达到日本沼虾的最适生长温度,导致试验虾摄入体内的饲料大多消耗于维持体温等日常代谢的稳定,从而降低了饲料的转化效率,致使饲料系数在计算上偏大。
3.2 中短链甘油酯和乳酸杆菌对日本青虾非特异性免疫的影响 甲壳类动物普遍缺乏获得性免疫能力,往往只能依赖先天非特异性免疫(袁伟等,2020);日本沼虾作为一种典型的甲壳类动物,其主要在血清、肝脏中通过相关酶的分泌获得非特异性免疫(管晓娟,2009),因而可以用特定酶活性衡量其免疫能力。现行的养殖体系中,为了提高养虾抗病毒能力,会在养殖过程中,少量使用各类抗生素类物质,但是在实际生产中,日本沼虾感染白斑综合征(WSSV)时有发生,在目前全球限抗和替抗的行业需求背景下,寻求营养物质类替抗新品成为产业急需要解决的问题。已经有研究表明,多糖对南美白对虾的免疫有显著的功能(季露等,2021)。本研究通过中短链脂肪酸的合理配伍,科学地评估了中短链脂肪酸对日本沼虾免疫性能的影响。
MDA、T-SOD、ACP等酶的活性或含量是评估生命体非特异性免疫的重要辅助指标。MDA是机体内自由基与脂质过氧化反应产生的具有细胞毒性的化学物,可以间接反映机体组织的氧化受损程度,被作为评估机体抗氧化能力的一个重要参数;T-SOD作为动物体内经典的抗氧化酶之一,具有维护机体氧化与抗氧化平衡的作用,是动物体内抗氧化的代表酶之一,在免疫指标评估中占据重要的地位;ACP是非底物特异性酶,参与机体钙磷代谢,与碱性磷酸酶构成经典的非特异性酶。在本研究中,月桂酸单甘油酯、丁酸甘油酯及乳酸杆菌三者联用的试验组G4和G5在MDA、T-SOD及ACP上都表现出与空白对照G0组的显著差异,且MDA与T-SOD的活性提升是单向同步的,证实了三种添加剂具有提升日本沼虾免疫能力;此外,单一添加月桂酸单甘油酯的G2组与单一添加丁酸甘油酯的G3组的T-SOD活性分别为9.04 U/mL和9.75 U/mL,显著低于两者联用且添加比例相对较高的G5组的12.90 U/mL,但却显著高于同样是两者联用但添加比例相对较低的G4组的7.63 U/ml,说明月桂酸单甘油酯及丁酸甘油酯两者之间可能存在一定的交互作用,且仅当两者的含量达到一定阈值时才会对免疫力产生显著的影响,这也是许多研究者呼吁在动物饲料中联用中短链脂肪酸的一个依据(刘宇等,2020)。
当机体长期处于应激状态时,机体的免疫能力会有所下降(Tahmasebi-Kohyani等,2012),因而可通过测定机体相关的应激蛋白表达量侧面评价机体的免疫能力(史晏如,2016);HSP60、HSP70分别为热休克蛋白的一种,当机体发生应激反应(如高温、低温和饥饿等)时,其表达量会相对增加从而减少由应激导致的机体损伤(Lv等,2021),因而通常将其相对表达量作为衡量生命体非特异性免疫的重要指标之一;在上述结果中,只加入乳酸杆菌的G1组对比G0组,HSP60相对表达量有所下降,此外,在加入乳酸杆菌的基础上,分别加入月桂酸单甘油酯或丁酸甘油酯都可以促进HSP60的表达,但两者加入剂量不同时,对机体的表达量的影响也不同,两者联用且相对添加比例为1.5%时(G4组)会抑制其表达,而两者联用且相对添加比例达到2.0%时(G5组)会促进其表达,这说明两种中短链脂肪酸之间在低浓度时可能存在一定的拮抗作用,而在高浓度时存在一定的互作效应;HSP70基因荧光定量结果显示,各试验组相对表达量显著高于空白对照组,说明乳酸杆菌具有促进HSP70表达的效应。
免疫缺陷同系物(IMD)信号通路(Li等,2013)及Toll信号通路(Xi等,2008)在虾类先天免疫系统中扮演着重要的角色;IMD信号通路的模式识别受体结合病原体后将促进抗菌肽的表达从而实现对革兰氏阴性菌入侵的防御;Toll信号通路具有抵抗细菌和病毒感染的能力(De Gregorio等,2002)。Myd88是Toll信号通路中的主要衔接蛋白之一,与细胞内活性氧的产生息息相关(Sharp等,1992)。本研究中Myd88的表达量在G5组达到最高,当前鲜见中短链脂肪酸影响IMD、Relish、Toll、Dorsal和Myd88表达的相关研究,但已有研究证实了IMD和Toll信号通路在疾病防御方面具有重要作用(Liu等,2016),与信号通路相关的免疫效应基因转录的增加表明了对病原体入侵的免疫反应,说明月桂酸甘油酯和丁酸甘油酯两者的含量达到一定阈值时能促进免疫力的提升,这也与血清T-SOD酶活性变化的趋势相一致。
4 结论
本研究结果表明,乳酸杆菌对日本沼虾的生长具有显著促进作用,在加入乳酸杆菌的基础上联合使用中短链脂肪酸不仅能促进生长,还能促进免疫力提升;从试验设计来看,丁酸甘油酯0.15%、月桂酸单甘酯0.15%和乳酸杆菌25%联用可显著提升日本沼虾血清中CAT酶活、肝胰腺组织ACP酶活及羟自由基清除能力,并降低肝胰腺组织中MDA的含量,同时还可促进热休克蛋白HSP70的表达,达到显著提升日本沼虾免疫能力的效果;从应用角度看,中短链脂肪酸可以作为营养成分,起减抗甚至是替抗的作用,在高质量健康养殖中意义重大,其微量但效果显著,具有良好的应用前景。
