不同连作年限菊花根际土壤微生物多样性变化特征
2022-12-24张晓波于春雷张文洋
张晓波,于春雷,张文洋,闫 烨,阮 芳
(辽宁省经济作物研究所,辽宁 辽阳111000)
菊花(Chrysanthemummorifolium)为菊科菊属多年生草本花卉[1],是我国十大传统名花,也是世界四大切花之一。我国是亚洲切花菊市场的主要供应地,菊花主要产区多采用每年2.5~3.0茬的高效生产模式。连续种植3 a以上就会导致菊花连作障碍问题,病虫害加重,植株生长发育不良,降低了切花的观赏与经济价值[2-3]。
根际土壤微生物数量庞大且功能活跃,在植物生长方面的作用显著[4],是植物生长必不可少的基础[5]。微生物多样性、丰度和群落结构是土壤生态系统功能和平衡的晴雨表[6],能够反映土壤的健康情况[7]。大量研究表明,长期连作一方面改变土壤基本理化性质,造成土壤养分不平衡,导致土壤微生物群落结构改变,进而影响植物生长发育[8];另一方面影响根际土壤微生物活性,导致有害微生物增加,种群结构失衡[9],最典型的表现是病原菌增多,有益微生物减少,土传植物病害频繁发生[10]。显然,根际微生态的变化是连作障碍的主要原因[11-12],但不同连作年限菊花根际微生物多样性变化尚不明晰。
土壤微生物常规的研究方法,比如平板培养法、生物鉴定系统法和标记法,不能详细统计出土壤微生物的群落结构、生物多样性信息,以及不同群体生理差异。第二代测序Illumina MiSeq凭借测序快、通量高和可信度高等优势,在根际土壤微生物区系研究中得到广泛应用[13-17]。本研究采用高通量测序技术,以不同连作年限菊花根际土壤为试验材料,揭示连作根际土壤微生物群落结构和多样性变化特征,明确菊花连作障碍的形成机制,为菊花产业可持续发展提供科学依据。
1 材料与方法
1.1 土壤样品采集
土壤样品采自长期种植菊花的日光温室,位于辽宁省辽阳市白塔区辽宁省经济作物研究所花卉研发区。菊花连作年限分别为无连作(CK)、连作2 a(2a)、连作9 a(9a),每个年限设置3个重复小区,统一进行田间管理。2021年11月盛花期,按多点取样法于每个小区内随机选取20株长势均匀的菊花,采用抖根法收集根际土壤,混合均匀并分成2份,一份存放于-80 ℃超低温冰箱用于土壤微生物分析,另一份4 ℃保存备测。
1.2 土壤DNA提取与MiSeq测序
样本土壤总DNA的提取采用试剂盒(HiPure Soil DNA Mini Kit,广州美基生物科技有限公司)。利用1%琼脂糖凝胶电泳检测抽提的基因组DNA,Qubit2.0Flurometer检测DNA样品浓度。采用ABI GeneAmp®9700型PCR仪,用通用引物338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)扩增细菌16S rDNA基因V3-V4区;以ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2(5′-GCTGCGTTCTTCATCGATGC-3′)作为引物,扩增真菌ITS rRNA基因序列ITS1高变区。PCR反应采用25 μL体系,其中含5×FastPfubuffer 5 μL,dNTPs(2.5 mmol·L-1)2.5 μL,上、下游引物(5 μmol·L-1)各1 μL,FastPfupoglymerase 0.5 μL,模板DNA 10 ng,补ddH2O至25 μL。PCR扩增条件:95 ℃ 3 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 45 s,27次循环;72 ℃ 5 min,10 ℃保存。同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测,使用AxyPrepDNA凝胶回收试剂盒(Axygen公司)切胶回收PCR产物,Tris-HCl洗脱定量后,用QuantiFluorTM-ST蓝色荧光定量系统(Promega公司)进行定量检测,按照每个样本的测序量要求,进行相应比例的混合。MiSeq测序分析由上海元莘生物医药科技有限公司进行。
1.3 数据处理
Novaseq 6000高通量测序所得到的原始数据,利用Cutadapt(version 1.16)软件除去其双端序列接头和低质量碱基,再使用FLASH(v1.2.11)软件进行拼接。拼接后的原始序列通过Usearch(v7.0.1090)去除嵌合体序列从而得到优质序列。
使用UPARSE(v7.0.1090)将OTU(operational taxonomic unit)按照97.00%的相似性阈值聚类。并选取每个OTU中丰度最高的序列作为该OTU的代表序列。RDP Classifier针对Silva(SSU132)16S rRNA数据库,使用70%的置信度阈值分析每个OTU代表序列的注释结果。为了保证分析结果的准确性,精简去掉丰度值小于总序列条数0.001%的OTU,后续分析均使用精简后的OTU列表。
基于Mothur v.1.21.1软件进行稀释曲线分析以揭示多样性指数,包括Chao、ACE和Shannon多样性指数。使用Bray-Curtis和PCoA分析比较群落差异,进行β-多样性分析。利用SPSS 22.0软件进行单因素方差分析(Duncan法),显著性水平选定为α=0.05。
2 结果与分析
2.1 微生物群落OTU特异性
从菊花根际土壤微生物维恩图(图1)可知,CK、2a、9a 3个处理的土壤细菌OTU数量分别为5 408、6 534、6 041个(图1-A)。其中,共有OTU 3 388个,占相应样品OTU总数量的63%、52%和56%;独有OTU数量分别为1 044、1 210和859个,占相应样品OTU总数量的19%、19%和14%。CK、2a、9a 3个处理土壤真菌OTU分别有1 172、1 466、1 168个(图1-B)。其中共有OTU 481个,占相应样品OTU总数量的41%、33%和41%;独有OTU数量分别为382、557和304个,占相应样品OTU总数量的33%、38%和26%。上述结果可以看出,独有OTU数量在连作2 a时最多,连作9 a时最少,2a、9a的共有OTU数量最多。表明短期连作可能引起某些土壤微生物的富集,但随着连作年限延长,一些微生物种类丧失,导致根际土壤微生物群落多样性发生改变。

图1 不同连作年限菊花根际土壤细菌(A)和真菌(B)群落维恩图
2.2 细菌、真菌丰度和多样性
如表1所示,真菌、细菌覆盖度指数均在0.99以上,说明样本序列检出概率高,测序结果能够说明不同种植年限菊花土壤微生物群落真实情况。随着种植年限的增加菊花根际土壤细菌和真菌的ACE指数和Chao指数均先升高后降低。2a和9a的细菌丰度指数显著高于CK,两者之间差异不显著;不同年限真菌丰度指数没有显著性差异。群落多样性指数方面,土壤细菌和真菌的Shannon指数与丰度指数变化趋势相似,Simpson指数先降低后升高。其中,2a的细菌Shannon指数显著高于CK和9a。
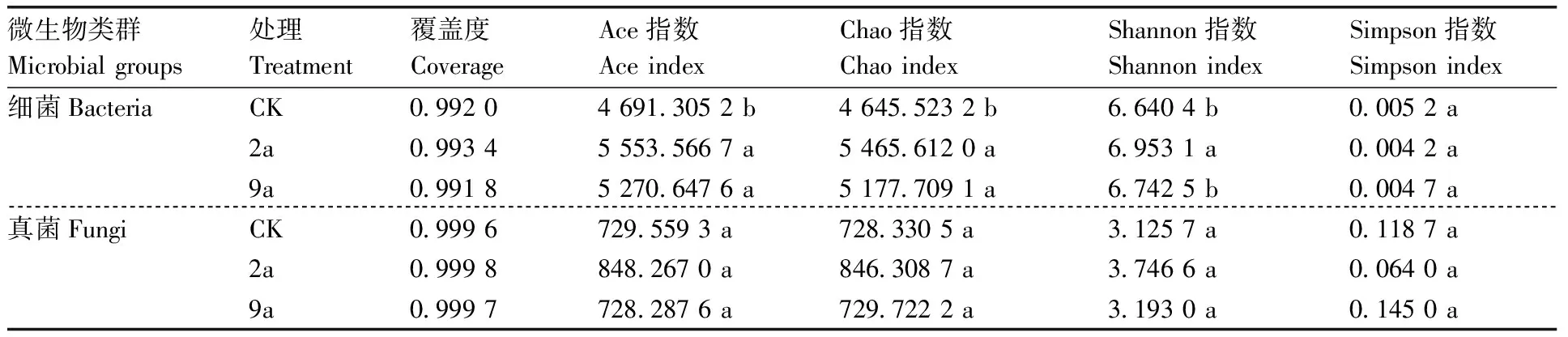
表1 不同连作年限菊花根际土壤细菌和真菌群落丰富度与多样性指数
2.3 细菌和真菌群落组成相似性
应用R语言vegan包、vegdist和hclust进行距离计算和聚类分析,将3个年限根际土壤微生物群落属水平上丰度排名前50的物种进行丰度相似性聚类,绘制heatmap图。利用颜色深浅表示属水平上群落组成的相似性和差异性,红色表示相似性最高,蓝色表示相似性最低。如图2-A所示,3个年限根际土壤细菌群落相对丰度均有明显差异,其中厌氧绳菌属(Anaerolineaceae_uncultured)、RB41、酸杆菌属(Acidobacteria_norank)、芽孢杆菌(Bacillus)、黄杆菌属(Flavobacterium)、Rhizomicrobium等细菌属差异性较大。样本间聚类结果显示,2a与9a聚为一类;图2-B显示了真菌群落的相似性,其中,被孢霉属(Mortierella)、未分类真菌属、Olpidiaster_unclassified、明梭孢属(Monographella)等相似性较大;异茎点霉属(Paraphoma)、Gibellulopsis、毛壳菌属(Chaetomiaceae_unclassified)、火丝菌属(Pyronemataceae_unclassified)、土赤壳属(Ilyonectria)等在不同年限根际土壤中差异性较大。样本间聚类结果为CK与9a聚为一类。
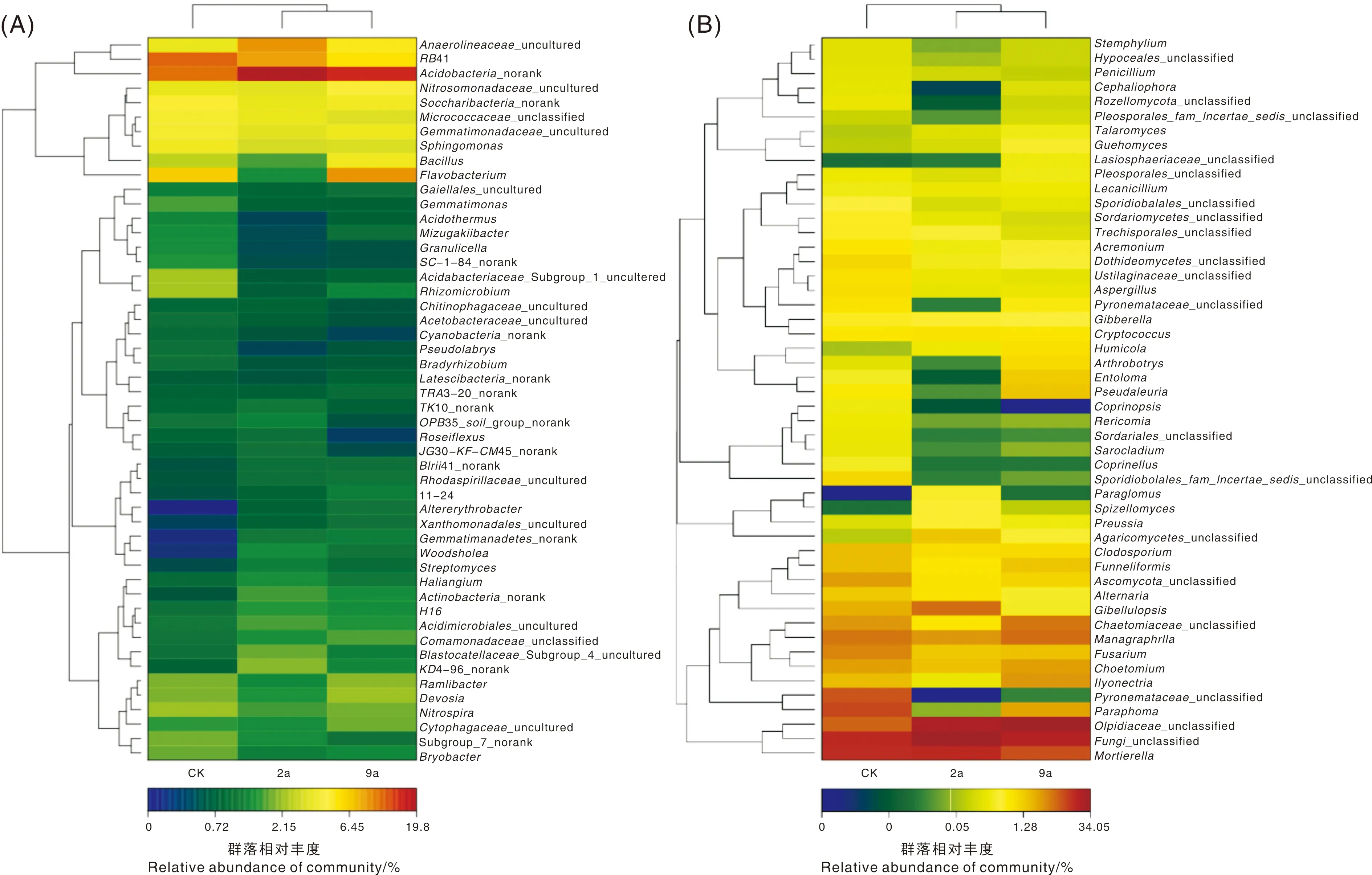
图2 不同连作年限菊花根际土壤细菌(A)和真菌(B)属水平群落热图
为了进一步说明群落组成的相似性,对不同连作年限土壤样本的OTU组成进行PCoA分析,分别提取两个主成分因子,用来表征细菌和真菌群落的特征。从细菌PCoA二维图可知(图3-A),PC1是造成样品差异性最大的主要因素,解释度为50.54%;PC2次之,解释度为17.75%,两个主坐标共解释了68.29%的样本信息。在PC1上,2a和9a距离较近,二者与CK距离较远,说明连作后的菊花根际土壤中细菌群落结构相似性较高。在真菌PCoA二维图中(图3-B),两个主坐标PC1与PC3共解释了50.44%的样本信息。9a与CK距离更近,并与2a距离较远。在真菌群落结构方面,CK与9a相似性更高,这与聚类热图对不同年限群落组成相似性分析结果一致。

图3 不同连作年限菊花根际土壤细菌(A)和真菌(B)PCoA分析
2.4 细菌和真菌群落结构分析
不同分类水平能够具体反映土壤微生物群落结构状态,本研究分别在门分类水平和属分类水平上对细菌、真菌菌群组成进行分析。
在门分类水平上,土壤细菌分属于45个门,共有主要菌门11个(图4-A),在3个年限细菌群落总数的占比分别为96.01%、96.25%、97.06%。土壤真菌共有主要菌门5个(图4-B),在3个年限真菌群落总数中占比分别为66.60%、84.01%、80.01%。从整体分布看,不同年限土壤细菌群落相对丰度变化较小,真菌群落相对丰度差异显著。2a土壤真菌优势门以子囊菌门为主,CK、9a土壤真菌优势门以壶菌门和子囊菌门为主;2a、9a的子囊菌门相对丰度显著高于CK。
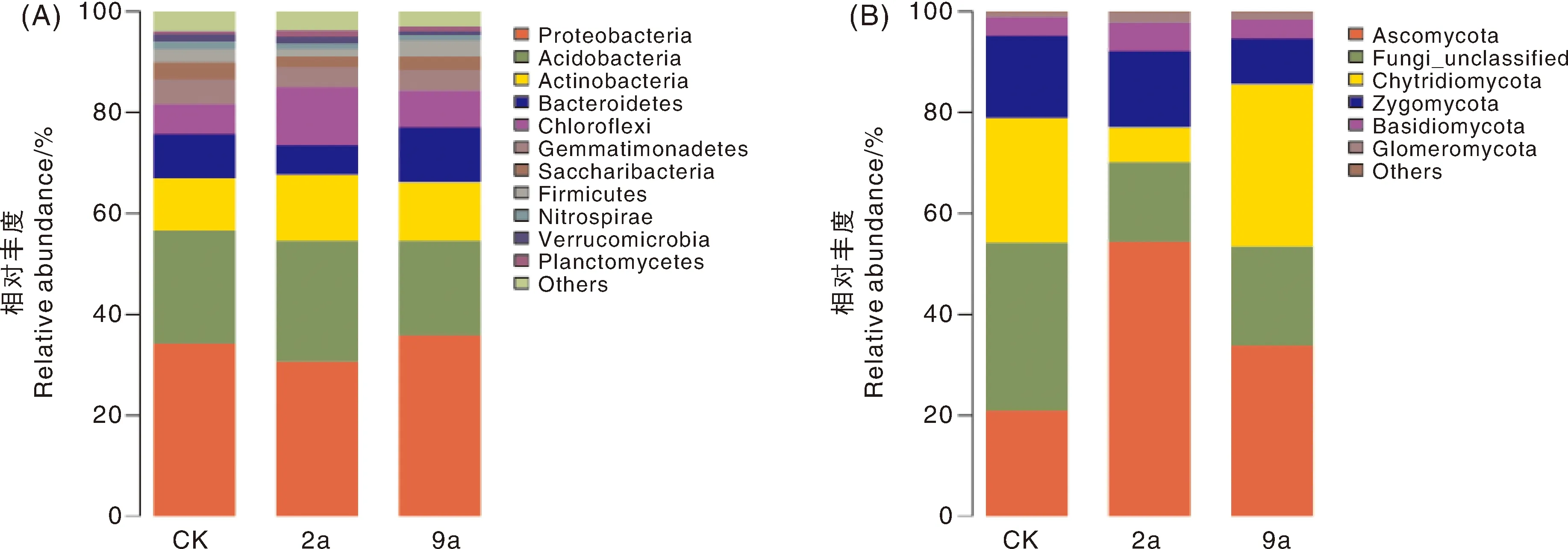
A:Proteobacteria,变形菌门;Acidobacteria,酸杆菌门;Actinobacteria,放线菌门;Bacteroidetes,拟杆菌门;Chloroflexi,绿弯菌门;Gemmatimonadetes,芽单胞菌门;Saccharibacteria,糖杆菌门;Firmicutes,厚壁菌门;Nitrospirae,硝化螺旋菌门;Verrucomicrobia,疣微菌门;Planctomycetes,扁平菌纲;Others,其他。
在属分类水平上,3个年限根际土壤细菌分属于884个属,除未鉴定的细菌属外,主要隶属于25个细菌属,占比达到47%以上。如图5-A所示,RB41、酸杆菌属、黄杆菌属、厌氧绳菌属、亚硝化单胞菌属为属水平上的优势菌。其中,RB41和酸杆菌属为3个年限共有优势菌属。与CK相比,2a、9a的酸杆菌属相对丰度分别增加75.14%、35.00%,RB41相对丰度分别降低30.20%、53.02%。厌氧绳菌属为2a、9a的优势菌属,其相对丰度比CK增加了127.45%、31.97%。3个年限根际土壤真菌分属于359个属,除未鉴定的真菌属外,主要隶属于20个真菌属,所占比例达到59%以上。如图5-B所示,CK根际土壤优势菌属有Olpidiaster、被孢霉属、Gibellulopsis,2a根际土壤优势菌属有被孢霉属、火丝菌属、Olpidiaster、明梭孢属、镰刀菌属,9a根际土壤优势菌属有Olpidiaster、被孢霉属、明梭孢属、毛壳菌属、土赤壳属。3个年限根际土壤真菌群落在属水平上的组成和优势菌属所占比例差异显著,随着种植年限的增加优势菌属被孢霉属相对丰度降低,明梭孢属相对丰度增加,Olpidiaster先降低后升高。此外,2a、9a土壤镰刀菌属、土赤壳属相对丰度增加。
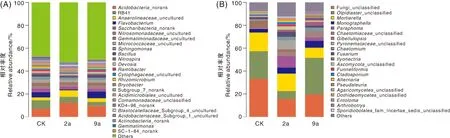
A:Acidobacteria_noran,k酸杆菌属;Anaerolineaceae_uncultured,厌氧绳菌属_未培养;Flavobacterium,黄杆菌属;Saccharibacteria_norank,糖杆菌属;Nitrosomonadaceae_uncultured,亚硝化单胞菌属_未培养;Gemmatimonadaceae_uncultured,芽单胞菌属_未培养;Micrococcaceae_uncultured,微球菌属;Sphingomonas,鞘氨醇单胞菌属;Bacillus,芽孢杆菌;Nitrospira,硝化螺菌;Devosia,德沃斯氏菌属;Ramlibacter,拉姆利氏杆菌;Cytophagaceae_uncultured,噬纤维菌属_未培养;Rhizomicrobium,根霉菌属;Bryobacter,苔藓菌属;Acidimicrobiales_uncultured,酸微菌属_未培养;Comamonadaceae_unclassified,丛毛单胞菌属_未培养;Blastocatellaceae_Subgroup_4_uncultured,酸杆菌属_亚组_4_未培养;Acidobacteriaceae_Subgroup_1_uncultured,酸杆菌属_亚组_1_未培养;Actinobacteria_norank,放线菌属;Gemmatimonas,芽单胞菌属;Others,其他。
3 讨论
3.1 连作菊花对根际土壤细菌群落结构与多样性的影响
土壤微生物的多样性和丰度反映土壤的质量状况[18],多样性指数对微生物群落变化具有指示作用[19]。本研究中,连作菊花根际土壤细菌ACE、Chao、Shannon指数升高,这与前人对大豆[20]、枸杞[21]连作后根际土壤细菌群落变化的研究结果不同,而与番茄[22]、棉花[23]、荞麦[24]、桔梗[25]的研究结果相似。随着菊花连作年限的增加,土壤细菌群落OTU呈先增加后减少的变化趋势,可能是由于连作初期(2 a),菊花根系分泌的自毒物质积累浓度较低,短期内根际土壤细菌能够利用根系分泌的营养物质生长和繁殖,进而出现短期连作后细菌群落多样性和丰富度逐渐升高的现象[15,26];而长期连作后(9 a),菊花根系自毒作用占据主导地位,根际微生态环境平衡遭到破坏,细菌群落数量在达到峰值后呈现减少趋势。
土壤细菌种类众多,功能各异,群落结构组成易受外界环境的影响[27]。本研究中,菊花连作使土壤中酸杆菌门、绿弯菌门相对丰度增加,变形菌门、放线菌门等优势菌门群落数量发生不同程度变化。张媚等[13]对连作山核桃根际土壤进行基因组测序分析,发现酸杆菌门、变形菌门、厚壁菌门等含量与土壤pH值显著相关。pH值作为土壤重要的基本理化性质之一,对土壤微生物群落结构影响较大[28-29]。Lauber等[30]研究发现,酸杆菌门丰度与土壤pH值负相关。在菊花生产中,由于施肥特性和设施特定环境的共同作用,导致其栽培土壤盐分积累,产生不同程度次生盐渍化现象,进而引起土壤pH值升高,影响群落结构组成。此外,从整个土壤系统角度,土壤酸碱度改变会引起其他土壤特性的变化,如有机质含量、营养物质有效性、土壤酶功能等,这些因素均有可能导致细菌群落结构发生改变[31]。以上结果表明,菊花连作改变了根际土壤细菌群落结构,土壤pH值升高是其变化的重要影响因素。
3.2 连作菊花对根际土壤真菌群落结构与多样性的影响
大量研究表明,连作使土壤细菌数量减少,真菌数量增多[32-34],土壤由细菌型向真菌型的转换是造成连作障碍的主要原因[35-36]。本研究中,不同种植年限的真菌丰度和多样性指数均表现先升高后降低的趋势,但变化差异不显著,说明连作菊花对根际土壤真菌群落多样性和丰度影响较小。土壤菌群结构与土壤抵抗逆境的能力密切相关[37],在土壤微生态保护方面发挥重要作用。菊花连作对其土壤真菌群落组成影响较大,在门分类水平上,随着连作年限的增加土壤中子囊菌门相对丰度增加,接合菌门、担子菌门相对丰度减少;属分类水平上,明梭孢属、镰刀菌属、土赤壳属等相对丰度增加,被孢霉属相对丰度减少,这与前人研究结果相似[38-39]。子囊菌门和接合菌门多数为腐生菌,对分解植物残体和降解土壤有机质具有重要作用[40],担子菌门对木质纤维素有较强的分解能力[41];被孢霉属真菌可通过影响土壤真菌群落组成和丰度,间接改变土壤碳、氮的转化能力及其有效性[42]。通常在有机质丰富的土壤中丰度较高。张宁[43]研究表明,明梭孢属与番茄发病率呈极显著正相关。邵慧慧等[44]在西洋参锈腐病病原菌生物特性研究中发现,土赤壳属真菌是引起西洋参锈腐病的主要病原菌,土赤壳属真菌还能引起多种植物根腐病[45-46]、褐腐病[47-48]。由此说明,菊花连作使土壤根际真菌组成发生定向改变,具有提高土壤抗逆能力和植物抗性的菌群减少[26,49-51],进而引起有益菌对有害菌的拮抗作用减弱甚至消除[52],由此导致一些病原菌群落的大量富集,在设施农业特定环境下某些一般病害逐渐演变成主要病害,加重了菊花的连作障碍,对菊花的生长发育产生不利影响。
4 结论
通过对不同连作年限菊花根际土壤微生物群落结构与多样性特征分析发现,连作显著增加了根际土壤细菌数量和多样性;细菌群落组成也发生改变,连作2 a和9 a菊花根际土壤细菌群落结构相似度较高。连作对根际土壤真菌群落多样性和丰度影响较小,但对真菌群落结构组成影响显著,在门与属分类水平上,优势菌均发生改变。整体表现为,有益菌群减少,病原菌群富集。本研究对连作土壤细菌和真菌群落的代谢功能未作深入探讨,待后续的工作中开展进一步研究。
