拟南芥N末端乙酰转移酶Naa50参与调控根细胞有丝分裂
2022-12-24冯金林席晓宇赵世凤
冯金林,席晓宇,赵世凤
(山西师范大学 生命科学学院,山西 太原 030031)
蛋白质N-末端乙酰化(N-terminal acylation,NTA)是广泛存在于真核生物中的一种蛋白质修饰方式,绝大多数真核生物的胞质蛋白会发生N-末端乙酰化修饰,但是原核生物和古细菌中很少发生[1-2]。NTA是以乙酰辅酶A作为乙酰基团的供体,通过N-末端乙酰转移酶(N-terminal acetyltransferase,NATs)的催化将乙酰基转移到蛋白质N-末端的α-氨基上,主要以共翻译方式进行乙酰化修饰,且这种修饰过程是不可逆的[3]。N-末端乙酰化修饰会影响多种蛋白质的功能。在分子水平上,蛋白质的N-末端乙酰化会影响蛋白质稳定性、抑制蛋白质转位到内质网、促进蛋白质折叠,以及调节蛋白质-蛋白质相互作用等,是维持蛋白质动态平衡的重要方式[4-8]。在细胞水平上,N-末端乙酰化修饰会影响细胞增殖发育、细胞代谢、细胞凋亡等[9-11]。此外,NTA的失调会导致人Ogden综合征、发育障碍和癌症等多种病理情况的发生,也会触发植物免疫和胁迫响应[12-13]。到目前为止,已经报道了8种不同的NATs,分别为NatA~NatH[12],其在进化上都是保守的[8]。Nats由一个催化亚基和一个或两个辅助亚基组成并且具有不同的底物特异性,可以参与调节细胞的生命活动[14-17]。
NatE复合物含有一个催化亚基Naa50,以及两个辅助亚基Naa10和Naa15[18]。在人细胞中,Naa50倾向以起始甲硫氨酸保留、并且第二个氨基酸是Leu、Ile、Phe或Trp的底物蛋白进行乙酰化修饰[19]。人Naa50通过调控姐妹染色单体的分离过程,进而参与细胞分裂的调控[20]。果蝇Naa50/San蛋白缺失引起膜翅发育缺陷,Naa50通过调控黏连蛋白Scc1与黏附素亚单位Smc3的相互作用,建立或者维持姐妹染色单体的内聚力[21-23]。Naa50的缺失会降低人细胞系和果蝇细胞内姐妹染色单体的内聚力,导致细胞发生异常有丝分裂[21]。人Naa50基因在酵母中过表达会导致N末端乙酰化水平明显增加,但是酵母Naa50基因过表达没有表现出N末端乙酰化水平增加,且敲除酵母中的Naa50基因也未表现出明显表型变化[19]。此外,Naa50对植物的生长发育也至关重要。在拟南芥中,Naa50参与调控植物免疫应答和逆境胁迫响应[24]。缺失Naa50会触发植物的免疫防御调节机制,表现出严重的发育缺陷,以及诱导的胁迫反应[24]。在前期研究中发现,敲除拟南芥Naa50基因的植株对渗透胁迫表现出超敏反应[25]。naa50-1突变体主根的生长明显受到抑制,相较于野生型表现出主根较短、根毛较早分化、细胞排列紊乱、细胞核大小不均一、幼苗严重矮化且高度不育的表型[25]。基于前期研究,猜测naa50-1突变体根短的现象可能是由于Naa50参与根尖细胞有丝分裂过程,进而调控根的正常生长。目前关于Naa50在植物生长发育方面的功能和作用机制研究较少。
H2B是组成核小体的组蛋白,分析H2B-YFP的荧光信号,可以反映不同分裂时期细胞中染色体的状态。为了进一步探究Naa50在植物根生长发育过程中的作用,本文利用H2B-YFP株系,对naa50-1突变体根中细胞分裂情况进行分析;用PI染色分析naa50-1突变体的根细胞活力,还检测了根细胞中CYCB1蛋白表达水平。研究结果进一步阐明了Naa50调控根生长发育的机制。
1 材料与方法
1.1 植物材料与试剂
拟南芥[Arabidopsisthaliana(L.)Heynh.]生态型为Columbia-0(Col-0)生态型,naa50-1突变体(CS350943)购自Arabidopsis Biological Resource Center。H2B-YFP植株、CYCB1-GUS植株为实验室前期保存。植物培养基所用MS培养基基盐购自北京中科起源科技有限公司,蔗糖购自天津市科密欧化学试剂有限公司,Phytagel、GUS染液和PI染液购自Sigma公司。
1.2 方法
1.2.1 拟南芥培养
实验用到的所有种子首先在4 ℃低温下处理3 d,之后用75%乙醇消毒1 min,1.5%次氯酸消毒10 min,再点种于MS培养基,放入光照16 h(22 ℃)、黑暗8 h(18 ℃)的植物培养箱中培养7 d,然后将幼苗移栽到含有蛭石和营养液的土壤中,并移入光照16 h(22 ℃)、黑暗8 h(18 ℃)、相对湿度为65%的培养室中培养。
1.2.2 植物根尖细胞分裂情况观察
将naa50-1突变体杂合植株与H2B-YFP植株开花后进行杂交,分别获得野生型和naa50-1纯合突变体背景下含H2B-YFP的稳定纯合株系,将纯合株系的种子在MS培养基上点种并培养6 d,在激光扫描共聚焦显微镜(Olympus FV1000)下观察幼苗根尖细胞分裂与野生型的差异。并统计有丝分裂率、染色体畸变率和微核千分率。
有丝分裂率(%)=(分裂细胞数/观察的细胞总数)×100;
染色体畸变率(%)=(染色体畸变细胞数/观察的细胞总数)×100;
微核率(%)=(微核细胞数/观察的细胞总数)×100。
1.2.3 PI染色
将在MS竖直培养基生长5 d的野生型与naa50-1突变体幼苗置于载玻片上,滴加适量10 μg·mL-1的碘化丙啶染液(propidium iodide,PI),染色50 s后,将幼苗用蒸馏水洗涤1次后置于新的载玻片上,将根尖置于载玻片中央,滴加25%甘油后盖上盖玻片,用滤纸吸去多余的甘油,在激光扫描共聚焦显微镜下观察根细胞标记的荧光信号。
1.2.4 GUS染色
将naa50-1突变体杂合植株与CYCB1-GUS植株开花后进行杂交,获得naa50-1纯合突变体背景下含CYCB1-GUS的稳定纯合株系,将纯合株系的种子在MS竖直培养基上培养5 d,然后将幼苗的根置于GUS染液中,在37 ℃恒温培养箱中避光染色过夜。之后将幼苗用蒸馏水洗涤3次,置于载玻片上,将根尖移至中央并滴加适量水合氯醛(水合氯醛∶水∶甘油体积比为8∶3∶1)透明化处理4 h后,在荧光显微镜(Olympus DP71)下观察根尖细胞有丝分裂活性。
1.3 数据统计
使用Microsoft Excel 2019软件统计实验数据。使用SPSS Statistics 17.0软件中的Duncan法进行差异显著性分析。所有实验均重复3次。
2 结果与分析
2.1 naa50突变体表型
利用H2B-YFP标记株系对Naa50在细胞分裂过程中的作用进行研究,通过观察YFP荧光信号发现,野生型植株中的主根分生区细胞排列规则,细胞核呈规则球状,大小相对均匀;而naa50-1突变体中主根分生区细胞排列无序,细胞核呈现出不同的形状,大小不均等,存在较大或较小的细胞核(图1)。细胞排列无序暗示Naa50的缺失可能造成细胞分裂方向改变,细胞核形态异常暗示Naa50的缺失可能影响了细胞分裂过程中遗传物质的平均分配。
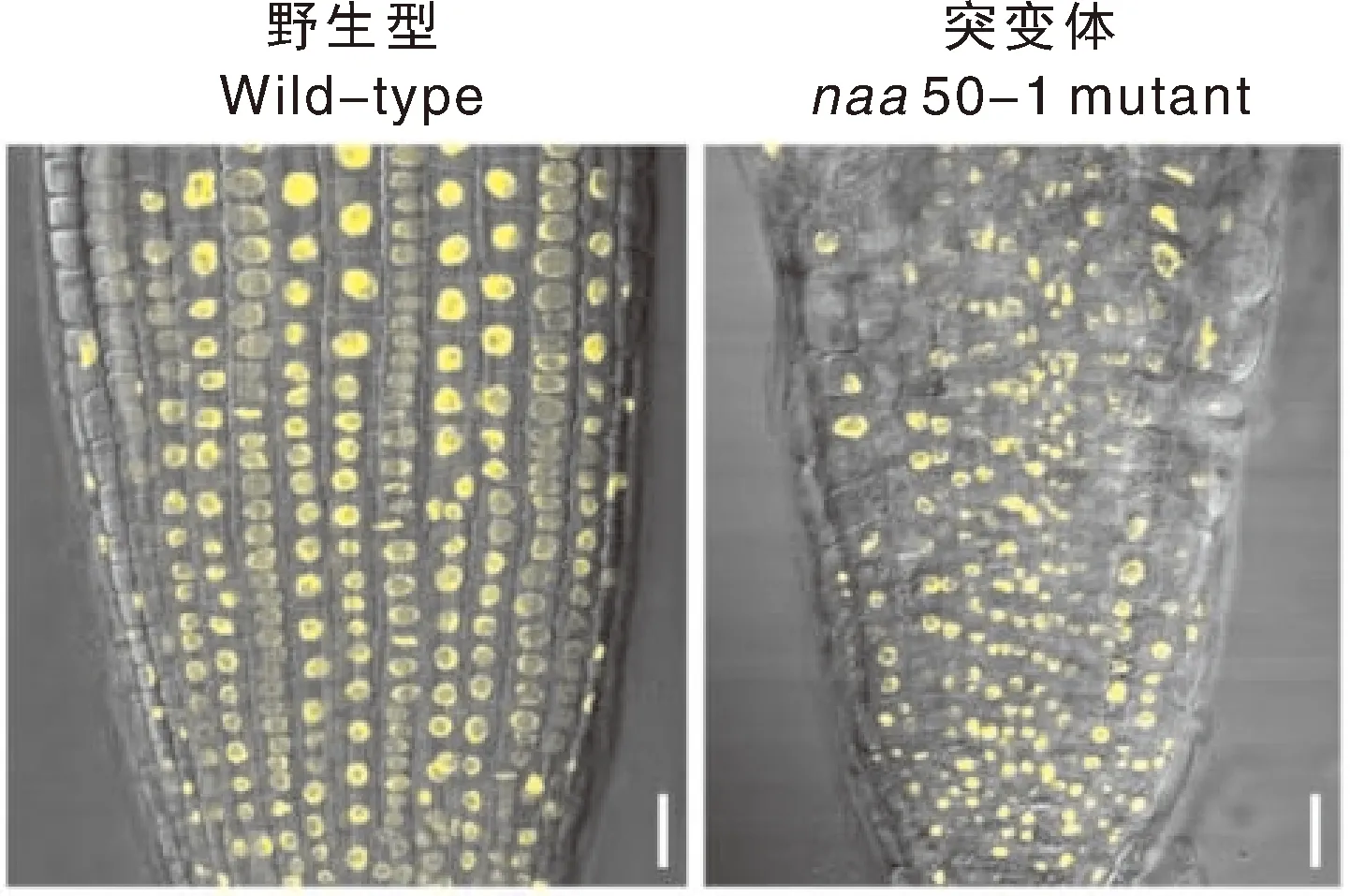
利用H2B-YFP对萌发5 d的拟南芥根尖细胞细胞核形态的观察。标尺=20 μm。
2.2 naa50-1突变体有丝分裂过程中染色体分离异常
进一步通过H2B-YFP的荧光信号观察野生型和naa50-1突变体中分裂期细胞的染色体分离行为。与野生型正常的细胞分裂过程中染色体分离行为相比,naa50-1突变体表现出异常的细胞分裂行为,包括微核、染色体桥、多束分裂、不等分裂、细胞板偏转和双核现象(图2)。naa50-1突变体的染色体畸变率和分裂中期细胞的比例高于野生型(表1、表2),表明在细胞分裂过程中Naa50对遗传信息的平均分配起重要作用,同时促进细胞分裂从中期向后期转变。
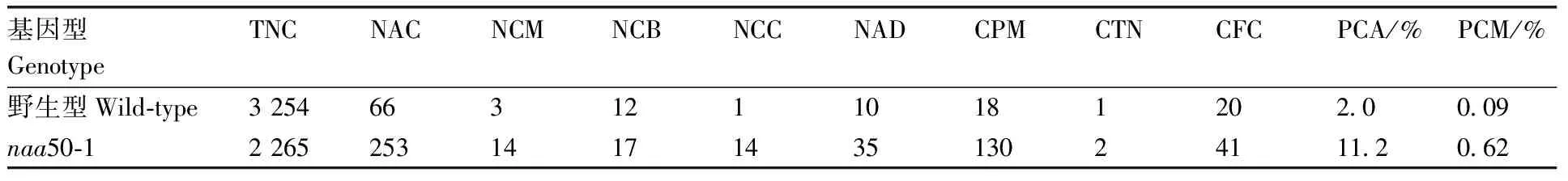
表1 Naa50对拟南芥根尖分生区细胞染色体畸变率的影响

表2 Naa50对拟南芥根尖分生区细胞有丝分裂率的影响
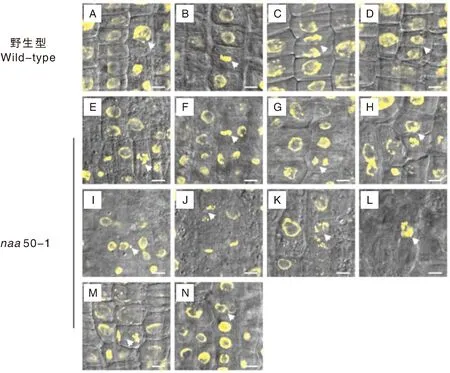
图A、B、C、D分别为野生型细胞分裂的前期、中期、后期、末期;图E、F、G、H为naa50-1突变体细胞分裂前期、中期、后期、末期;图I为微核;图J为染色体桥;图K为多束分裂;图L为不等分裂;图M为细胞板偏转;图N为双核现象。标尺=5 μm。
2.3 naa50突变体有丝分裂活性
为进一步探究Naa50在细胞分裂过程中的作用,利用细胞周期蛋白CYCB1-GUS标记株系对naa50-1突变体中的CYCB1蛋白表达丰度进行检测。GUS信号的结果表明,相较于野生型,naa50-1突变体中CYCB1的蛋白丰度明显增加(图3-A)。这可能是由于naa50-1突变体中分裂中期细胞比例增加,进而促进细胞进入分裂期,是植物本身的一种反馈响应。
2.4 naa50突变体根细胞活性
细胞的异常分裂可能会导致遗传信息无法平均分配到两个子细胞中,造成产生非整倍体的子细胞。为了进一步探究非整倍体对植物细胞的影响,对野生型和naa50-1突变体的主根进行PI染色。PI主要对野生型根细胞的细胞壁进行着色,而在naa50-1突变体中,部分细胞内部也有红色的PI染色信号,表明这些细胞已经死亡(图3-B)。以上结果暗示naa50-1突变体中出现的死细胞可能是由于非整倍体细胞的基因组不稳定,导致细胞凋亡。
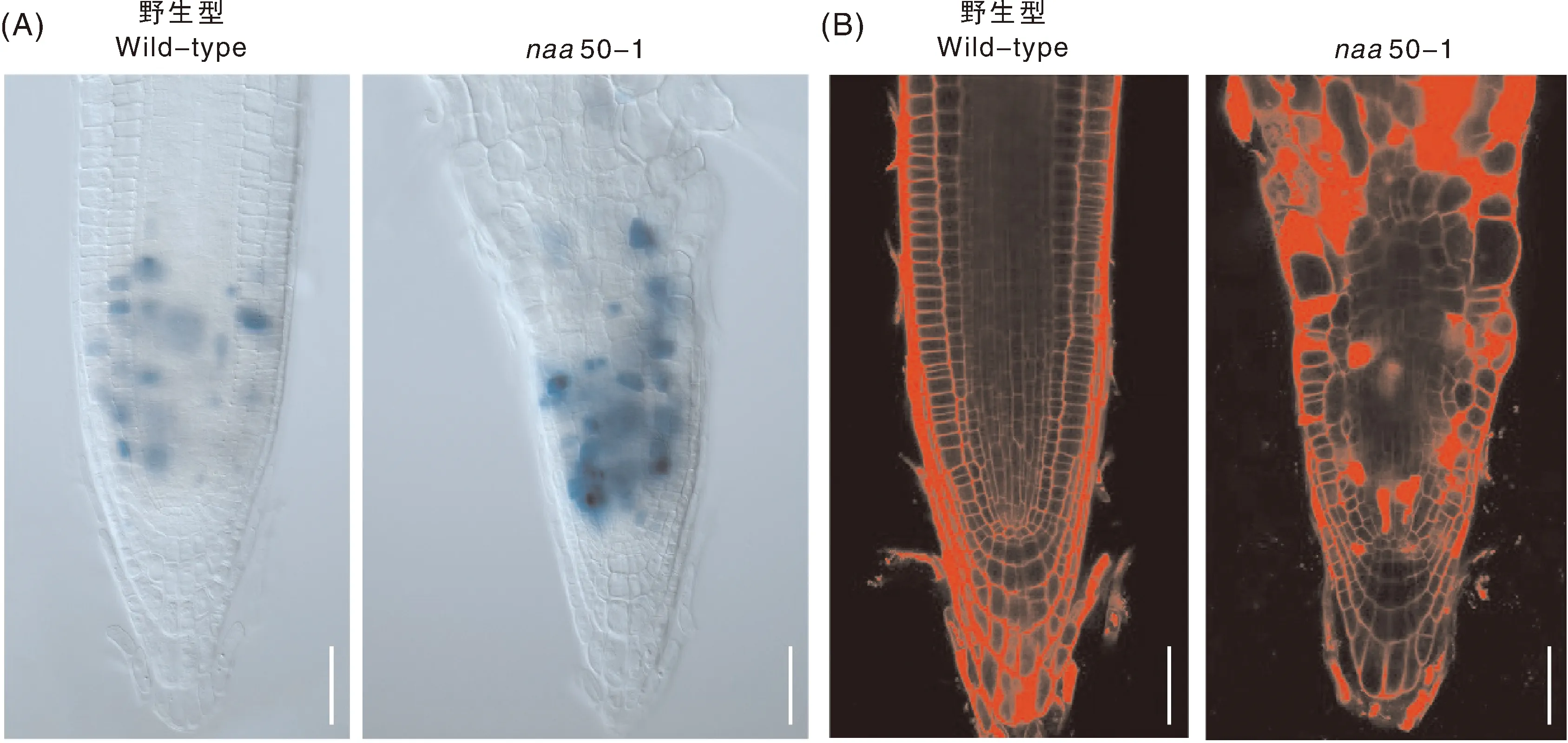
A,利用CYCB1-GUS对生长5 d的拟南芥根中CYCB1的表达量进行检测;B,对生长5 d的拟南芥根尖进行PI染色;标尺=50 μm。
3 结论与讨论
有丝分裂在细胞生长发育过程中发挥重要作用,是真核生物细胞分裂的基本方式。在有丝分裂期间,细胞骨架与膜结构重塑,纺锤体重新组装,染色体被均匀地分配到两个子细胞中[26]。正是由于多种细胞机制调控染色体的正确分离,才确保了物种之间稳定的遗传特性[27]。异常的有丝分裂会造成染色体非整倍体的产生,导致染色体不能正常分离,使子代基因组信息不完整,甚至会影响个体的生长发育。
本研究结果表明,Naa50对拟南芥根尖细胞有丝分裂至关重要,缺失Naa50会导致根细胞散乱排列,细胞核呈现出大小不均等的现象,且表现出不同程度的细胞死亡现象。另外,naa50-1突变体相较于野生型有丝分裂率下降,染色体畸变率和微核率显著增加,处在有丝分裂中期的细胞数多,细胞周期蛋白CYCB1的表达量明显增加,说明Naa50能促进细胞有丝分裂从中期向后期转移,并且促进细胞分裂过程中遗传信息平均分配到两个子细胞中。综上所述,Naa50参与调控根细胞有丝分裂,在蛋白质共翻译修饰水平上发挥作用进而影响植物的生长发育。
在真核生物中,NDE1(nuclear distribution protein nudE homolog 1)是动力蛋白Dynein的调节蛋白,敲除小鼠的Nde1基因,会造成其皮质神经前体细胞纺锤体组装紊乱和有丝分裂延迟的现象;PLK1是一种在有丝分裂期高表达的丝苏氨酸激酶,可以促进细胞有丝分裂和细胞骨架的形成,NDE1是PLK1的一种结合蛋白,两者相互作用参与调节细胞有丝分裂[26]。植物分生组织(如根尖分生区、茎尖生长点等)中连续、规则的细胞依赖于有丝分裂增加其细胞数目,促进根的生长发育,保证植物从周围土壤中吸收所需的营养成分[28-29]。拟南芥根分生组织细胞的染色体实时成像表明,有丝分裂的细胞中染色体和细胞骨架表现出高度的动态变化[29]。微管(microfllaments,MFs)和肌动蛋白微丝(actin microfilaments,AFs)是细胞骨架和细胞分裂所需的重要组成成分,分裂过程中早前期细胞分裂平面的选择、染色体向两极迁移和形成新细胞板都需要MFs和AFs的参与[29-31]。因此,MFs和AFs对细胞形态建成和细胞分裂至关重要,Naa50是否通过调控MFs和AFs而影响细胞有丝分裂也是未来非常值得关注的内容。
