棉花陆海渐渗系次级分离群体产量和纤维品质QTL 定位
2022-12-22杨芮李鹏涛肖向辉李俊文龚举武刘爱英巩万奎商海红葛群卢全伟潘境涛邓晓英范森淼石玉真袁有禄
杨芮,李鹏涛,肖向辉,李俊文,龚举武,刘爱英,巩万奎,商海红,葛群,卢全伟,潘境涛,邓晓英,范森淼,石玉真*,袁有禄*
(1. 中国农业科学院棉花研究所/ 棉花生物学国家重点实验室/ 农业农村部棉花生物学与遗传育种重点实验室,河南 安阳 455000;2. 安阳工学院生物与食品工程学院,河南 安阳 455000)
棉花是世界范围内广泛种植的重要经济作物之一,其天然纤维为全球提供关键的纺织原材料[1]。 面对日益减少的耕地和不断增长的人口数量,以及现代棉纺织工业的迅猛发展和人们对衣着品质需求的逐渐提高,同步改良产量和纤维品质性状,培育和推广优质高产的棉花品种成为越来越迫切的需求。 然而,棉花产量和纤维品质均属于复杂的数量性状,由多基因控制且易受环境因素的影响,两者表现出负相关关系[2-4],导致产量和纤维品质的同步遗传改良进展缓慢。
四大棉花栽培种中, 陆地棉(Gossypium hirsutum)和海岛棉(G.barbadense)是异源四倍体,每年贡献超过95%的棉花总产量。 陆地棉品种具有高产、适应性广等优点,但纤维品质普通;而海岛棉品种纤维品质优异,但产量低、适应性差[5]。 因此,借助染色体片段代换系(chromosome segment substitution line,CSSL) 将海岛棉优异的纤维品质基因导入陆地棉,拓宽陆地棉栽培品种狭窄的遗传基础(长期人工选择所致),为充分聚合陆海优异性状基因提供了新的方法, 也为培育高产且优质的棉花新品种奠定了材料基础[6]。作为近等基因系(near-isogenic line,NIL)[7],CSSL可降低群体遗传背景的干扰,适用于对调控复杂数量性状或多变表型的遗传位点进行精确定位,尤其是遗传效应相对较小的农艺性状,因而被广泛应用于数量性状位点(quantitative trait loci,QTL)定位和候选基因鉴定[8]。 目前,利用CSSL群体鉴定到大量与棉花产量、纤维品质、黄萎病抗性等性状相关的QTL[9-10]。
2005 年,Stelly 等[11]首次利用杂交、回交和自交等常规育种技术构建陆海渐渗系群体,获得17个陆地棉TM-1 遗传背景下含海岛棉3-79 染色体片段的种质系。 王鹏等[12]以陆地棉TM-1 和海岛棉7124 为亲本, 结合常规育种手段和现代分子标记辅助选择(marker-assisted selection,MAS)技术构建了包含330 个株系的陆海渐渗系群体,为后续开展棉花重要性状QTL 的精细定位和候选基因的克隆研究奠定了坚实的基础。 基于陆地棉TM-1 和海岛棉3-79 构建的陆海渐渗系群体,Luan 等[13]利用轮回亲本TM-1 分别与CS-B14Sh和CS-B22Sh 构建次级分离群体, 在F2和F2:3群体中鉴别到24 个与产量或纤维品质性状相关的QTL。 付央等[14]利用陆地棉TM-1 和染色体片段置换系Sub18(陆地棉TM-1×海岛棉3-79)构建了包含45 个置换系的群体,检测到与产量、纤维品质相关的7 个加性效应QTL 和5 个上位性效应QTL。 李超等[15]利用陆地棉中棉所8 号和海岛棉Pima90-53 构建包含182 个株系的陆海渐渗系群体, 挖掘到59 个与产量或纤维品质相关的QTL。Shi 等[16-17]以陆地棉中棉所36 和海岛棉海1为亲本构建了包含408 个株系的陆海渐渗系群体,结合高密度遗传连锁图谱和多年多环境的表型数据,挖掘到227 个与产量、纤维品质、黄萎病抗性等相关的QTL。 向丹[18]在以陆地棉TM-1 为背景获得了3 个海陆渐渗系材料的基础上,通过进一步组配作图群体,检测到7 个与产量、纤维品质相关的QTL。 焦梦佳等[19]选取海岛棉优异纤维渐渗系和陆地棉品种鲁棉研37 号创制F2群体, 成功鉴定出20 个与产量和纤维品质性状相关的QTL。 孙颖[20]以黄褐棉和陆地棉中棉所35为双亲构建BC3F2群体, 分别鉴定到63 个产量相关QTL、58 个纤维品质相关QTL。郝永水等[21]以毛棉和陆地棉中棉所35 作为渐渗系的亲本材料,在3 个群体中共鉴定出177 个与产量和纤维品质相关的QTL。
本研究利用高产且适应性广的陆地棉品种中棉所36(CCRI 36)和纤维品质优异、高抗黄萎病的海岛棉品系海1(Hai 1)为亲本,通过杂交、高代回交和自交构建陆海渐渗系群体BC5F3:5,基于多年多环境表型数据,选育出产量和纤维品质优良、整齐一致的陆海渐渗系MBI9626;以中棉所36 为轮回亲本构建次级分离群体; 结合多态性简单重复序列(simple sequence repeat,SSR)标记和多年多点表型数据,开展产量、纤维品质性状相关的QTL 定位分析; 借助已发表的转录组数据和生物信息分析软件, 挖掘稳定QTL 区间内的候选基因,为后续精细定位和分子标记辅助选择育种奠定基础,同时也为同步改良产量和纤维品质性状提供参考。
1 材料与方法
1.1 试验材料和田间种植
2015 年, 利用陆海渐渗系MBI9626 与轮回亲本中棉所36 配制杂交组合, 去雄授粉后获得F1。2016 年,在中国农业科学院棉花研究所(简称中棉所)农场(河南省安阳县白壁镇)种植F1产生包含152 个单株的F2群体, 用AYF2表示该群体。 2017 年,在中棉所农场和新疆石河子北泉镇综合试验站(记为“XJ”)按株行种植F2:3株行,共设置2 个重复, 分别用AYF2:3和XJF2:3表示这2个群体。 2018 年在中棉所农场种植F2:4株系,不设重复,用AYF2:4表示该群体。
1.2 遗传标记筛选和群体基因型检测
F2群体的基因组DNA 提取采用改良CTAB法[22],依据Nanodrop 检测的DNA 浓度,将3 个亲本和杂交后代全部单株的DNA 原液稀释成50 mg·L-1工作液。 基于Shi 等[5]利用中棉所36与海岛棉海1 构建的陆海渐渗系BC1F1群体绘制的长度为5 115.16 厘摩(centimorgan,cM)的高密度遗传连锁图谱,利用亲本从2 292 个SSR 标记中筛选多态性标记。 然后,用筛选出的标记对F2群体进行检测,扩增产物采用聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis, PAGE)和银染法快速检测。
1.3 数量性状的测定
F2群体按单株收取所有成熟棉铃并记录铃数,F2:3和F2:4群体分别以株行为单位收取30 个棉铃,自然风干后室内考种,分别称量籽棉和皮棉质量,计算铃重(boll weight, BW)和衣分(lint percentage,LP)。 F2群体按单株,F2:3按株行分别取皮棉样品15 g,送农业农村部棉花品质监督检验测试中心使用HFT900 测试仪按HVI 校准棉花标准校准测定纤维品质,包括纤维上半部平均长度(upper half mean length, FL)、断裂比强度(breakingtenacity,FS)、马克隆值(micronaire,FM)、长度整齐度指数(uniformity index,FU)和断裂伸长率(breaking elongation,FE)。 AYF2:4群体仅测定了铃重。
1.4 统计分析和QTL 定位
用SPSS 20.0 对各群体的性状表型值进行描述性统计及相关性分析。 用GGT32 分析渐渗系MBI9626 中渐渗片段的数量、大小及其分布。 基于Shi 等[5]构建的遗传图谱,用QTL IciMapping软件进行标记的连锁分析,结合表型数据和图谱进行单标记作图分析, 设置条件数(condition number)为1 000,对数优势比(logarithm of the odd score,LOD) 大于等于2.5。 使用Map Chart 2.2 软件绘制QTL 染色体分布图。QTL 命名方式统一采用:q+性状英文缩写+染色体编号+QTL 编号。 此外,对包含2 个及以上QTL 的区间进行分析, 如果距离在10 cM 以内则视为1个QTL 簇,QTL 簇命名规则为:Clu+染色体编号+QTL 簇编号。
选择包含重要QTL 的区段, 根据QTL 的侧翼标记查找其在陆地棉TM-1 参考基因组[1]对应的物理区间,从CottonFGD 网站(http://cottonfgd.net/)下载区间内所有基因的序列。 基于Hu 等[23]发表的陆地棉标准系TM-1 开花当天(0 day post anthesis,0 DPA)、1 DPA、3 DPA、5 DPA 的胚珠和10 DPA、20 DPA 和25 DPA 的纤维RNA-seq 数据(http://cotton.zju.edu.cn/),利 用 软 件edgeR 对区间内的基因进行差异表达基因鉴别,按照伪发现率(false discovery rate, FDR)小于0.05 和log2(fold change)大于2(fold change 是差异倍数)的过滤阈值对上述7 个样本进行两两比对。 对富集到GO 类别中前25 个生物学过程的差异表达基因(differentially expressed gene, DEG)和富集到KEGG 类别中前25 个信号通路的差异表达基因取并集后, 筛选在纤维发育过程中高表达的基因,用每个基因的表达量减去这个基因在所有样本中表达量的均值,然后除以其标准差进行归一化处理,比较基因表达水平。 利用Blast 软件比对找到相应的海岛棉基因ID,将陆地棉和海岛棉中同源基因编码的蛋白序列进行比对以确定候选基因。
2 结果与分析
2.1 产量和纤维品质性状描述性分析
利用SPSS 20.0 软件对4 个群体的2 个产量相关性状和5 个纤维品质相关性状的表型值进行描述性分析, 并以中棉所36 为对照 (表1)。AYF2分离群体的铃重、衣分、上半部平均长度、长度整齐度指数的平均值均低于中棉所36。 XJF2:3和AYF2:3群体除衣分外的6 个指标的表型值,以及F2:4群体的铃重均高于中棉所36。 各性状变异系数在1.05%~7.93%,易受环境影响,分离群体中存在明显的差异。 所有性状偏度的绝对值均小于1,表明4 个群体中均没有出现异常的偏分离。峰度的绝对值均小于1, 表明采集到的各群体的表型数据基本符合正态分布,可以用于QTL 分析。

表1 亲本及群体产量和纤维品质相关性状表型数据的统计分析Table 1 Analysis of phenotypic data of yield- and fiber quality-related traits for parents and four populations
2.2 产量和纤维品质性状相关性分析
对同一环境下不同性状间的相关性分析发现 (表2),AYF2:3和XJF2:3的铃重与衣分显著负相关,AYF2的铃重与上半部平均长度显著负相关, 铃重与其他纤维品质指标无显著相关关系。衣分与马克隆值在2 个环境中表现为极显著正相关(AYF2除外);上半部平均长度与长度整齐度指数(AYF2除外)、断裂比强度、断裂伸长率极显著正相关,与马克隆值(XJF2:3除外)极显著负相关; 断裂比强度与马克隆值极显著负相关、与断裂伸长率极显著正相关;长度整齐度指数与断裂伸长率显著或极显著正相关。
2.3 基因型分析
共筛选出109 个SSR 多态性标记,分布于除8 和18 号染色体之外的24 条染色体上。参考Shi等[5]构建的遗传图谱,渐渗系MBI9626 的遗传背景94.8%恢复为轮回亲本中棉所36, 遗传长度4 851.3 cM; 渐渗纯合片段长212.7 cM, 占比为4.2%;杂合片段长51.2 cM,占比为1.0%,明显少于纯合片段,便于后续的遗传分析。
利用筛选出的标记对AYF2群体中152 个单株进行检测, 恢复到中棉所36 背景的平均值为99.6%;海1 渐渗片段的平均长度为20.46 cM,占比为0.4%。 共有136 个单株包含海1 片段,最多有22 个片段, 平均每株有10.68 个渐渗片段;这些单株中海1 片段的长度集中于10.23~35.81 cM。
2.4 QTL 定位分析
共检测到28 个与产量和纤维品质性状相关的QTL,分布在6 条染色体上(表3,图1),17 号染色体上最多,共13 个。 共检测到16 个与产量相关的QTL, 其中7 个与铃重相关,9 个与衣分相关,表型贡献率为2.25%~6.14%,包括6 个在多年/多环境被检测到的QTL。表型贡献率大于5%的有qLP-17-2、qLP-17-3和qLP-17-4。 其中3个铃重QTL、6 个衣分QTL 的加性效应值为负值,加性效应均来源于中棉所36。
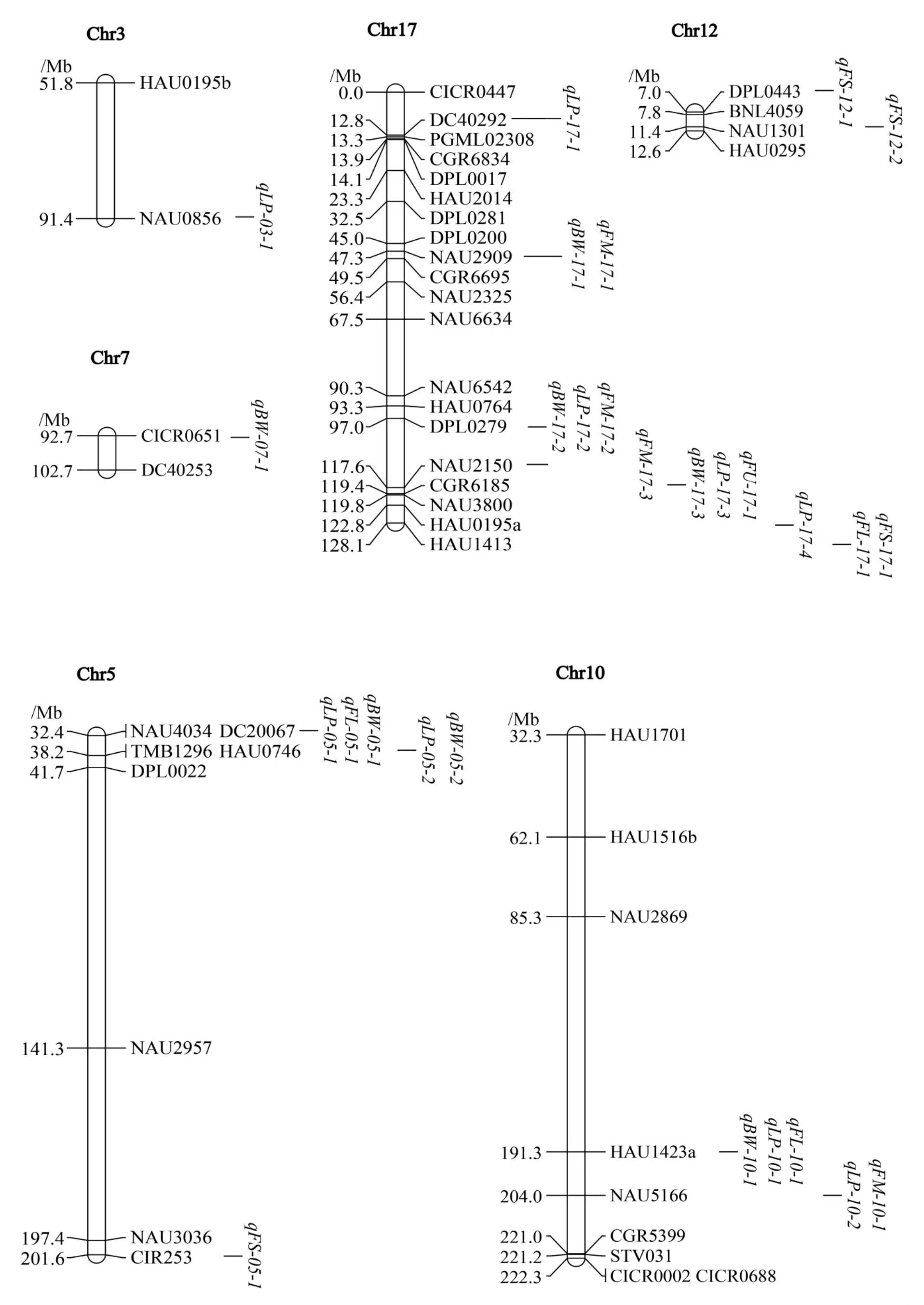
图1 在3 个群体中检测到的与产量、纤维品质相关的QTLFig. 1 QTL mapping for yield- and fiber quality-related traits in three populations
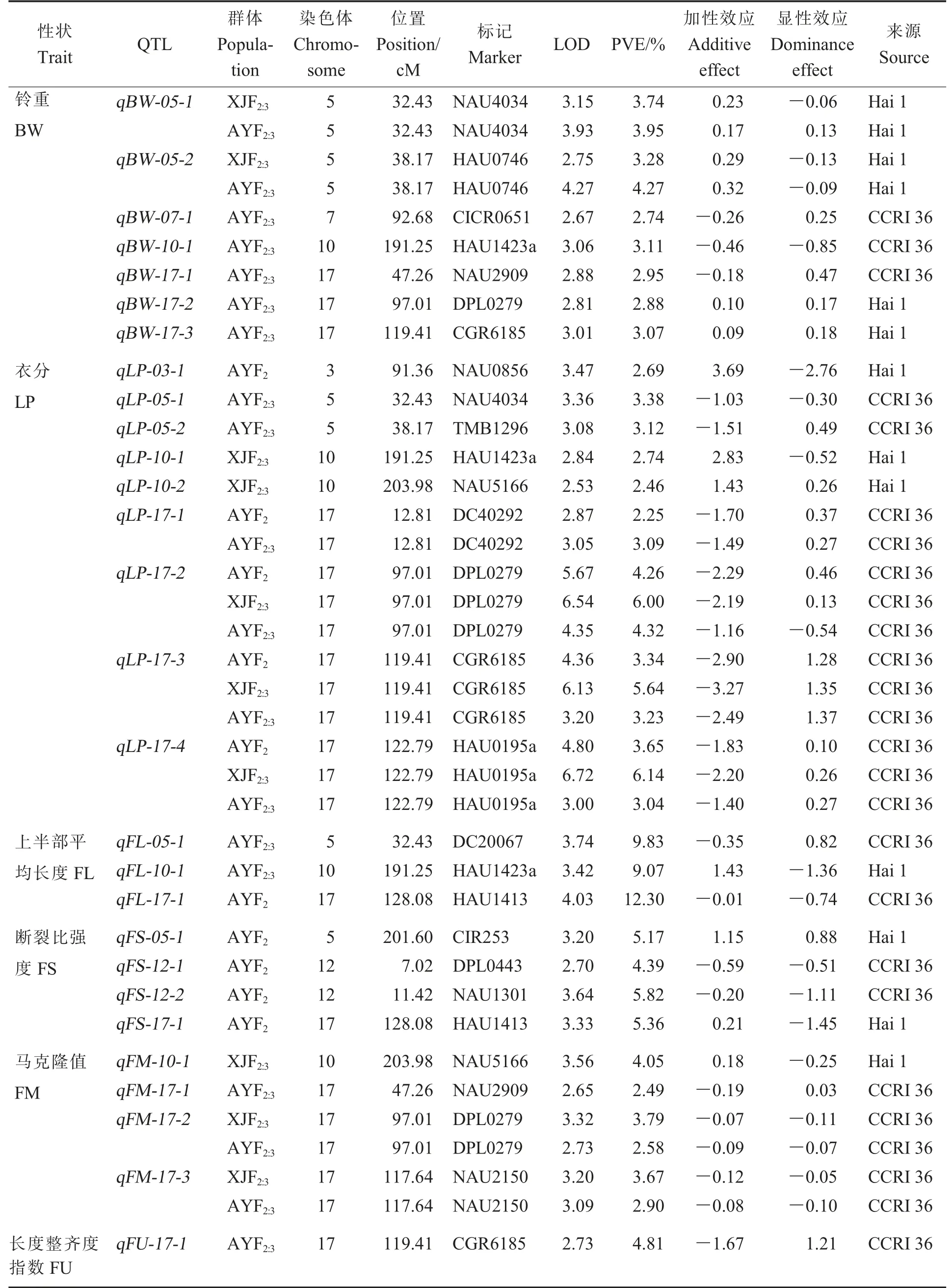
表3 在3 个群体中检测到的与产量、纤维品质相关的QTLTable 3 QTL for yield- and fiber quality-related traits in three populations
同时,检测到12 个与纤维品质相关的QTL,包括3 个上半部平均长度QTL、4 个断裂比强度QTL、4 个马克隆值QTL、1 个长度整齐度指数QTL, 表型贡献率为2.49%~12.30%; 其中qFM-17-2和qFM-17-3在2 个群体中被检测到。加性效应来自海1 的QTL 有4 个, 上半部平均长度QTL 1 个、断裂比强度QTL 2 个、马克隆值QTL 1 个。
2.5 QTL 簇的分布
分析QTL 定位结果(图1), 发现部分QTL具有成簇分布的特点,共有7 个QTL 簇分布在4条染色体上。 QTL 簇Clu-05-1 包括qBW-05-1、qLP-05-1、qFL-05-1、qBW-05-2和qLP-05-2共5个QTL,Clu-10-1 包 括qBW-10-1、qLP-10-1和qFL-10-1,Clu-10-2 包括qLP-10-2和qFM-10-1,Clu-12-1 有qFS-12-1和qFS-12-2,Clu-17-1 包括qBW-17-1和qFM-17-1,Clu-17-2 包括qBW-17-2、qLP-17-2和qFM-17-2共3 个QTL,Clu-17-3 包括qBW-17-3、qFU-17-1、qLP-17-3、qLP-17-4、qFL-17-1和qFS-17-1共6 个QTL。
2.6 候选基因的预测
鉴于Clu-17-3 中QTL 数目最多, 本研究对该QTL 簇进行深入分析。 该区间位于标记CGR6185 和HAU1413 之间,对应陆地棉标准系TM-1 参考基因组的44.1~47.8 Mb 区间内,物理长度为3.7 Mb,包含233 个基因。
随后, 基于已发表的陆地棉标准系TM-1 0、1、3、5、10、20 和25 DPA 的胚珠和纤维转录组数据,共筛选到118 个差异表达基因,并对这些基因进行GO 富集和KEGG 分析。 GO 聚类结果显示, 差异表达基因显著富集到木质部发育(GO:0010089)、气孔运动的调节(GO:0010119)、气孔运动(GO:0010118)等生物学过程(附表1),而在KEGG 聚类结果中显著富集到的信号通路主要包括内酰胺生物合成(ko00261)、磷酸肌醇代谢(ko00562)、赖氨酸生物合成(ko00300)等(附表2)。 随后,对富集到GO 类别中前25 个生物学过程的9 个差异表达基因和富集到KEGG 类别中前25 个信号通路的10 个差异表达基因取并集,筛选出17 个候选基因。
对17 个候选基因的表达模式分析发现,GH_D03G1443、GH_D03G1511、GH_D03G1526、GH_D03G1544和GH_D03G1611这5 个候选基因的表达量较低,FPKM(fragments per kilobase of exon model per million mapped fragments)均低于10,因此对其余的12 个候选基因进行序列分析。序列比对发现GH_D03G1458、GH_D03G1460、GH_D03G1511、GH_D03G1517、GH_D03G1552和GH_D03G1593这6 个基因与其在海岛棉中的同源基因编码的氨基酸序列完全一致,另外6 个候选基因GH_D03G1428、GH_D03G1466、GH_D03G1518、GH_D03G1570、GH_D03G1586和GH_D03G1640与其在海岛棉中的同源基因的编码蛋白存在序列差异,推测这6 个基因可能与纤维发育相关。
对这6 个基因的表达模式(图2)和注释信息分析, 同时参考拟南芥中同源基因信息,编码磷酸丙糖异构酶 (triosephosphate isomerase)的GH_D03G1428在0~3 DPA 胚珠中的表达量持续升高且在3 DPA 达到最大值,然后在5 DPA胚珠和10~25 DPA 纤维中的表达量逐渐降低,暗示该候选基因可能在纤维起始阶段发挥重要的作用。 编码RNA 结 合 蛋 白(RNA-binding protein)的GH_D03G1466在0~5 DPA 胚珠和10~25 DPA 纤维中的表达量呈波浪式上调、下调,且在1 DPA 和20 DPA 分别达到最高值和第二高值,暗示该候选基因可能在纤维发育起始阶段和次生壁加厚期发挥重要的作用。编码3-磷酸甘油醛脱氢酶(glyceraldehyde-3-phosphate dehydrogenase-like family protein)的GH_D03G1518在0~5 DPA 胚珠和10~25 DPA 纤维中的表达量不断升高且在20 DPA 纤维中达到最大值,在25 DPA 纤维中表达量降低, 暗示该候选基因可能在纤维发育起始和伸长阶段发挥重要的作用。 编码重金属转运/ 解毒超家族蛋白(heavy metal transport/detoxification superfamily protein)的GH_D03G1570的表达量由0 DPA 纤维中的最高值逐渐降低,随后先上升、再下降、又上升,暗示该候选基因在纤维发育起始阶段发挥重要的作用。 编码CHY 型/ 环型锌指蛋白(CHY-type/RING-type zinc finger protein)的GH_D03G1586在0~3 DPA 胚珠中的表达量逐渐升高,在5 DPA 胚珠中的表达量降低,此后在10~25 DPA 纤维中的表达量逐渐由最高降至最低,暗示该候选基因可能在纤维发育起始阶段和伸长与次生壁加厚期发挥重要的作用。 编码盐超敏感互作蛋白(salt overly sensitive interacting protein,SOS3) 的GH_D03G1640在0~5 DPA 的胚珠及10~25 DPA 的纤维中的表达量呈波浪式上调和下调交替变化,且在1 DPA 胚珠中的表达量最高,暗示该候选基因可能在纤维发育起始阶段发挥重要的作用。

图2 6 个候选基因在陆地棉TM-1 胚珠和纤维中的表达热图Fig. 2 Expression heatmap of six candidate genes in ovule and fiber of upland cotton TM-1
3 讨论
3.1 染色体片段代换系群体适于QTL 定位
陆地棉和海岛棉共同起源、独立进化,有明显的基因组分化,二者在植株形态、产量、纤维品质等表型方面差异明显[23]。 本研究中利用陆地棉和海岛棉杂交构建的BC6F2、BC6F2:3和BC6F2:4群体,其产量及纤维品质性状存在广泛变异,表型值符合正态分布。CSSL 群体属于永久分离群体,变异位点以纯合基因型为主, 遗传组成相对稳定,遗传力高[24]。 本研究构建的CSSL 群体中,单株的遗传背景恢复率平均值为99.6%。 这类遗传背景清晰的群体适用于QTL 定位分析[25],即连锁分析的QTL 结果不容易受群体结构的影响,直接测定后续世代的基因型和表型,通过两者的相关性可以确定控制产量和纤维品质性状的基因在染色体上的位置及其遗传学效应。已经在番茄[26]、水稻[27]、小麦[28]、玉米[29]等作物中证实CSSL能提高QTL 分析的准确性。
3.2 稳定QTL 的鉴定
分子育种是当前作物遗传育种的重要研究方向, 产量和纤维品质性状的QTL 定位有助于解决棉花产量和纤维品质同步遗传改良难度大的问题[30]。 因此,多环境稳定的QTL 鉴定是后续精细定位和分子标记辅助选择育种的基础。 通过陆地棉种内杂交和陆海群体构建获得的QTL 定位结果可以通过比较基因组学分析很好地整合在一起[31]。 不少来源于野生棉、海岛棉等的主效QTL 已被检测出来,但在不同的群体中重复被鉴定的QTL 数量并不多。 本研究利用陆海优质渐渗系MBI9626 为父本、陆地棉品种中棉所36 为母本构建次级分离群体F2、F2:3和F2:4进行产量和纤维品质性状的QTL 定位。 稳定的QTL 能够在多年/ 多环境中同时被检测出来,本研究定位到的28 个QTL 中,有5 个QTL 可以在2 个环境中同时被检测到, 分别是qBW-05-1、qBW-05-2、qLP-17-1、qFM-17-2和qFM-17-3, 有3 个QTL可以在3 个环境中同时被检测到, 分别是qLP-17-2、qLP-17-3和qLP-17-4。
与前人利用陆海渐渗系进行QTL 定位的研究比较,发现qBW-05-2与梁燕[32]在BC5F2群体中检测到的qBW-1-22均位于5 号染色体与标记HAU0746 紧密连锁,增效基因均来自海岛棉,与李骏智[33]在BC2F1群体中检测到的qBW-33-1一样。qBW-17-1和何蕊[34]在BC5F3:5群体中检测到的qBN-17-6均与NAU2909 紧密连锁,增效基因都来源于陆地棉。qBW-17-3与Shi 等[17]在BC1S1群体检测到的qBW-C17-2均位于17 号染色体且与标记CGR6185 紧密连锁,增效基因均来自于 海 岛 棉。qLP-03-1与 何 蕊[34]在BC5F3:4群 体中检测到的qLP-3-1均位于3 号染色体且与NAU0856 紧密连锁,加性效应值均为正值,增效基因来源于海岛棉。qLP-05-2与宋威武[35]在F2群体中检测到的qLP-5-1均位于5 号染色体与TMB1296 紧密连锁,增效基因来源于陆地棉。qLP-17-3与Shi 等[5]在BC1S1群体中检测到的qLP-C17-5均位于17 号染色体与相同标记CGR6185 紧密连锁, 增效基因均来自于陆地棉。qLP-17-4与梁燕[32]在BC5F2群体中检测到的qLP-14-4均位于17 号染色体, 且与标记HAU0195a 紧密连锁,增效基因来源于陆地棉。qFS-17-1与李邵琦[36]在F2群体中检测到的qFS-17-2均位于17号染色体与标记HAU1413 紧密连锁, 加性效应方向一致, 增效基因来自海岛棉。qFM-17-1与Shi 等[24]在BC1S1群体中检测到的qFM-C17-2均位于17 号染色体与标记NAU2909 紧密连锁,增效基因均来自于陆地棉,同时也与Zhai 等[37]在F2群体中检测到的qFM-17-7结果一致。 这9 个QTL 位点与可能与上述已经报道的QTL 为同一QTL。本研究中鉴定的另外19 个QTL 未见报道,是新检测到的QTL。 综合前人研究结果,本研究中获得14 个多环境稳定的QTL, 为候选基因的克隆验证及产量和纤维品质性状的遗传改良奠定了重要基础。
3.3 增效基因的来源
本研究利用陆海渐渗系定位了7 个控制铃重的QTL 和9 个控制衣分的QTL, 其中4 个铃重QTL 和3 个衣分QTL 加性效应值为正值,其增效基因来自海岛棉,表明控制铃重和衣分的增效基因并非总来自高值亲本中棉所36,也可以来自低值亲本海1。 纤维品质相关QTL 中qFL-10-1、qFS-05-1、qFS-17-1和qFM-10-1的增效基因来自海1,另外8 个纤维品质相关QTL 的增效基因来自中棉所36,即控制纤维品质的增效基因有的来自高值亲本海1, 有的来自低值亲本中棉所36,与前人研究结论[38-41]一致。 因此,优异性状基因的挖掘不能忽视低值亲本。
3.4 QTL 簇和候选基因的比较
本研究共发现7 个QTL 簇。 相关性分析中,铃重与衣分负相关, 它们相关的4 对QTL(qBW-05-1 与qLP-05-1、qBW-05-2与qLP-05-2、qBW-10-1与qLP-10-1、qBW-17-2与qLP-17-2以及qBW-17-3与qLP-17-3加性效应来自不同亲本,育种工作中应注重这类QTL 的应用。 衣分与马克隆值正相关, 它们相关的2 对QTL(qLP-10-2与qFM-10-1、qLP-17-2与qFM-17-2)加性效应方向相同,海岛棉的渐渗片段能够同时增加纤维的衣分和马克隆值。QTL 簇的加性效应方向与性状的相关性一致,该结果与Zhang 等[42]的研究结果一致,为产量和品质的同步改良提供了参考。
本研究中QTL 簇Clu-17-3 包括6 个QTL,该区间内存在233 个基因,预测其中的6 个候选基因可能与纤维发育相关。GH_D03G1428编码棉花磷酸丙糖异构酶, 在拟南芥幼苗萌发过程中,磷酸丙糖异构酶能动员种子中存储的营养物质为幼苗提供能量和碳源,直到自养生长成为可能[43]。GH_D03G1466在拟南芥中的同源基因编码裂解酶(alginate lyase,ALY),具有将mRNA 从细胞核转运到细胞质进行翻译的功能。 拟南芥aly1突变体在全基因组水平的甲基化、 转座子转录抑制和启动转基因沉默方面存在缺陷,研究发现ALY1 可与细胞骨架和运动蛋白功能相关[44];此外,ALY1 蛋白与RNA 解旋酶相互作用,发挥高效的mRNA 核质转运功能确保拟南芥正常生长发育[45]。 棉花GH_D03G1518与拟南芥中质体甘油醛-3- 磷酸脱氢酶(plastid glyceraldehyde-3-phosphate dehydrogenase,GAPCp)基因具有较高同源性,该酶在拟南芥花药中的表达和催化活性都是花粉发育所必需的,GAPCp 的缺乏导致雄性不育[46];另一研究发现,拟南芥中GAPCp 的下调会导致植物体内糖和氨基酸平衡发生剧烈变化,导致根系发育停滞和不育,突变体根系发育受阻归因于丝氨酸的缺乏, 根中GAPCp 的主要功能是为丝氨酸的生物合成提供前体[47]。GH_D03G1570在拟南芥中的同源基因中编码重金属相关异戊二烯基植物蛋白(heavy metal-associated isoprenylated plant protein,HIPP),通过调节拟南芥内质网中细胞分裂素降解蛋白的含量调控细胞分裂素信号活性,从而控制细胞增殖和分生组织分化的发育过程,细胞分裂素的内稳态和信号应答也对HIPP基因具有反馈调控机制[48]。 另外,有研究表明,上游调控因子HIPP3过表达会抑制拟南芥中水杨酸盐的反应和水杨酸靶基因的表达, 并刺激茉莉酮酸酯通路靶基因的表达,促进种子的发育[49]。GH_D03G1586在拟南芥中的同源基因编码环型锌指蛋白1(CHY zinc-finger and RING ptotein1 , CHYR1),属于泛素连接酶,响应脱落酸和干旱胁迫的信号转导, 在拟南芥发育和干旱响应中起积极作用,其环型结构域被磷酸化后,泛素连接酶的活性会随之增强[50]。GH_D03G1640与拟南芥SOS3基因同源, 后者是维持胞内Na+和K+稳态和拟南芥对高Na+、低K+环境的耐受性所必需的,在调节细胞质酶的活性和维持膜电位、渗透调节方面至关重要[51];另有研究发现,在轻度盐胁迫下,SOS3基因通过增加拟南芥的芽源生长素和由上往下的生长素运输活性参与维持局部生长素最大值,而生长素在根中柱鞘细胞中的局部积累会触发侧根器官发出指示信号,确定起始细胞身份[52]。这些基因在纤维发育过程中的功能有待进一步验证。
4 结论
本研究共检测到28 个与产量、 纤维品质性状相关的QTL,分布在6 条染色体上,集中在5号和17 号染色体上。 与产量相关的QTL 有16个,解释2.25%~6.14%的表型变异,与纤维品质相关的QTL 有12 个, 解释2.49%~12.30%的表型变异。 本研究检测到9 个与前人结果一致的QTL,其中有3 个多环境稳定的QTL;另外19 个QTL 中包含5 个多环境稳定的QTL, 可为棉花分子标记辅助选择育种工作提供借鉴及依据,也为后续进一步开展QTL 精细定位奠定基础。在17 号(D3)染色体的1 个QTL 簇中预测了6个可能与纤维发育相关的基因GH_D03G1428、GH_D03G1466、GH_D03G1518、GH_D03G1570、GH_D03G1586和GH_D03G1640, 为候选基因的鉴定提供了基础。
