棉花陆海杂交亲本及子代光合叶绿素荧光参数分析
2016-07-23王志军谢宗铭董永梅
王志军++谢宗铭++董永梅
摘要:以海岛棉(新海22号)、陆地棉(自育品系589)和以其为亲本配置的正反交杂种F1、F2为试验材料,大田自然环境条件下,测定花铃期倒四叶的叶绿素含量、光合参数、叶绿素荧光参数,结果发现,海岛棉的叶绿素a、叶绿素b、总叶绿素含量、Fm、ΦPSⅡ、Fv/Fm显著高于陆地棉(P<0.05);胞间CO2浓度(Ci)、Fo、Y(NO)、Y(NPQ)显著低于陆地棉(P<0.05);叶绿素a/b、蒸腾速率(Tr)、气孔导度(Gs)、光合速率(Pn)、水分利用效率(WUE)、ETR、qP、NPQ差异不显著;陆海杂交F1在Fm、NPQ、Y(NPQ)、Fv/Fm等荧光参数表现出中亲优势,叶绿素a含量、叶绿素b含量、总叶绿素含量、蒸腾速率(Tr)、气孔导度(Ci)、净光合速率(Pn)、电子传递速率(ETR)表现出超亲优势,其余参数均没有表现出杂种优势;陆海杂交F2的大部分光合叶绿素荧光参数表现出不同程度的优势退化,变化较为复杂,这可能与陆海杂交F2代性状疯狂分离有关。通过从光合生理方面挖掘陆海杂交后代潜在优势,以期探明陆海杂交后代光合及叶绿素荧光参数的杂种优势表现。
关键词:海岛棉;陆地棉;光合作用;叶绿素荧光参数
中图分类号: S562.03文献标志码: A文章编号:1002-1302(2016)05-0134-04
水稻、玉米、油菜等农作物的杂种优势利用,已取得了巨大的经济效益和社会效益,棉花杂种优势是提高棉花产量、改善纤维品质、增强抗逆性的一种有效途径。陆地棉产量高、适应性广,但纤维品质性状不足,海岛棉纤维品质优良,但生育期较长、产量低[1],棉花海陆杂种表现出巨大的杂种优势:产量高、铃多而大、品质好。张小全等研究认为海陆种间杂交在株高、果枝数、果节数、单株结铃数显著高于陆陆杂交[2]。菲力申等研究认为陆海杂种F1代较亲本在产量和品质性状均具有超亲优势[3-4]。石玉珍等研究认为海陆杂交种纤维长度、伸长率、整齐度和强度等性状均具有正向优势[5]。李哲等研究认为,海陆杂交F1代纤维长度、比强度、马克隆值3大品质性状均可达到或超过海岛棉亲本[6]。对于海岛棉及陆海杂交棉的光合及叶绿素荧光参数的研究也有较多报道。邓仲萀等研究认为,陆海杂种的光合速率介于双亲之间,不表现杂种优势[7]。姜磊等研究认为,棕色棉与白色棉杂交F1的净光合速率也没有表现出明显的杂种优势[8]。伍维模等研究认为,吐絮期陆地棉对高光强光能的利用能力强于海岛棉,而对低光强光能的利用能力弱于海岛棉[9]。关于棉花海陆杂种优势的研究多集中在农艺性状的优势表现,而对于光合特性和叶绿素荧光参数杂种优势表现的研究仍相对较少,本研究将对陆海杂交种光合叶绿素荧光参数的杂种优势表现进行深入研究。
本研究将以海岛棉、陆地棉为亲本,测定了亲本与正反交杂种F1、F2代,花铃期倒四叶的叶绿素含量及光合-叶绿素荧光参数,探讨陆海杂交后代光合性能及叶绿素荧光特性的杂种优势表现,以期从光合生理方面挖掘海陆杂交后代的潜在优势,进一步为棉花利用杂种优势提供理论依据。
1材料与方法
1.1植物材料
试验材料为海岛棉(新海22号)、陆地棉(自育品系589)和以其为亲本配置的正反交后代F1代和F2代,于2013年5—11月在新疆农垦科学院生物技术研究所试验地(45°19′ N,86°03′ E,10 ℃以上活动积温为3 570~3 729 ℃,7月份平均气温23.9~25.5 ℃)进行,采用随机区组设计,行长5 m,膜下滴灌栽培,膜宽2.05 m,1膜3管(滴灌带)6行,株距10 cm,行距20 cm,每材料播种2行,3次重复,田间管理同大田,灌水方式为有压滴灌。
1.2测定方法和参数
1.2.1光合色素测定当棉花发育至花铃期,于09:30取倒四叶,每个材料取3张叶,叶绿素的提取与测定采用高俊凤的方法[10] 。
1.2.2光合参数随机选取各材料长势一致的植株挂牌标记,晴天11:00—13:00,使用德国WALZ公司的GFS-3000光合仪,利用自然光强,大气CO2浓度约为(370±10) μmol/mol,叶室温度25 ℃,快速测定标记植株倒四叶的净光合速率[Pn,μmol/(m2·s)]、气孔导度[Gs,mmol/(m2·s)]、胞间CO2浓度(Ci,μmol/mol)等光合参数、每个材料测10株,分别取平均值。
1.2.3叶绿素荧光参数选择晴天09:30—13:00 利用德国WALZ公司的MINI-PAM荧光仪,测量上述标记叶片的叶绿素荧光参数,先暗适应30 min,测定暗适应下的初始荧光(Fo)、最大荧光(Fm)得到Fv/Fm,此时打开光化光,可测定对应光强下叶片的实际光量子产量ΦPSⅡ及其他所有的荧光参数,每个材料测10张叶,取平均值。
1.2.4杂种优势计算
中亲优势率=F1-MPMP×100%;
超亲优势率=F1-HPHP×100% 。
式中:F1为F1代杂交种观察值;MP为双亲平均值;HP为高值亲本观察值。
2结果与分析
2.1光合色素
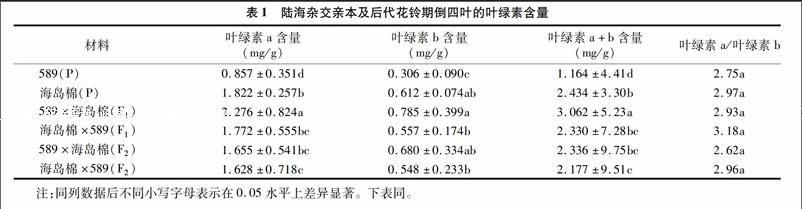
由表1可知,589为陆地棉亲本,海岛棉新海22号为海岛棉亲本,589×海岛棉(F1)为正交F1,海岛棉×589(F1)为反交F1,589×海岛棉(F2)为正交F2,海岛棉×589(F2)为反交F2。海岛棉的叶绿素a、叶绿素b、总叶绿素含量均显著高于陆地棉品系589(P<0.05),589分别是新海22号的47%、50%、48%。正交F1叶绿素a、叶绿素b、总叶绿素含量的中亲优势率为70%、71%、70%,超亲优势率分别为25%、28%、26%。反交F1中亲优势率分别为32%、21%、30%,超亲优势率分别为-3%、-9%、-4%,正交F2的叶绿素a、叶绿素b、总叶绿素含量分别比F1降低37.52、15.44、31.08百分点,显著高于589,但显著低于海岛棉(P<0.05),反交F2的叶绿素a、叶绿素b、总叶绿素含量与F1代差异不显著,分别降低885、1.64、7.03百分点,但高于父本589,低于海岛棉,差异均显著(P<0.05)。说明海岛棉较陆地棉叶绿素含量高,海岛棉做父本时,杂种F1代在叶绿素含量方面有较强的杂种优势,而且正反交之间差异显著,F2代时,杂种优势退化显著,正反交F2之间差异不显著。亲本及杂种后代的叶绿素a/叶绿素b值差异均不显著(P<0.05)。
2.2光合参数
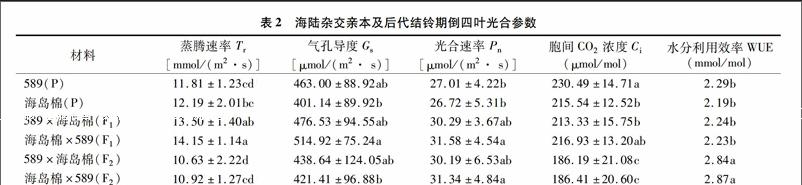
由表2可知,陆地棉589和海岛棉新海22的蒸腾速率(Tr)、净光合速率(Pn)、气孔导度(Gs)、水分利用效率(WUE)等参数无显著差异;胞间CO2浓度(Gs)海岛棉是陆地棉的93.51%,差异显著(P<0.05)。正反交F1之间光合参数及WUE均无显著差异(P<0.05)。正交F1的Tr显著高于双亲,中亲优势率为12.50%,超亲优势率为10.74%,Gs差异均不显著,中亲优势率10.30%,超亲优势率为2.9%,Pn中亲优势率为12.75,超亲优势率为12.21%,Gs较海岛棉升高但差异不显著,较陆地棉亲本显著降低,是陆地棉亲本(589)的73.44%,其他参数与双亲均没有显著差异(P<005)。反交F1的Tr显著高于双亲,中亲优势率为17.92%,超亲优势率为16.08%,Gs的中亲优势率为 19.18%,超亲优势率为11.21%,Pn中亲优势率为17.55%,超亲优势率为16.92%,Ci,WUE与双亲差异均不显著(P<0.05)。正交F2的Tr比 F1降低26.99百分点、比两亲本降低且差异显著,Gs比F1下降8.64百分点,比陆地棉亲本降低,比海岛棉亲本升高,差异均不显著,Ci比F1下降14.58百分点,比两亲本均降低且差异显著,Pn和F1差异不显著,比亲本升高,差异不显著,WUE比F1增加26.78百分点,比两亲本显著升高。反交F2的Tr比F1降低29.58百分点,比两亲本降低,Gs比F1下降22.19百分点,比陆地棉亲本降低,但比海岛棉亲本升高,差异均不显著、Ci比F1下降16.37百分点,比两亲本均降低且差异显著,Pn和F1差异不显著,比亲本升高,差异显著,WUE比F1增加28.70%,比两亲本显著升高(P<0.05)。
2.3叶绿素荧光参数
Fo为最小荧光,当光系统Ⅱ的所有反应中心均处于开放态时得到Fo。表3亲本之间差异显著,海岛棉是589的8106%,正反交F1之间差异不显著,正交F1中亲优势率为5.69%,负超亲优势率为18.03%,反交F1中亲优势率为-6.79%,负超亲优势率为4.1%,正交F2比F1降低8.19百分点,差异不显著,较陆地棉亲本降低,差异不显著,较海岛棉亲本显著升高,反交F2比F1显著升高,增幅达23.23百分点,较两亲本均升高。
Fm为暗适应样品的最大荧光,当光系统Ⅱ的所有反应中心均处于关闭状态时测得Fm,亲本之间差异显著,589是海岛棉的84.90%,正反交F1之间没有显著差异。正交F1中亲优势率为8.61%,超亲优势率为0.41%,反交F1中亲优势率为1.18%,负超亲优势率为6.46%,正交F2较F1下降达1415百分点,较陆地棉亲本升高,比海岛棉亲本降低,差异均不显著,反交F2较F1显著增高,增幅13.38百分点,较两亲本均增高。
ΦPSⅡ是光系统Ⅱ的实际量子产量,亲本之间差异显著,589是海岛棉的88.36%,正反交F1之间没有显著差异。正交F1中亲优势率为-0.59%,超亲优势率为-6.82%,反交F1中亲优势率为2.33%,超亲优势率为-3.77%,正反交F2之间也没有显著差异,正交F2较F1下降3.31百分点,差异不显著,较亲本589升高,较海岛棉下降,反交F2较F1降低105百分点,差异也不显著,较589升高,较海岛棉下降。
ETR及相对电子传递速率,在光化光强为 1 116 μmol/(m2·s) 下测定,亲本之间没有显著差异,589是海岛棉的87.05%,正反交F1之间也没有显著差异,正交F1中亲优势率为21.66%,超亲优势率为15.52%,反交F1中亲优势率为24.21%,超亲优势率为16.17%,正反交F2之间差异显著,正交F2是反交的184.48%,正交F2较F1显著升高16.30百分点,较双亲显著升高,反交F2较F1下降62.03百分点,较双亲下降,但差异不显著(P<0.05)。
qP是光化学淬灭系数,由光合作用引起的荧光淬灭,反映了光合活性的高低。由表3可以看出,亲本之间qP差异不显著,589是海岛棉的98.65%,正反交F1之间差异也不显著,正交F1中亲优势率为-1.09%,超亲优势率为-1.77%,反交F1中亲优势率为0.05%,负超亲优势率为0.63%,正反交F2之间差异显著,正交F2较F1降低2.27百分点,较双亲较低,反交F2较F1上升1.46百分点,较双亲增高。
NPQ为非光化学荧光淬灭,反映了植物耗散过剩光能为热的能力,是衡量植物光保护能力的重要指标。由表3可知,亲本之间NPQ差异不显著(P<0.05),海岛棉NPQ是589的3571%,正反交F1代差异也不显著,正交F1中亲优势率为49.47%,超亲优势率为1.43%,反交F1中亲优势率为1759%,负超亲优势率为20%,正反交F2之间差异不显著,正交F2较F1下降12.68百分点,较亲本589降低,较海岛棉亲本升高,反交F2较F1增高39.13百分点,较双亲升高。
Y(NO)为非调节性能量耗散,是光损伤的重要指标,亲本之间差异显著,海岛棉是589的71.9%,正反交F1之间差异不显著,正交F1中亲优势率为-0.64%,负超亲优势率为14.6%,反交F1中亲优势率为-7.8%,负超亲优势率为208%,正交F2较F1升高11.11百分点,较亲本589降低,较海岛棉升高,反交F2和F1无显著差异,较亲本589降低,较海岛棉升高。
3结论与讨论
海岛棉的品质优于陆地棉,纤维长、细度高、强度大,是重要的纺织品原料[7]。陆地棉皮棉产量高,但品质较差,陆地棉与海岛棉种间的杂种一代,可综合双亲的优良性状,利用其杂种优势,在保持高产的同时,可提高纤维品质,被认为是一条可行的途径[12]。
叶绿素含量与光合作用密切相关,是衡量光合作用强弱的重要因素[13-14]。植物叶绿素含量及叶绿素a/叶绿素b值变化是植物本身的遗传特性[15],同时在一定程度上受环境条件的影响[16-18]。本研究结果表明,花铃期海岛棉(新海22号)的叶绿素a、叶绿素b、总叶绿素含量均高于陆地棉(589),这与张亚黎等研究发现海岛棉新海22光合色素含量高于陆地棉新陆早13的结果[15]一致。陆海杂交F1表现出超亲优势,优势率为25%~28%,海陆杂交F1叶绿素a、叶绿素b、总叶绿素含量高于陆地棉但低于海岛棉,正反交之间有显著差异(P<0.05)正反交F2优势退化明显,略高于陆地棉亲本,但显著低于海岛棉,说明以海岛棉做父本,陆海杂交F1代叶绿素含量具有明显优势。
已有研究表明,海岛棉叶片的实际光合能力低于陆地棉,但两者具有相似的光合潜力[19]。张亚黎等研究发现,早晨和下午外界光强较低时,海岛棉新海22号和陆地棉新陆早13号的Pn无明显差异外,其余时间段新海22号叶片的Pn值显著低于新陆早13号。本研究结果表明,陆地棉589Pn、Gs、Ci、WUE略高于海岛棉新海22号,但差异不显著(P<0.05),Tr低于新海22号。Gs反映外界环境CO2通过气孔进入到叶肉细胞叶绿体羧化部位的限制程度,是决定Pn高低的一个重要因素[20],新海22号Gs下降,导致Pn的降低,而Tr却高于陆地棉,可能与海岛棉叶片栅栏组织较薄以及叶片形态有关。正反交F1在Tr、Gs、Pn、WUE等光合参数表现出杂种优势,和Chen等研究认为杂种棉通过维持相对较高的光合能力来提高皮棉产量的研究结果[21]一致,正反交F2表现出明显的优势退化现象,但WUE显著增加,可能与陆海杂交后代叶型改变有关。
叶绿素荧光是光合作用的有效探针,通过对各种荧光参数的分析,可以得到有关光能利用途径的信息[22]。Fo和Fm分别是暗适下的最小荧光和最大荧光,它们是光系统Ⅱ(PSⅡ)反应中心处于完全开放和关闭时的荧光产量,Fo和Fm在陆海杂交F1代并未表现出明显优势,海岛棉做父本,杂交F2代的Fo和Fm值高于F1和亲本,ΦPSⅡ=Yield=(Fm′-Fs)/Fm′[23]任一光照状态下PSⅡ的实际量子产量,在相同环境条件下,海岛棉的实际量子产量显著高于陆地棉,正反交F1的实际量子产量介于两者之间,没有表现出杂种优势,正反交F2的实际量子产量也介于海岛棉和陆地棉之间。ETR代表了光电子传递的能力[24],杂种F1表现出了一些优势,但不明显,陆海杂交F2显著高于F1和亲本,反交F2电子传递速率显著降低,因此可以选用陆地棉做母本,比较容易选育高光效品种。qP=(Fm′-Fs)/Fv′=1-(Fs-Fo′)/(Fm′-Fo′)[25](基于“沼泽模型”)即由光合作用引起的荧光淬灭,反映了光合活性的高低,植物吸收的光能主要用于光化学反应、荧光、热耗散,本研究发现,海岛棉新海22号和陆地棉589光化学荧光淬灭系数qP差异不显著,正反交F1没有表现出杂种优势,陆海杂交F2的qP显著降低,但其电子传递能力显著高于亲本和F1,说明虽然光合电子传递能力较强,但其吸收的光能用于光化学反应的比例较低。NPQ=(Fm-Fm′)/Fm′=Fm/Fm′-1[25]反映了植物耗散过剩光能为热的能力,是衡量植物光保护能力的重要指标。589的NPQ值显著高于海岛棉,说明其可将过剩光能转化为热耗散,避免强光损伤,进行自我保护的能力较强,而陆海杂交后代则差异不显著,也没有杂种优势。Y(NO)[26]是指除了热扩散以外的荧光淬灭部分,若Y(NO)较高,表明入射光强超过了植物能接受的程度。这时,植物可能已经受到损伤,或者(尽管还未受到损伤)继续照光的话植物将要受到损伤,是光损伤的重要指标。陆地棉589的Y(NO)显著高于海岛棉,一方面说明其耗散的过剩光能较多,另一方面也说明其受到强光损伤的几率也在增大,陆海杂交后代的Y(NO)值介于陆地棉和海岛棉之间,且没有表现出杂种优势。Y(NPQ)[26]与光保护机制相关的所有热扩散,若Y(NPQ)较高,一方面表明植物接受的光强过剩,另一方面则说明植物仍可以通过调节(如将过剩光能耗散为热)来保护自身,是光保护的重要指标,陆地棉的Y(NPQ)值显著强于海岛棉,说明其进行光保护的能力高于海岛棉,当强光照射时,可将过剩光能转化为热扩散的能力高于海岛棉,从另外一个方面可说明陆地棉适应环境的能力较强,陆海杂种后代没有表现出杂种优势。Fv/Fm=(Fm-Fo)/Fm是PSⅡ[23]的最大量子效率,反映植物潜在最大光合能力,高等植物一般在 0.8~0.84 之间,当植物受到胁迫(stress)时,Fv/Fm显著下降,陆地棉Fv/Fm值显著低于海岛棉,说明海岛棉的最大光合能力显著高于陆地棉,陆海杂种后代的Fv/Fm介于陆地棉和海岛棉之间,也没有表现出明显的杂种优势。
参考文献:
[1]钱能. 陆地棉遗传多样性与育种目标性状基因(QTL)的关联分析[D]. 南京:南京农业大学,2009.
[2]张小全,王学德. 细胞质雄性不育陆地棉与海岛棉间杂种优势初步研究[J]. 棉花学报,2005,17(2):79-83.
[3]菲力申. 陆地棉与海岛棉杂种优势利用研究[D]. 南宁:广西大学,2008.
[4]常俊香. 棉花海陆杂交主要经济性状的遗传与杂种优势研究[D]. 郑州:河南农业大学,2007.
[5]石玉真,刘爱英,李俊文,等. 陆海种间杂交纤维品质性状的遗传及其F1群体优势分析[J]. 棉花学报,2008,20(1):56-61.
[6]李哲,杨金玉,崔秀珍,等. 棉花海陆杂交种高支纱纤维特征与应用研究[J]. 河南农业科学,2007(6):49-53.
[7]邓仲篪,孙济中,张金发,等. 杂交棉及其亲本光合特性的研究[J]. 华中农业大学学报,1995,14(5):429-434.
[8]姜磊,王旺华,李廷春,等. 棕色棉与白色棉杂交F1代吐絮期光合特性的杂种优势研究[J]. 棉花学报,2011,23(4):323-328.
[9]伍维模,董合林,危常洲,等. 南疆陆地棉与海岛棉光合-光响应及叶绿素荧光特性分析[J]. 西北农业学报,2006,15(4):141-146.
[10]高俊凤. 叶绿素含量定量测定[M]. 北京:高等教育出版社,2006:74-77.
[11]Jiang Y J,Zheng D M,Lu S Q,et al. Salt of soil affect quality of cotton[J]. Journal of Tarlm Agricultural Reclanation University,1994,8(2):23-27.
[12]张小全,王学德. 细胞质雄性不育陆地棉与海岛棉间杂种优势初步研究[J]. 棉花学报,2005,17(2):79-83.
[13]许大全. 光合速率影响因素[M]. 上海:上海科学技术出版社,2002:46-47.
[14]Kount R,Ilik P,Nans J,et al. On the limits of applicability of spectrophotometer and spectrofluorime tricmethods for the determination of chlorophyll a/b ratio[J]. Photosynthesis Research,1999,62(5):107-116.
[15]李丹丹,司龙亭,李季,等. 弱光下黄瓜幼苗叶片叶绿素含量的遗传分析[J]. 华北农学报,2009,24(1):133-137.
[16]尚莉,武继承,管秀娟,等. 河南主栽小麦品种叶绿素含量的变化特征[J]. 河南农业科学,2009(2):25-27.
[17]秦鸿德,张天真.棉花叶绿素含量和光合速率的QTL定位[J]. 棉花学报,2008,20(5):394-398.
[18]支金虎,伍维模,危常洲,等. 水分与氮素对膜下滴灌棉花叶片叶绿素含量时空分布的影响[J]. 西北农业学报,2007,16(1):7-12.
[19]张亚黎,姚贺盛,罗 毅,等. 海岛棉和陆地棉叶片光合能力的差异及限制因素[J]. 生态学报,2011,31(7):1803-1810.
[20]Farquhar G D,Sharkeyt D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Biology,1982,33:317-345.
[21]Chen Z H,Wu F B,Wang X D,et al. Heterosis in CMS hybrids of cotton for photosynthetic and chlorophyll fluorescence parameters[J]. Euphytica,2005,144(3):353-361.
[22]刘金山,王有武,高山,等. 陆海杂交棉叶绿素荧光参数及生理动态变化研究[J]. 种子,2012,31(6):4-9.
[23]Krause G H,Weis E. Chlorophyll fluorescence and photosynthesis:the basis[J]. Plant Physiol,Plant Mol,Biol,1991,42:313-349.
[24]吕晓菡,柴伟国.低温弱光下不同起源地辣椒幼苗光合特性的比较研究[J]. 浙江农业学报,2014,26(1):48-53.
[25]van Kooten O,Snel J F. The use of chlorophyll fluorescence nomenclature in plant stress physiology[J]. Photosynthesis Research,1990,25(3):147-150.
[26]Kramer D M,Johnson G,Kiirats O,et al. New fluorescence parameters for the determination of q(a) redox state and excitation energy fluxes[J]. Photosynthesis Research,2004,79(2):209-218.索欢,陈龙正,徐海,等. 小白菜根肿病接种鉴定研究[J]. 江苏农业科学,2016,44(5):193-195.
