贝莱斯芽孢杆菌EBV02 对棉花黄萎病的防治作用及机理
2022-12-22白红燕赵丽红蒲丹丹冯自力魏锋冯鸿杰顾爱星朱荷琴彭军张亚林
白红燕,赵丽红,蒲丹丹,冯自力,魏锋,冯鸿杰,顾爱星,朱荷琴,彭军,3*,张亚林*
(1.中国农业科学院棉花研究所/ 棉花生物学国家重点实验室,河南 安阳 455000;2. 新疆农业大学农学院/ 教育部棉花工程研究中心,乌鲁木齐 830052;3. 三亚中国农业科学院国家南繁研究院,海南 三亚 572024)
棉花(Gossypiumspp.)是重要的天然纤维作物,作为战略物资来源在国民经济中的地位举足轻重。 棉花黄萎病是由大丽轮枝菌(Verticillium dahliaeKleb.) 引起的一种土传真菌维管束病害,对棉花产量和纤维品质造成了巨大损失。 该病原菌能常年存活在土壤中,其寄主范围极广,能够危害多种农作物,且容易产生变异。 目前,对黄萎病采取了不同的防治措施, 如选择抗病品种、轮作倒茬和化学防治等,但防治效果不理想。 因此,亟需找到一种对环境友好且有效的防治方法,生物防治以绿色环保、发展潜力大等优势被人们关注并且应用于实践中。 目前,用于防治病害的拮抗微生物有细菌、真菌、放线菌等,其主要通过诱导植物产生系统抗性(induced systemic resistance,ISR),进一步争夺养分和定植空间,或通过产生植物激素和提供营养物质抑制病原侵染、促进植物生长[1]。生防细菌作为生防微生物重要的类群,在病害防治中发挥重要作用, 其中以芽孢杆菌(Bacillussp.)应用较多,如枯草芽孢杆菌(B.subtilis)、蜡状芽孢杆菌(B.cereus)、短小芽孢杆菌(B.pumilus)、 地衣芽孢杆菌 (B.licheniformis)等。贝莱斯芽孢杆菌(B.velezensis)作为芽孢杆菌属的一个新种,在自然界中广泛分布,对人畜无害,对环境无污染,并且能产生对多种病原微生物有拮抗作用的次生代谢物[2]。Martínez-Raudales等[3]发现,B.velezensis2A-2B 能够显著抑制辣椒疫霉(Phytophthora capsici)、腐皮镰刀菌(Fusarium solani)、尖孢镰刀菌(F.oxysporum)、立枯丝核菌 (Rhizoctonia solani) 的生长, 抑制率均在60%以上。此外,贝莱斯芽孢杆菌对一些植物病原细菌也具有较强的拮抗活性。 Cui 等[4]从马铃薯(Solanum tuberosum) 块茎中分离到的贝莱斯芽孢杆菌8-4, 对引起马铃薯疮痂病的链霉菌(Streptomyces galilaeus)具有较强的抑菌活性;田间试验结果显示,该菌不仅能够有效控制马铃薯疮痂病的发病率, 还能够显著提高马铃薯的产量。 可见,贝莱斯芽孢杆菌在农业病害防治方面有较好应用前景。
前期,本团队从健康棉花植株内筛选出一株对棉花黄萎病菌具有拮抗效果的内生细菌菌株EBV02。 本研究以该菌株为对象,经菌种鉴定为贝莱斯芽孢杆菌,进一步分析该菌株对棉花黄萎病菌菌丝生长、 产孢量和微菌核萌发的影响,以及对棉花的诱导抗性,以期探明该菌株防治棉花黄萎病的作用机理,从而为研发可用于田间防治棉花黄萎病的微生物菌剂奠定基础。
1 材料与方法
1.1 试验材料
供试内生细菌菌株:EBV02, 来自中国农业科学院棉花研究所棉花病害课题组建立的棉花生防菌种群资源库[5-6];黄萎病菌菌株为大丽轮枝菌强致病力菌株Vd080[7],棉花品种为耐黄萎病品种鲁棉研21 号[8],均由中国农业科学院棉花研究所棉花病害课题组提供。
培养基:马铃薯葡萄糖琼脂(potato dextrose agar,PDA)固体培养基、察氏(Czapek)培养基、卢里亚- 贝尔塔尼(Luria-Bertani, LB)固体培养基及液体培养基。
1.2 形态观察和分子生物学鉴定
形态观察和生理生化试验参照汪静杰等[9]的方法。 利用Bacterial DNA Kit 试剂盒(Omega 公司)提取EBV02 的基因组DNA,利用16S 核糖体DNA(ribosomal DNA, rDNA)的通用引物1492R 和27F,以50 μL 体系(DNA 1 μL)进行聚合酶链式反应(polymerase chain reaction,PCR)扩增,产物经琼脂糖凝胶电泳纯化回收后,送至上海派森诺生物科技有限公司测序,将测序结果与美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI)16S rDNA 数据库中的序列进行同源性比对,选出同源性最高且已公布的典型菌株序列, 采用Mega 7 的邻接法(neighbor-joining method)构建系统发育树,所选模型为Kimura 两参数模型, 自展法(Bootstrap)参数(抽样次数)设置为1 000。
1.3 EBV02 对大丽轮枝菌拮抗作用的检测
1.3.1对峙培养法测定EBV02 对V.dahliae 菌丝生长的影响。 EBV02 培养液的制备:将筛选到的贝莱斯芽孢杆菌EBV02 在LB 固体培养基上进行活化,选择单菌落接到LB 液体培养基中,置于37 ℃、150 r·min-1摇床培养24 h,获得贝莱斯芽孢杆菌培养液备用(600 nm 波长处的吸光值为0.8)。 在PDA 固体培养基中心接种已经活化的Vd080 菌饼(直径5 mm),然后在距离培养基中心20 mm 处对称放置2 个牛津杯,每杯内加入已经活化的50 μL 的EBV02 培养液, 以接种Vd080 不加EBV02 培养液作为空白对照, 每个处理重复5 次。 于25 ℃恒温培养箱培养,观察抑菌情况,在第10 天,利用十字交叉法测定Vd080菌落直径,计算抑制率(I)。 计算公式:I=[(D1-5)-(D2-5)]/(D1-5)×100%[10]。 式中,D1和D2分别为空白对照和处理的菌落直径。
1.3.2对扣培养法测定EBV02 对V.dahliae 菌丝生长的影响。 在PDA 固体培养基中心接种已经活化的Vd080 菌饼(直径5 mm),在LB 固体培养基上接种5 μL 已经活化好的EBV02 培养液,将上述2 个培养基对扣密封,以接种Vd080的PDA 培养基和空白LB 对扣培养为空白对照,每个处理重复5 次。 于25 ℃恒温培养箱培养10 d。利用十字交叉法测定Vd080 菌落直径, 采用1.3.1 中公式计算抑制率。
1.3.3混合培养法测定EBV02 培养液对V.dahliae 产孢量的影响。取EBV02 培养液1 mL 与等体积的Vd080 孢子悬浮液(含量为1×107mL-1)混合均匀,置于50 mL 离心管中,以无菌LB 液体培养基与Vd080 孢子悬浮液的等体积混合液为对照, 每个处理重复5 次, 在25 ℃摇床上培养, 分别在24 h、48 h 和96 h 时借助血球计数板计算孢子含量[11]。
1.3.4混合培养法测定EBV02 培养液对V.dahliae 微菌核萌发的影响。微菌核的培养及其悬浮液的制备参照Hu 等[12]的方法。 取100 μL 微菌核悬浮液与等体积的EBV02 培养液、1/2 稀释液和1/4 稀释液混合均匀, 置于18 ℃暗培养20 h后镜检,检测其萌发情况,计算萌发率,以微菌核悬浮液与无菌LB 液体培养基的等体积混合液为对照。 每个处理重复5 次, 每次观察100 个微菌核。
1.4 EBV02 对棉花生长的影响以及对黄萎病的防治效果
1.4.1EBV02 浸种对棉种萌发的影响。利用高速离心机收集菌体,菌体洗涤3 次,溶解在5%(质量分数,下同)羧甲基纤维素钠(carboxyl methyl cellulose sodium,CMC-Na) 溶液中制成原液,再制成1/2 稀释液和1/4 稀释液,备用。 将种子消毒后, 取50 粒棉种在原液、1/2 稀释液和1/4 稀释液中浸泡12 h,以5%CMC-Na 溶液浸种处理为对照,重复5 次,25 ℃培养,2 d 后检测萌发率和芽长[11]。
1.4.2温室试验测定EBV02 对棉花生长的影响以及对黄萎病的防治效果。 浸种法: 将蛭石、沙子、营养土按质量比3∶2∶1 混合均匀后装入纸钵(直径6 cm,高度10 cm),棉苗培育和Vd080的接种方法参照蛭石沙子无底纸钵定量蘸菌液法[13]。同时,用EBV02 培养液浸泡鲁棉研21 号棉种12 h,每个纸钵中播种8 粒棉种,每6 个纸钵为1 个处理,每个处理重复3 次,以无菌LB 液体培养基浸种作为对照。播种后7 d 进行间苗,每钵保留5 株棉苗。 待1 片真叶初现时,每个纸钵接种Vd080 孢子悬浮液(含量为1×107mL-1)10 mL,跟踪监测棉花长势情况,适时调查棉花发病情况。
灌根法:温汤(初始温度55 ℃)浸泡鲁棉研21 号棉种12 h,棉苗的播种、定苗参照浸种法。待棉花1 片真叶初现后,用EBV02 培养液灌根,处理后3 d,每个纸钵接种Vd080 孢子悬浮液(含量为1×107mL-1)10 mL,每6 个纸钵为1 个处理,以等体积无菌LB 液体培养基灌根为对照。
接种Vd080 孢子悬浮液后15 d 参照5 级分级标准调查发病情况,同时计算病情指数和防治效果[14-15]。 播种后60 d,每个处理随机选取15 株棉花测量株高、主根长、鲜物质质量等生物量指标。棉花黄萎病病情指数计算公式为DI=Σ(Ni×i)/(N×4)×100。式中,DI为病情指数,Ni为各级病株数,i为病级数值,N为调查总株数。 防治效果(E)计算公式为E(%)=(DI0-DI1)/DI0×100。式中,DI0为对照病情指数,DI1为处理病情指数。
1.4.3大田试验测定EBV02 对棉花生长的影响以及对黄萎病的防治效果。 在棉花黄萎病发生严重的病圃中种植耐病品种鲁棉研21 号。 根据试验设计种植小区,每个小区长3.3 m,宽2.8 m,株距20 cm。 试验设置3 次重复,3 行为1 个处理,即9 行为1 个处理区组。
培养液浸种:取适量消毒后的种子在EBV02培养液中浸种处理12 h,无菌水多次洗涤种子后播种,以无菌LB 液体培养基浸种为对照,播种后10 d 统计鲁棉研21 号种子的出苗数并计算出苗率;出苗后25 d,每个处理随机取15 株棉苗,测量主根长、株高、鲜物质质量等指标。 培养液灌根:待棉花出苗后,在棉花根部加入EBV02 培养液,每小区用量0.8 L,以等体积无菌LB 液体培养基灌根为对照。 培养液喷雾: 将制备好的EBV02 培养液喷施在棉苗上, 之后每隔20 d 喷雾1 次,共2 次,每小区用量0.4 L,以等体积无菌LB 液体培养基喷雾为对照。 病情的分级标准、病情指数和防治效果的计算同1.4.2。
棉花收获时调查其株高、单株果枝数、单株结铃数、30 铃籽棉质量和30 铃皮棉质量。
1.5 EBV02 防治棉花黄萎病作用机理探究
1.5.1EBV02 培养液诱导棉花叶片抗性的检测。将消毒后的种子浸泡在已活化好的EBV02 培养液中12 h,将种子用无菌水冲洗2~3 次,以每钵8 粒播种在纸钵中, 棉苗长到3 片真叶后,用EBV02 培养液进行灌根处理,2 d 后取棉花真叶进行表面消毒,再用无菌水冲洗2~3 次,置于水琼脂培养基表面,将叶片表面造成伤口,在叶片伤口处接种Vd080 菌饼。 以单独接种无菌LB 液体培养基作为对照,每个处理重复5 次。 25 ℃培养7 d,观察叶片损伤情况[11]。
1.5.2EBV02 培养液诱导棉花叶片活性氧爆发的检测。 棉苗长出2 片真叶后,用EBV02 培养液灌根,每个纸钵10 mL,以接种等量无菌LB 液体培养基为对照。 接种2 d 后,利用3,3- 二氨基联苯胺(diaminobenzidine,DAB)组织染色法检测棉花叶片中活性氧爆发和积累,取长势相近的棉花真叶,用无菌水洗涤后置于50 mL 离心管中,取适量DAB 染液(1 g·L-1,pH 7.5)加入离心管中,室温避光染色8 h;去除染液后,加入适量95%(体积分数)乙醇在沸水中水浴2 min 脱去叶绿素, 去除液体后加适量无水乙醇继续脱色,直至绿色完全脱去。 最后将叶片浸泡到70%(体积分数)甘油中,赶出叶片细胞间气泡,将叶片放置到载玻片上采用体式显微镜Leica M165FC 进行观察。
1.5.3棉花叶片胼胝质沉积测定。 棉苗长出2 片真叶后, 用EBV02 培养液灌根, 在棉花接菌前(对照)以及接菌后48 h 分别随机取棉苗5 株。取1 片完整真叶,在乙醇与乙酸体积比为3∶1 的固定液中固定2~3 h,脱去叶绿素。 依次在体积分数为70%和50%的乙醇中浸泡3 h, 然后于无菌水中浸泡过夜。 次日倒掉水后,轻轻漂洗叶片,将叶片在10%(质量分数)NaOH 中浸泡1~2 h,使叶片变透明。 用蒸馏水清洗叶片4 次, 随后于0.01%(质量分数)的苯胺蓝染色液中避光培养3 h, 最后用荧光体式显微镜Nikon 80i 观察胼胝质的积累情况。
1.5.4棉花防御相关基因表达量的测定。 棉花1片真叶初现后,用EBV02 培养液灌根,处理后3 d,每纸钵接种10 mL 的Vd080 孢子悬浮液(含量为1×107mL-1), 每6 个纸钵为1 个处理,每个处理重复3 次,以等体积无菌LB 液体培养基灌根为对照。 接种Vd080 后24 h、48 h 和72 h 取样。 采用RNAprep Pure Plant Kit(天根公司)提取棉花叶片RNA,用NanoDrop 2000 检测RNA 的质量浓度,并将RNA 的质量浓度调到100 mg·L-1,备用。cDNA 第1 链的合成和荧光定量PCR 参照张芸等[16]的方法,检测棉花叶片中防御相关基因过氧化物酶基因(peroxidase gene,POD)、多酚氧化酶基因(polyphenol oxidase gene,PPO)、苯丙氨酸氨裂合酶基因 (phenylalanine ammonia-lyase,PAL)、病程相关蛋白10 基因(pathogenesis-related protein 10 gene,PR10)和茉莉酸信号转导途径基因JAR1(jasmonate resistant gene 1)的表达量,以棉花中高度保守的基因ubiquitin为内参基因,相关引物见表1。
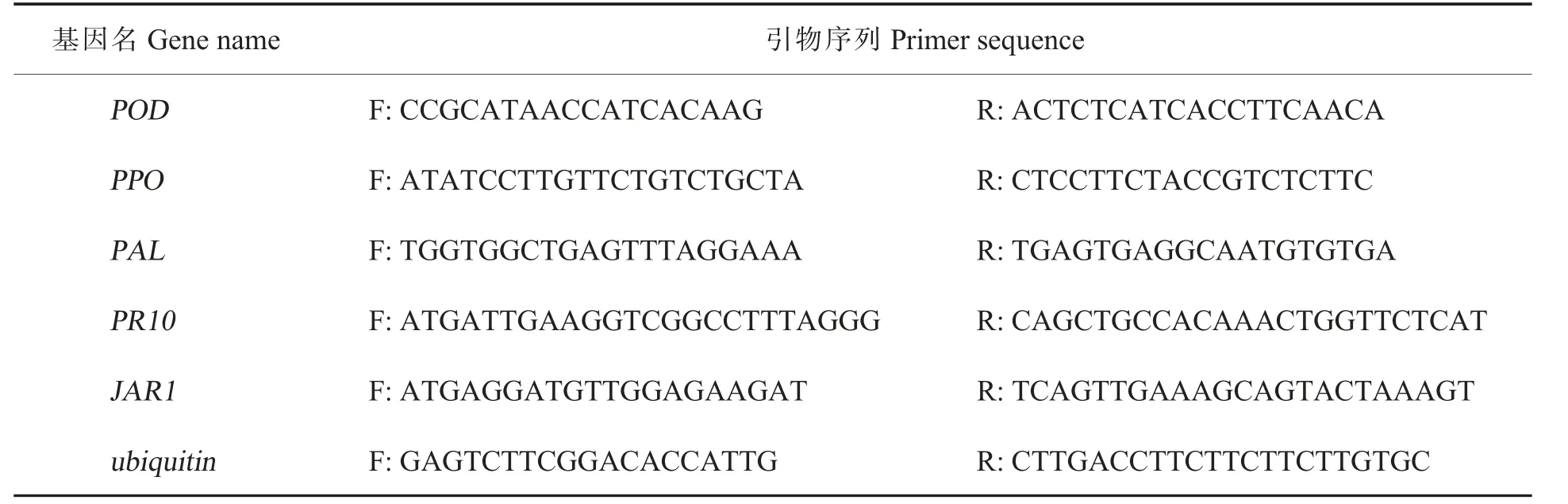
表1 防御相关基因的特异性引物Table 1 Specific primer sequences of defense-related genes
1.5.5数据处理。 试验数据利用Microsoft Excel 2019 进行处理,采用IBM SPSS Statistics 26.0 统计软件中的单因素方差分析程序进行差异显著性分析。 采用邓肯新复极差法比较处理之间的差异显著性(P<0.05)。 所有数据在统计分析时,先进行了正态性检验,均符合正态分布。
2 结果与分析
2.1 EBV02 的菌种鉴定
EBV02 在LB 培养基上生长良好,经形态学鉴定,菌落颜色为乳白色,表面光滑,不透明,单菌落形态为球形(图1A);经革兰氏染色,菌体呈紫红色的短杆状(图1B),确定为革兰氏阳性细菌。 根据测序结果与GenBank 中相关菌株16S rDNA 序列构建系统发育树, 其与模式菌株B.velezensis的相似性最高, 为99.72%(图1C)。伏- 波试验(Voges-Proskauer test,VP test)和硝酸盐还原反应均呈阳性,可以水解淀粉,不能产生H2S。 结合其培养特征、生理生化反应和分子生物学鉴定, 确定该内生细菌为贝莱斯芽孢杆菌,记为EBV02。
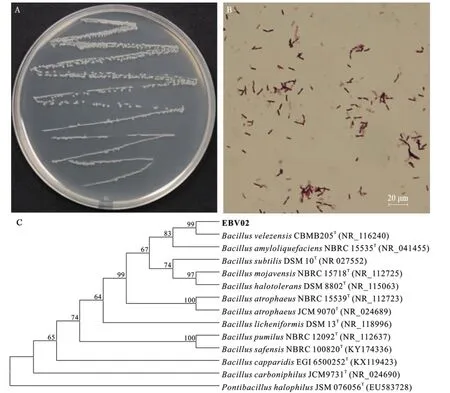
图1 EBV02 的菌种鉴定Fig. 1 Species identification of the strain EBV02
2.2 内生细菌对大丽轮枝菌的拮抗作用分析
2.2.1EBV02 对大丽轮枝菌菌丝生长的抑制作用。对峙培养试验结果(图2A、B)显示,培养10 d后,经EBV02 处理的Vd080 菌落直径为19.4 mm,未经EBV02 处理的Vd080 菌落直径为44.2 mm,EBV02 对Vd080 菌丝生长的抑制率为63.27%。表明,EBV02 的非挥发性代谢产物对Vd080 的菌丝生长有较强的抑制作用。
对扣培养试验结果(图2C、D)显示:对照中Vd080 菌落正常生长的直径为51.3 mm, 菌落为白色、无微菌核;而接种EBV02 的Vd080 菌落上长出了微菌核, 菌丝生长被抑制, 菌落直径为23.6 mm,抑制率为59.83%。表明,EBV02 的挥发性代谢产物对Vd080 的菌丝生长也有抑制作用。
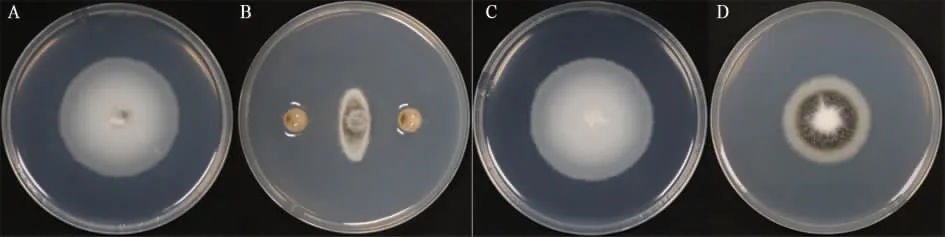
图2 培养10 d 后EBV02 对大丽轮枝菌菌丝的抑制作用Fig. 2 Inhibitory effect of the strain EBV02 on the hyphae of V. dahliae after 10 d of culture
2.2.2EBV02 对大丽轮枝菌产孢的抑制作用。产孢量试验结果(图3)显示:在混合培养24 h、48 h和96 h 时,EBV02 对Vd080 的产孢量均有显著抑制作用, 抑制率分别为32.24%、41.61%和21.86%; 不同培养时间的EBV02 培养液对Vd080 产孢量的抑制作用有差异, 其中混合培养48 h 时的抑制效果最好。 上述结果表明EBV02 培养液对大丽轮枝菌分生孢子的产生具有抑制作用。

图3 大丽轮枝菌产孢量的比较Fig. 3 Comparisons of spore production of Verticillium dahliae
2.2.3EBV02 对大丽轮枝菌微菌核萌发的抑制效果。 微菌核与EBV02 培养液暗培养20 h 的萌发结果(图4)显示:EBV02 培养液原液、1/2 稀释液和1/4 稀释液中混合培养的微菌核萌发率均与对照(41.20%)差异显著,抑制率分别为64.56%、41.75%和31.55%,表明EBV02 培养液对微菌核萌发有抑制作用,且抑制作用随着EBV02 培养液浓度的增大而增强。
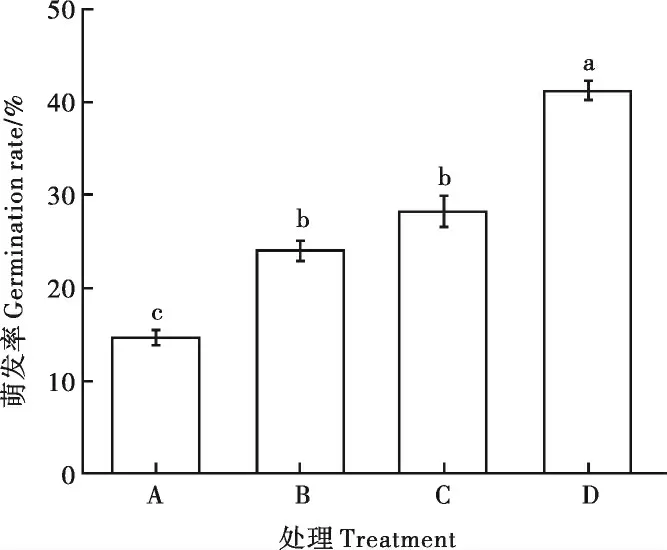
图4 暗培养20 h 微菌核萌发率比较Fig. 4 Comparisons of sclerotium germination rates at 20 h of dark culture
2.3 EBV02 对棉花黄萎病的防治效果
2.3.1EBV02 浸种处理对种子萌发的影响。如表2 所示,经过不同浓度菌液浸泡的处理组,棉种萌发率和芽长分别比对照组提高了3.26%和8.50%、3.26%和6.54%、4.88%和16.34%,但差异均不显著。

表2 浸种处理棉种萌发率和芽长比较Table 2 Comparisons of germination rates and shoot length between seed soaking treatments
2.3.2EBV02 对棉花黄萎病的温室防治效果。结果(表3)显示:EBV02 浸种处理的发病率和病情指数均显著低于对照,对棉花黄萎病的温室防治效果为68.33%;EBV02 灌根处理的棉苗其发病率和病情指数与对照组相比,分别显著降低了24.97%和57.62%。 表明,EBV02 浸种和灌根处理都可以显著降低棉苗的发病率和病情指数,对棉花黄萎病具有良好的防治效果。 此外,与灌根法相比,浸种法对棉花黄萎病的防治效果更好(图5)。

图5 温室条件下EBV02 处理对棉花黄萎病的防治效果Fig. 5 Control effect of EBV02 treatment on cotton Verticillium wilt in a greenhouse
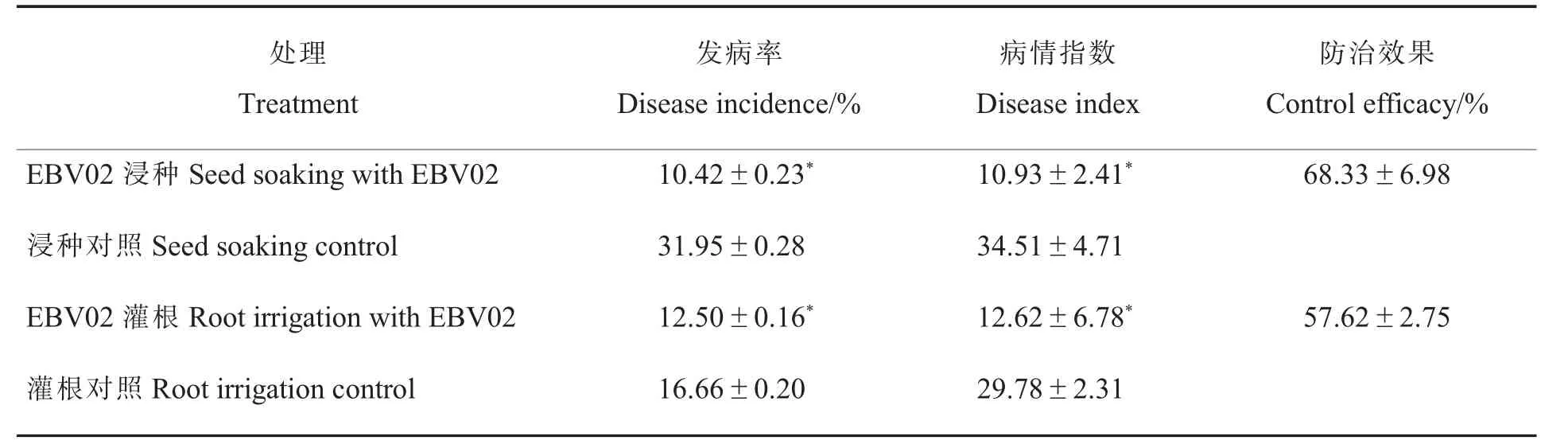
表3 接种大丽轮枝菌15 d 后EBV02 对棉花黄萎病的温室防治效果Table 3 Control effect of EBV02 on cotton Verticillium wilt at 15 d after inoculation with V. dahliae in a greenhouse
2.3.3温室环境中不同处理下的棉花生长指标分析。 如表4 所示,培养液浸种和灌根处理生长25 d 的棉苗主根长、株高、地上部鲜物质质量和总鲜物质质量均显著高于对照, 增幅分别为62.26%和28.57%、77.77%和34.00%、108.00%和65.43%、103.61%和65.91%,表明EBV02 对棉苗的生长发育有促进作用。

表4 出苗后25 d 不同处理棉花生长指标的比较Table 4 Comparison of cotton growth indexes of different treatments at 25 d after seedling emergence
2.3.4EBV02 对棉花黄萎病的大田防治效果。棉花播种后60 d 的大田棉苗发病情况调查结果(表5)显示,培养液浸种、灌根和喷雾处理的棉苗发病率和病情指数与对照相比均显著降低,防治效果为23.82%~37.25%。 其中,培养液喷雾防治棉花黄萎病效果最好,发病率和病情指数与对照相比分别降低了41.09%和37.25%。
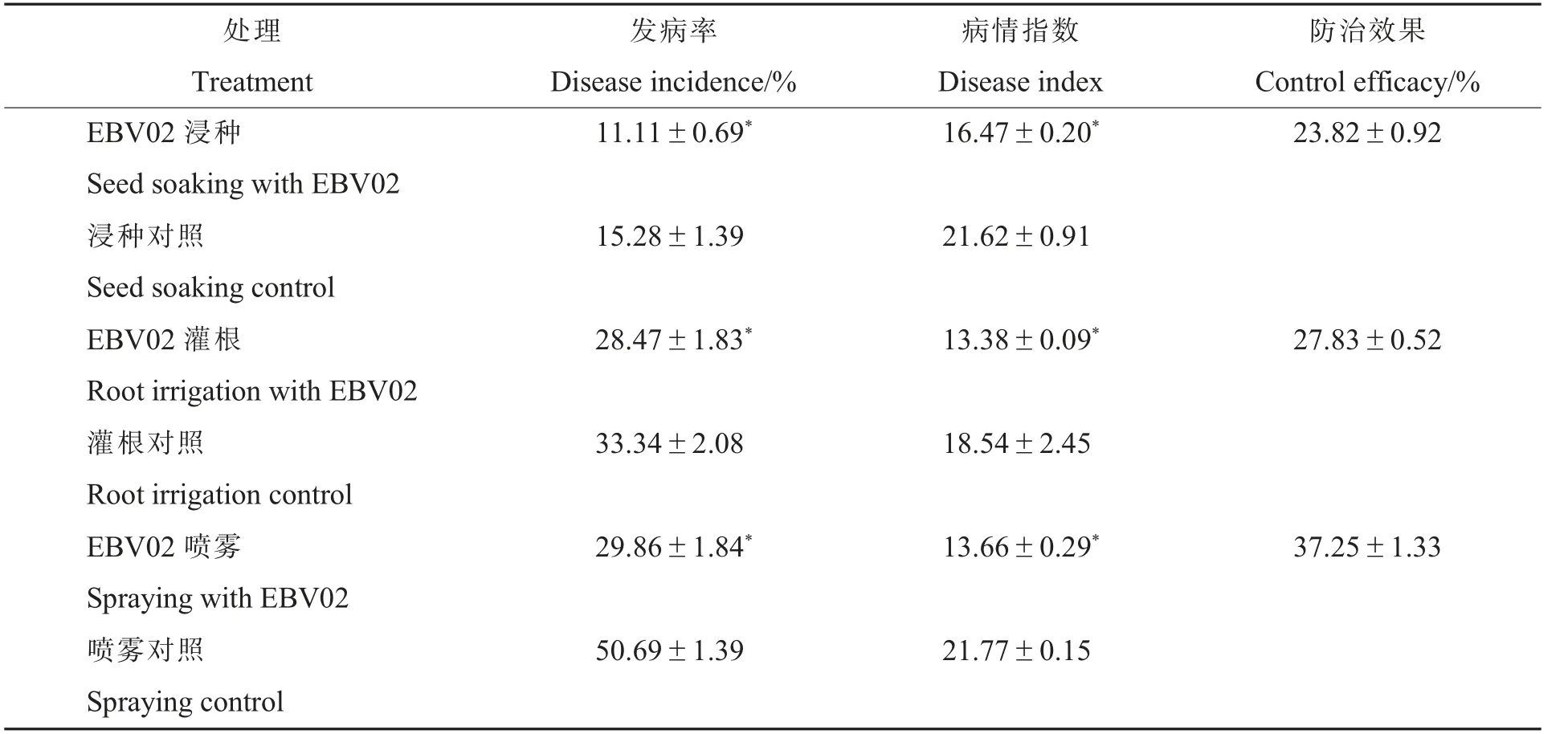
表5 棉花播种后60 d EBV02 对棉花黄萎病的大田防治效果Table 5 Field control effect of the strain EBV02 on cotton Verticillium wilt at 60 d after sowing
2.3.5浸种处理的大田棉花出苗率及生长指标分析。由表6 可知,与对照相比,EBV02 浸种处理的棉花出苗率显著提高,增幅为6.40%,棉苗的地上部鲜物质质量和总鲜物质质量分别显著增加8.41%和7.67%,表明EBV02 浸种处理有利于棉苗的物质积累。此外,EBV02 处理的棉苗主根长、株高与对照均无显著差异。
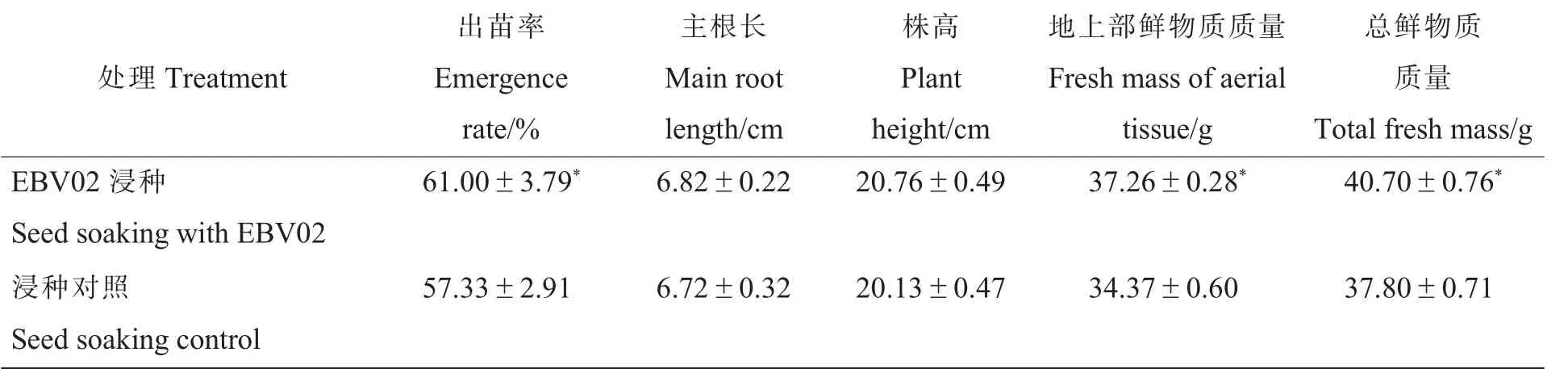
表6 浸种处理棉花出苗和生长指标的比较Table 6 Comparisons of cotton emergence rates and growth indexes between seed soaking treatments
2.3.6EBV02 对大田棉花产量相关性状的影响。如表7 所示,EBV02 培养液浸种、 灌根和喷雾处理均能显著增加收获期棉花的株高、30 铃籽棉质量和30 铃皮棉质量。 其中,浸种、灌根和喷雾处理的30 铃籽棉质量和30 铃皮棉质量与对照相比分别增加12.60 g 和5.00 g、5.53 g 和3.47 g、11.53 g 和4.47 g, 增幅分别为8.34%和8.26%、3.38%和5.60%、7.04%和7.06%。
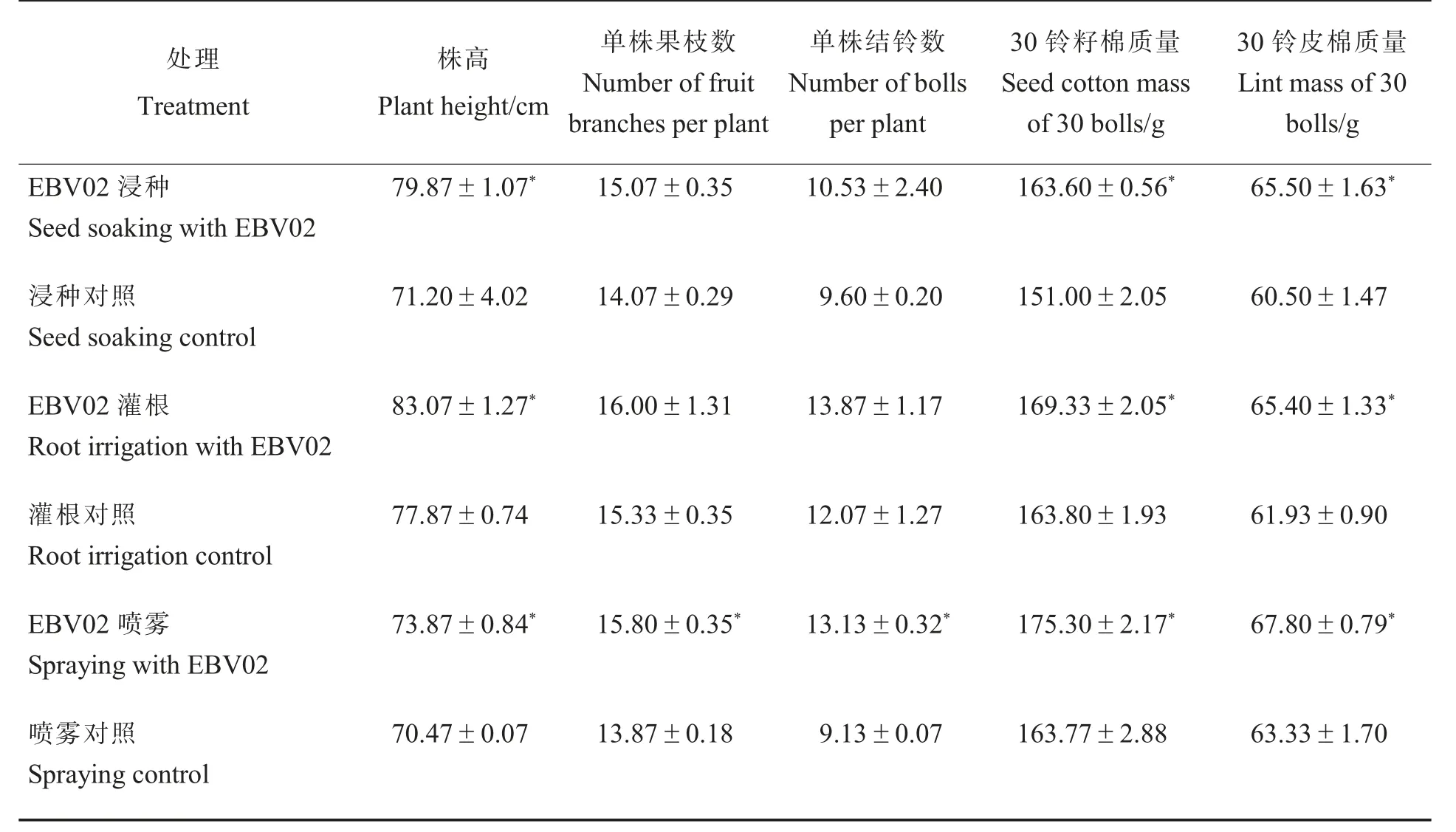
表7 大田条件下不同EBV02 处理与对照棉花产量及产量相关性状的比较Table 7 Comparisons of cotton yield and yield-related traits between EBV02 treatment and control under field conditions
2.4 EBV02 防治棉花黄萎病的作用机理探究
2.4.1EBV02 培养液诱导棉花抵御大丽轮枝菌。将棉花真叶在水琼脂培养基表面培养7 d 后,EBV02 培养液灌根处理的叶片表面菌丝附着较少, 而灌根对照的叶片表面有大量菌丝定植,坏死面积较大,且叶片损伤程度更严重(图6)。

图6 EBV02 诱导棉花叶片抗大丽轮枝菌表现Fig. 6 Performance of EBV02 enhanced cotton leaves against V. dahliae
2.4.2EBV02 培养液诱导棉花叶片活性氧爆发。镜检结果(图7)表明,经EBV02 培养液灌根处理的棉花叶片中褐色沉淀较多,而在对照中,褐色沉淀较少, 表明EBV02 诱导了棉花叶片中活性氧爆发。
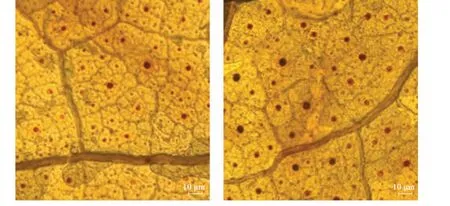
图7 EBV02 引起棉花叶片活性氧爆发表型Fig. 7 Performance of EBV02 triggered the burst of reactive oxygen species in cotton
2.4.3棉花叶片中胼胝质沉积对比。 镜检结果(图8) 表明, 接种EBV02 培养液2 d 后, 经EBV02 灌根处理的棉花叶片中胼胝质的积累量较多, 而在对照中胼胝质的积累量较少, 表明EBV02 诱导了棉花叶片中胼胝质的积累。

图8 EBV02 诱导棉花叶片胼胝质的积累Fig. 8 EBV02 induced callose accumulation in cotton leaves
2.4.4EBV02 诱导棉花防御相关基因的表达分析。 基因表达量检测结果(图9)表明,经过EBV02 培养液灌根处理后, 叶片中POD、PPO、PAL、PR10和JAR1基因的表达量均升高。PAL和PR10在接种后24 h 的表达量分别是对照组的1.80 倍和1.32 倍, 在接种后72 h 分别是对照组的21.35 倍和108.00 倍。 表明EBV02 灌根处理能激活棉花的防御系统,增强其对棉花黄萎病的抵抗能力。

图9 防御相关基因表达量检测结果Fig. 9 The expression levels of defense-related genes
3 讨论
利用拮抗微生物及其代谢产物来抑制植物病原菌的发生和危害, 是绿色环保的防治策略。目前,研究发现贝莱斯芽孢杆菌对多种植物病原菌具有抑制作用, 例如贝莱斯芽孢杆菌DPT-03对花生白绢病的防治效果达到了62.50%,可以有效抑制病原菌菌丝生长和微菌核的形成,抑制率高达87.20%~88.51%, 显著降低花生白绢病的发病率和发病程度,且提高花生产量[17]。本研究从棉花的内生细菌EBV02 入手,经形态特征、生理生化特征和分子生物学鉴定, 明确EBV02 为贝莱斯芽孢杆菌。 通过对峙培养和对扣培养试验,发现EBV02 对大丽轮枝菌Vd080 菌丝生长的
抑制率分别为63.27%和59.83%;EBV02 培养液对Vd080 产孢量和微菌核萌发的平均抑制率分别为31.90%和45.95%; 对棉花黄萎病菌温室防治效果高达68.33%,并能激活棉花抗病防御系统。
目前,对内生菌生防潜力的测定绝大多数在实验室或温室进行,在田间进行的较少。 室内筛选出对病原菌拮抗效果较强的生防菌株,在大田中常出现防治效果不稳定的情况[18]。温室试验中,采用EBV02 培养液浸种和灌根处理对棉花苗期黄萎病均表现出较好的防治效果(57.62%~68.33%), 但是大田防治效果仅为23.82%~37.25%,分析其原因可能是内生菌是一个生物活体,大田环境中的一些生物和非生物因素都会影响其防治效果[19]。董玉洁[20]用生防菌链霉菌GX-8处理苦瓜幼苗,与对照相比,株高增加60.00%,茎粗增加12.00%,鲜物质质量增加54.05%,干物质质量增加20.59%。有研究已证实生防菌可以提高棉花产量、株高、单株结铃数、单株果枝数等[16]。温室试验中经EBV02 培养液处理的棉花,其主根长、株高、地上部鲜物质质量和总鲜物质质量均显著高于对照组, 对棉苗生长具有较好的促生作用。 大田试验中EBV02 培养液浸种、灌根和喷雾处理对棉花的籽棉和皮棉增产效果显著。 这些结果表明,贝莱斯芽孢杆菌EBV02 不仅能够防控棉花黄萎病, 还能够明显提高棉花产量。
生防菌的诱导抗性作用须与竞争作用、重寄生作用、拮抗作用以及植物促生作用以及其他作用机制共同协作才能发挥增效作用[21]。 活性氧在调节植物生长和增强抗逆性方面具有重要作用[22],胼胝质被视为植物抵抗病原菌的物理屏障[23]。 EBV02 培养液灌根处理,可显著抑制大丽轮枝菌对棉花叶片的侵染,促进棉花叶片活性氧爆发和胼胝质积累,减少大丽轮枝菌在叶片中的定植。 苯丙氨酸解氨酶是合成木质素、酚类物质等抑菌物质的关键酶,对植物抗病系统的激活至关重要[24]。PR10 是植物在受到各种生物和非生物胁迫后产生的一类病程相关蛋白,在植物的发育和对外界逆境环境的应激反应中发挥重要作用[25]。 本研究发现,EBV02 培养液灌根处理可诱导PAL、POD、PPO、PR10和JAR1的显著上调表达。 综上,贝莱斯芽孢杆菌EBV02 能够诱导棉花对大丽轮枝菌产生系统抗性,具有较好的防病效果。 但本研究对其防病机理只是初步探讨,对其更深层次的作用机理包括其中的信号转导途径还有待进一步研究。
4 结论
筛选并鉴定了内生细菌贝莱斯芽孢杆菌EBV02, 其非挥发性代谢产物能有效抑制棉花黄萎病菌Vd080 的菌丝生长、分生孢子的产生和微菌核的萌发, 挥发性代谢产物也可以抑制Vd080 菌丝的生长;温室和大田试验表明,施用EBV02 能降低黄萎病发病率和病情指数, 且对棉苗生长发育及产量具有促进作用。 此外,EBV02 可提高棉花叶片抵抗黄萎病菌侵染的能力,引起叶片活性氧爆发、 胼胝质积累和防御基因的上调表达,表明EBV02 能诱导棉花对黄萎病的系统抗性。 综上所述,EBV02 在防治棉花黄萎病方面具有较好的应用前景,可为研发微生物菌剂提供参考。
