陆地棉-异常棉异附加系抗旱耐盐性评价及关键生理生化指标测定
2022-12-22上官艺馨曹静季为陈祥龙徐鹏郭琪沈新莲徐珍珍
上官艺馨,曹静,季为,陈祥龙,徐鹏,郭琪,沈新莲,徐珍珍
(江苏省农业科学院经济作物研究所/ 农业农村部长江下游棉花与油菜重点实验室,南京 210014)
棉花是世界上最重要的经济作物之一,是纺织工业原料、食用油和饲料蛋白的重要来源。 我国既是棉花生产大国,也是原棉消费大国,还是棉纱、棉纺织品及服装出口大国。2001—2019 年,我国棉花年均消费量在800 万t 左右, 而年均产量约600 万t,缺口量大[1]。 随着粮棉争地矛盾加剧,我国棉花生产区域化更加明显,由原来的以长江流域、黄河流域、西北内陆三大棉区为主逐步向西北内陆和滨海盐碱地转移。2021 年新疆棉花种植面积和总产均达到全国的80%以上[2]。 但新疆地区气候干燥、蒸发强烈、年降水量少且时空分布不均,干旱问题十分突出,制约了新疆棉花生产的可持续发展[3]。另外,新疆地区受不同程度盐渍化危害的耕地面积占耕地总面积的30.12%,占低产田面积的63.2%,其中受到中度、重度盐渍化威胁的低产田面积占耕地总面积的22.06%[4]。因此,最大限度地节省水资源并发展盐碱地植棉是新疆乃至我国棉花生产的发展方向。
现有的陆地棉(Gossypium hirsutumL.)品种均是在正常环境条件下选育, 抗旱耐盐性有限,抗逆种质资源缺乏。刘国强等[5]鉴定了3 000 多份陆地棉材料的耐盐性,发现只有3 份材料在0.4%NaCl 条件下表现为耐盐;Ashraf 等[6]考察了6 个不同来源陆地棉杂交种的耐盐性,5 000 株F2幼苗中,仅有20 株具有较强的耐盐性;杨淑萍等[7]鉴定了新疆30 个棉花品种萌发期和苗期的耐盐性,发现仅有8 个品种的耐盐级别为2 级,无高耐盐材料;王宁等[8]研究了甘肃河西走廊棉区19个主栽棉花品种(系),表现稳定的强耐盐品种仅有3 个,其他材料的耐盐性中等或较弱。
棉花野生种(近缘种)大多生长于半干旱、亚热带的沿海和岛屿,不但对干旱环境具有较强的适应性,而且能忍受潮汐的侵袭。 如加拉帕戈斯群岛的旱地棉(G.aridum)利用发达的根系、较小的叶片和较高的叶片蜡质含量提高对土壤水分的吸收能力并减少蒸腾失水;生长于墨西哥西南部的棉花野生种可通过脱落叶片应对干旱胁迫[9];早期驯化的陆地棉野生种系尖斑棉(G.punctatum)通过降低角质层蒸腾作用来提高水分利用率[10]。长距离漂移在棉属种进化过程中发挥着重要作用,野生种种子对海水的耐受性使得长距离漂移成为可能[9]。 加拉帕戈斯群岛上的野生种达尔文氏棉(G.darwinii) 和克劳茨基棉 (G.klotzschianum)可以生长在潮汐带标志3 m 之内;戴维逊氏棉(G.davidsonii)种子在海水中浸泡8 周后,依然有100%的发芽率。 在长期的进化过程中,一些棉属种形成了沿海物种独特的结构与生理特点,如辣根棉(G.armourianum)和哈克尼西棉(G.harknessii)叶片的皮质结构可以抵抗海水的浸泡[11]。棉属野生种在进化过程中形成了多种形态结构、生理特征及分子机制等抵御恶劣的外部环境,可能存在可用于改良陆地棉抗旱耐盐性的优异基因或位点。 因此,通过种间杂交将野生种的优良基因渐渗到陆地棉中是棉花遗传改良的有效方法。
棉属二倍体野生种异常棉(G.anomalum)生长于非洲的西南部和撒哈拉沙漠的边缘[12]。 与陆地棉栽培种相比,异常棉具有植株较矮且密被茸毛、 叶色浅等与抗旱或耐盐相关的性状特点,可能具有可用于改良陆地棉抗旱耐盐性的遗传多样性。 由于染色体倍性水平和遗传结构的差异,二倍体野生棉和陆地棉之间存在严重的种间重组抑制以及低重组率导致的连锁累赘等问题,导致异常棉在陆地棉遗传改良中的利用十分有限,大量优异基因尚未被挖掘。 前期研究中,我们以陆地棉86-1 为母本,异常棉为父本,杂交获得了三倍体F1,并通过秋水仙素加倍获得了六倍体F1杂种[13]。 以六倍体F1为母本,陆地棉苏棉8289(Su8289)为轮回亲本,连续回交4 次、自交3 次,每一世代利用覆盖异常棉基因组的特异的简单重复序列(simple sequence repeat,SSR)标记[14]进行辅助选择, 获得了一套完整的陆地棉-异常棉异附加系材料[15]。 本研究通过对陆地棉- 异常棉异附加系相关生理生化指标的测定,进行苗期抗旱和耐盐性鉴定,筛选出具有抗旱或耐盐潜力的异附加系,旨在为棉花抗旱耐盐育种提供理论与材料基础。
1 材料与方法
1.1 植物材料
供试材料包括轮回亲本陆地棉苏棉8289 和一套由本实验室保存的完整的陆地棉- 异常棉单体异附加系 (monosomic alien addition line,MAAL)[15]。 异常棉基因组属于B1组,陆地棉-异常棉异附加系的命名以MAAL 开头, 按照异常棉13 条染色体的顺序编号,B 代表附加的外源异 常 棉 染 色 体, 即MAAL_1B、MAAL_2B、MAAL_3B、MAAL_4B、MAAL_5B、MAAL_6B、MAAL_7B、MAAL_8B、MAAL_9B、MAAL_10B、MAAL_11B、MAAL_12B 和MAAL_13B。
1.2 基因型鉴定
Su8289 和陆地棉- 异常棉异附加系的棉籽经工业硫酸脱绒后,在晒场上自然晾晒风干。 种子完全干燥后, 每份材料选择约200 粒种子在28 ℃恒温培养箱中催芽。 基质和蛭石按5∶1 的体积比充分混匀,等量称取60 g 分装于一次性纸杯中(直径7 cm,高8 cm)。 将发芽的种子播于纸杯中,每纸杯1 粒,浇水70 mL;置于光照/ 黑暗时间为16 h (28 ℃)/8 h (25 ℃), 相对湿度为60%培养箱中生长。待棉苗生长至一叶一心期,掐取少量叶片,采用改良的CTAB 法提取DNA[16]。分别利用异常棉1~13 号染色体上的20、16、13、12、24、18、19、18、15、16、27、15、17 对 特 异SSR引物(附表1)[14,17]进行聚合酶链式反应(polymerase chain reaction, PCR)以鉴定陆地棉- 异常棉异附加系MAAL_1B~MAAL_13B。 PCR反应体系为10.0 μL, 包括DNA 模板2.0 μL、10×缓冲液(Mg2+)1.0 μL、dNTP 0.2 μL、Taq DNA 聚合酶(宝日医生物技术有限公司,北京)0.1 μL、正反向引物各0.5 μL 和dd H2O 5.7 μL。PCR 程序共六步,第一步:95 ℃预变性5 min,第二步:94 ℃变性30 s,第三步:55 ℃退火45 s,第四步:72 ℃延伸1 min, 第二步到第四步重复30 个循环,第五步:72 ℃延伸10 min,第六步:10 ℃保存。 将PCR 产物在8%(质量分数)非变性聚丙烯酰胺凝胶上进行电泳检测,每个样品上样1.0 μL,180 V 恒压电泳1.5 h,电泳结束后进行银染[18]。
1.3 苗期抗旱和耐盐性鉴定
抗旱性评价方法:经PCR 筛选到的陆地棉-异常棉异附加系和Su8289 生长至两叶一心时,每纸杯浇水70 mL,干旱15 d 后复水,观察幼苗表型并在复水后1 d 统计存活率。 3 次生物学重复, 每个生物学重复选取10 株长势一致的幼苗进行测定。
耐盐性评价方法:经PCR 筛选到的陆地棉-异常棉异附加系和Su8289 生长至两叶一心时,每纸杯浇350 mmol·L-1的NaCl 溶液70 mL,以每纸杯浇70 mL 清水为对照处理。处理后7 d,测定清水对照和NaCl 处理植株的株高、 地上部鲜物质质量和地上部干物质质量; 统计Su8289 和异附加系的相对株高、相对地上部鲜物质质量和相对地上部干物质质量。 各设3 次生物学重复,每个生物学重复挑选5 株长势一致的幼苗进行测定。
株高(cm):从子叶节到主茎顶端的距离。
地上部鲜物质质量(g):植株子叶节以上部分的鲜物质质量。
地上部干物质质量(g):取植株子叶节以上部分,于100 ℃烘箱中烘干至质量恒定后称量。
按照以下公式分别计算相对株高、相对地上部鲜物质质量和相对地上部干物质质量:

1.4 生理生化指标测定
于干旱处理当天和处理后10 d、盐处理当天和处理后3 d 选取具有抗旱和耐盐潜力的陆地棉-异常棉异附加系和Su8289 的倒二叶进行相关生理生化指标的测定。 过氧化氢 (H2O2)、 丙二醛(malondialdehyde,MDA)和脯氨酸(proline,PRO)的含量以及抗氧化系统中过氧化物酶(peroxidase, POD)、超氧化物歧化酶(superoxide dismutase, SOD)和过氧化氢酶(catalase, CAT)的活性均采用试剂盒测定, 具体操作参照试剂盒说明书。 生理生化试剂盒均购自于南京建成生物工程研究所,包括过氧化氢测定试剂盒(比色法)、丙二醛(MDA)测定试剂盒(硫酸巴比妥酸法)、脯氨酸(Pro)测定试剂盒(比色法)、过氧化物酶(POD)测定试剂盒(测植物)(比色法)、总超氧化物歧化酶(T-SOD)测试盒(羟胺法)和过氧化氢酶(CAT)测定试剂盒(可见光法)(钼酸铵法)。 以上所有生理生化指标测定均进行3 次生物学重复,每次重复选取5 个单株的叶片混合后进行测量。利用SPAD-502 便携式叶绿素测定仪(日本柯尼卡美能达)测定倒二叶SPAD(soil plant analysis development)值表征叶绿素含量,设3 次生物学重复,每次重复选取5 个单株进行测定。
1.5 数据处理与分析
在Microsoft Excel 2007 中整理试验数据;利用SPSS 20.0 软件以独立样本t检验法 (置信区间95%)进行两组数据的差异显著性分析(存活率、相对株高、相对地上部干物质质量和相对地上中鲜物质质量均通过了正态性检验)。
2 结果与分析
2.1 陆地棉-异常棉异附加系的基因型鉴定
由于MAAL_13B 自交不能产生后代, 为了获得足够多的MAAL_13B 种子, 配制了MAAL_13B×Su8289 杂交组合。陆地棉-异常棉异附加系MAAL_1B~MAAL_12B 的自交后代和MAAL_13B×Su8289 杂交后代的基因型分2种, 即轮回亲本的带型和异附加系的杂合带型(图1)。 最终筛选到的每个单体异附加系均不少于60 株。经统计陆地棉-异常棉异附加系的传递率 为34.66%~51.50% , 其 中MAAL_7B 和MAAL_8B 的传递率较高, 均超过了50%,而MAAL_5B 的传递率最低(表1)。
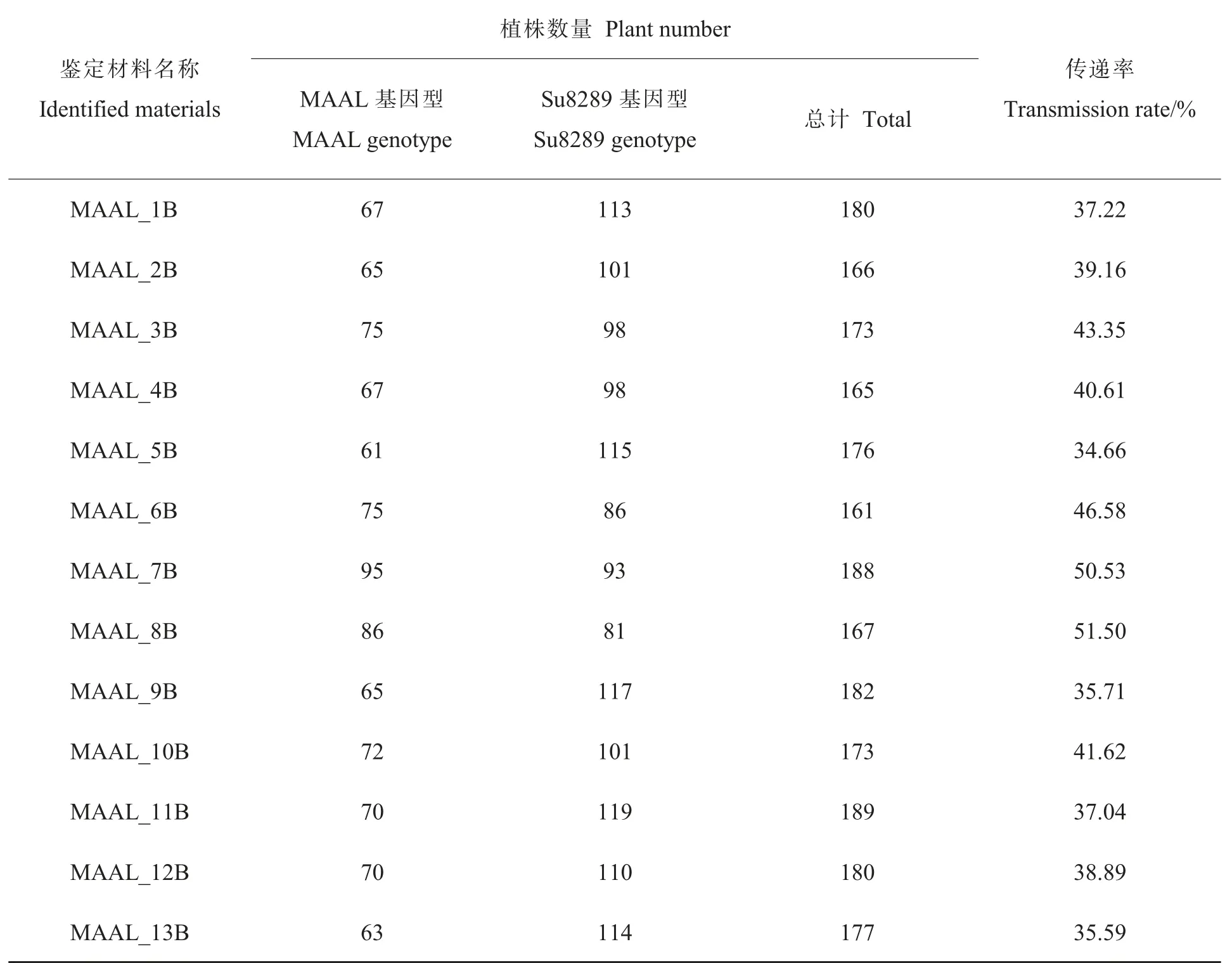
表1 陆地棉-异常棉异附加系的传递率Table 1 Transmission rate of MAALs derived from G. anomalum in G. hirsutum background

图1 部分SSR 引物检测陆地棉-异常棉异附加系的基因型图谱Fig. 1 Amplification patterns of a set of MAALs derived from G. anomalum in G. hirsutum background using G. anomalum-specific SSR markers
2.2 陆地棉-异常棉异附加系的抗旱性鉴定
干旱15 d 后, 轮回亲本陆地棉Su8289 及陆地棉- 异常棉异附加系MAAL_2B、MAAL_3B、MAAL_4B、MAAL_6B、MAAL_9B、MAAL_10B和MAAL_13B 的植株已干枯;复水后1 d,除了MAAL_2B 有较低的存活率外 (3.33%), 其他7份材料全部死亡,存活率为0。 陆地棉- 异常棉异 附 加 系MAAL_1B、MAAL_5B、MAAL_7B、MAAL_8B、MAAL_11B 和MAAL_12B 在 干 旱15 d 后仅出现萎蔫且仍具有生长点, 复水后1 d的存活率为83.33%~96.67%, 极显著高于Su8289(图2)。 通过表型观察和复水后的存活率统计,推测陆地棉- 异常棉异附加系MAAL_1B、MAAL_5B、MAAL_7B、MAAL_8B、MAAL_11B和MAAL_12B 可能具有较强的抗旱性。
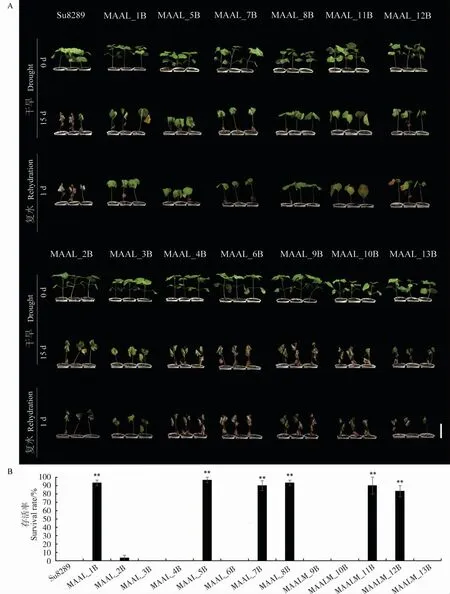
图2 干旱处理下轮回亲本和陆地棉-异常棉异附加系的表型(A)及存活率(B)Fig. 2 Phenotype (A) and survival rate (B) of the recurrent parent and MAALs derived from G. anomalum in G. hirsutum background under drought treatment
2.3 陆地棉-异常棉异附加系的耐盐性鉴定
清水处理下,所有植株均能正常生长,而在350 mmol·L-1NaCl 处理后7 d,所有材料的幼苗生长均受到了不同程度的抑制,叶片也受到了不同程度的损伤。 其中轮回亲本Su8289 部分子叶脱落,未脱落的子叶发黄;异附加系MAAL_2B、MAAL_7B、MAAL_11B 和MAAL_12B 的子叶已脱落,MAAL_8B 和MAAL_9B 的子叶状态较好,仅部分发黄,而其他异附加系材料表现为子叶部分脱落或子叶和真叶出现不同程度的发黄(图3)。
NaCl 处 理 后7 d,MAAL_2B、MAAL_3B、MAAL_4B、MAAL_6B、MAAL_7B、MAAL_8B、MAAL_9B、MAAL_10B 和MAAL_11B 的相对株高显著或极显著高于Su8289。MAAL_4B、MAAL_6B、MAAL_8B、MAAL_9B 和MAAL_10B 的相对地上部鲜物质质量显著或极显著高于Su8289,而MAAL_2B 的相对地上部鲜物质质量极显著低于Su8289。 MAAL_4B、MAAL_6B、MAAL_8B、MAAL_9B 和MAAL_10B 的相对地上部干物质质量极显著高于Su8289, 而MAAL_3B 和MAAL_11B 的相对地上部干物质质量极显著低于Su8289(图3)。 综合以上分析结果,推测陆地棉- 异常棉异附加系MAAL_4B、MAAL_6B、MAAL_8B、MAAL_9B 和MAAL_10B 可能具有较强的耐盐性。

图3 盐胁迫处理后7 d 轮回亲本和陆地棉-异常棉异附加系的表型(A)及相关性状分析(B)Fig. 3 Phenotype (A) and salt related traits analysis (B) of the recurrent parent and MAALs derived from G. anomalum in G. hirsutum background after 7 d of salt stress
2.4 干旱胁迫下陆地棉- 异常棉异附加系生理生化指标测定
与0 d 相比,干旱胁迫处理后10 d,轮回亲本Su8289 与陆地棉- 异常棉异附加系MAAL_1B、MAAL_5B、MAAL_7B、MAAL_8B、MAAL_11B和MAAL_12B 倒二叶中的丙二醛和H2O2的含量均有不同程度地增加,表明干旱胁迫下不同材料可能发生了不同程度地膜脂过氧化。 干旱胁迫处理后10 d, 与Su8289 相比,6 个异附加系叶片中的丙二醛和H2O2含量均极显著降低, 表明干旱胁迫下Su8289 膜脂过氧化程度加剧, 叶片受害程度更重;而抗氧化酶类活性和脯氨酸含量则呈现出不同的变化趋势,SPAD 值出现不同程度地升高。 干旱处理后10 d, 与Su8289 相比,MAAL_1B 中CAT 和SOD 的活性极显著升高,MAAL_5B 中CAT 和SOD 的活性及脯氨酸含量均极显著升高,MAAL_7B 中SOD 活性极显著升高,MAAL_8B 中POD、CAT 和SOD 的 活 性 及SPAD 值极显著或显著升高,MAAL_11B 中POD和CAT 的活性极显著升高,MAAL_12B 中CAT活性及SPAD 值极显著升高(图4)。 由此推断,6种具有抗旱潜力的陆地棉-异常棉异附加系可能通过不同的生理生化过程响应干旱胁迫。
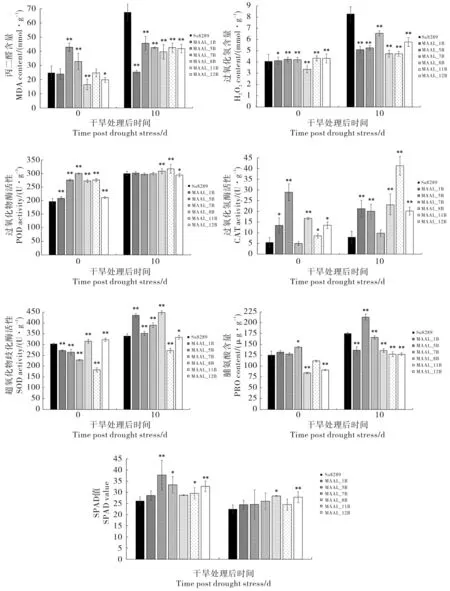
图4 干旱胁迫下轮回亲本和陆地棉-异常棉异附加系的生理生化指标分析Fig. 4 Analysis of physiological and biochemical indexes for the recurrent parent and MAALs derived from G. anomalum in G. hirsutum background under drought stress
2.5 盐胁迫下陆地棉- 异常棉异附加系生理生化指标测定
与清水对照相比,350 mmol·L-1NaCl 处理后3 d,异附加系MAAL_4B、MAAL_6B、MAAL_8B、MAAL_9B 和MAAL_10B 及轮回亲本Su8289倒二叶中的丙二醛和H2O2的含量均有不同程度地升高,表明盐胁迫下不同材料发生了不同程度的膜脂过氧化(图5)。350 mmol·L-1NaCl 处理后3 d , 与Su8289 相比,MAAL_4B 和MAAL_6B中的丙二醛和H2O2含量显著或极显著降低,表明盐胁迫下Su8289 膜脂过氧化程度加剧, 叶片受害程度更重;5 个异附加系中脯氨酸含量均极显著降低;抗氧化酶类活性和SPAD 值则呈现出不同的变化趋势(图5)。350 mmol·L-1NaCl 处理后3 d, 与Su8289 相 比,MAAL_6B 中POD 和SOD 的活性及SPAD 值显著或极显著升高,MAAL_8B 中CAT 活性显著升高,MAAL_9B 中POD 和SOD 的活性极显著升高,MAAL_10B 中SOD 活 性 及SPAD 值 均 极 显 著 升 高, 而MAAL_4B 中SOD 活性、脯氨酸含量及SPAD 值均极显著降低(图5)。 综合以上结果推测5 种具有耐盐潜力的陆地棉-异常棉异附加系MAAL_4B、MAAL_6B、MAAL_8B、MAAL_9B 和MAAL_10B 可能通过不同的生理生化过程响应盐胁迫。

图5 盐胁迫下轮回亲本和陆地棉-异常棉异附加系的生理生化指标分析Fig. 5 Analysis of physiological and biochemical indexes for the recurrent parent and MAALs derived from G. anomalum in G. hirsutum background under salt stress
3 讨论
棉花是一种中等耐盐和抗旱作物,研究人员已通过形态学和生理生化指标测定、 转录组分析、蛋白质组学、关联分析和分子生物学等手段广泛研究了陆地棉栽培种的耐盐性和抗旱性[19-25]。总体上看,我国现有的陆地棉品种均是在正常环境条件下选育的,其抗旱耐盐性有待提高。 棉属野生种为棉花育种提供了丰富的基因资源,从野生棉渐渗外源基因进入陆地棉基因组是陆地棉遗传改良行之有效的方法,其中与四倍体黄褐棉(G.mustelinum)、达尔文氏棉和毛棉(G.tomentosum)相关的研究报道较多[26-28]。 国内外也有不少关于利用二倍体野生棉对陆地棉进行遗传改良的研究报道,如利用哈克尼西棉创造了棉花细胞质雄性不育系[29];利用瑟伯氏棉(G.thurberi)改良陆地棉的纤维品质[30];利用异常棉创制出一批纤维强度较高的棉花种质系7235 和7250 等[31-32];分别将控制植株高酚和种子低酚性状的染色体片段从斯特提棉(G.sturtianum)渐渗到陆地棉[33];将控制肾形线虫抗性的染色体片段从旱地棉(G.aridum)和长萼棉(G.longicalyx)渐渗到陆地棉中,进而提高了陆地棉对肾形线虫的抗性[34-35]。
为了挖掘二倍体野生种异常棉的优异基因或位点,前期研究中,本实验室利用远源杂交和分子标记辅助选择等技术,获得了二倍体野生种的高质量渐渗系群体[17]和一套完整的陆地棉- 异常棉异附加系材料[15]。 本研究系统评价了陆地棉-异常棉异附加系苗期的抗旱性和耐盐性, 发现MAAL_1B、MAAL_5B、MAAL_7B、MAAL_8B、MAAL_11B 和MAAL_12B 具有较强的抗旱性,MAAL_4B、MAAL_6B、MAAL_8B、MAAL_9B和MAAL_10B 具有较强的耐盐性,MAAL_8B同时具有较强的抗旱性和耐盐性,推测异常棉1、5、7、8、11 和12 号染色体存在可用于改良陆地棉抗旱性的优异位点(基因),而4、6、8、9 和10 号染色体存在可用于改良陆地棉耐盐性的优异位点(基因)。 陆地棉-异常棉异附加系不能稳定遗传,自交后代发生基因型分离,因此进行耐盐性
和抗旱性等性状鉴定时需提前进行基因型检测。在今后研究中,将系统评价可能含有抗旱或耐盐优异位点(基因)的、且可稳定遗传的异常棉渐渗系并挖掘关键位点(基因),从而为棉花抗旱耐盐性的遗传改良提供重要的材料与基因资源。
当植物遭受干旱或盐胁迫时会产生大量的活性氧(如H2O2等),活性氧积累可诱发膜脂过氧化, 丙二醛则是膜脂过氧化的产物之一[36-38]。为了消除活性氧造成的伤害,植物会通过改变自身的一些生理生化特性(如通过提高CAT、POD和SOD 等的活性清除细胞中多余的活性氧,降低膜脂过氧化水平,减轻膜受害程度,进而维持胞内稳态)有效适应干旱或盐胁迫环境[39-43]。 各种有机与无机物质 (如可溶性糖和游离的脯氨酸等)的积累可提高胞间浓度,降低细胞渗透势,使植物在低渗条件下仍能正常吸收水分等。 李少昆等[44]研究发现,随着干旱胁迫程度的加重,棉花叶片颜色逐渐由绿变灰,叶绿素含量明显下降。 本研究中,干旱或盐胁迫下轮回亲本和部分陆地棉-异常棉异附加系叶片中的丙二醛和H2O2含量与对照处理相比均有不同程度地增加,且轮回亲本叶片中的丙二醛和H2O2含量更高, 说明其膜脂过氧化程度加剧,受害程度更重。陆地棉-异常棉异附加系材料可能通过不同的生理生化过程响应干旱或盐胁迫环境,MAAL_1B、MAAL_7B、MAAL_11B 的抗旱性和MAAL_9B 的耐盐性可能与抗氧化酶的活性有关,MAAL_5B 的抗旱性可能与抗氧化酶的活性和脯氨酸含量有关,MAAL_12B 的抗旱性和MAAL_6B、MAAL_10B的耐盐性可能与抗氧化酶的活性及叶绿素含量有关。 盐胁迫处理后, 与轮回亲本相比,MAAL_4B 叶片中的SOD 活性、叶绿素含量和脯氨酸含量均极显著降低,推测MAAL_4B 可能通过其他机制应答高盐胁迫。 MAAL_8B 既抗旱又耐盐,其抗旱和耐盐性可能与抗氧化酶的活性有关, 其抗旱性还可能与叶绿素含量有关, 推测MAAL_8B 的抗旱性和耐盐性可能既存在相似的机制,又有各自特异的分子机制。
4 结论
苗期,陆地棉- 异常棉异附加系MAAL_1B、MAAL_5B、MAAL_7B、MAAL_8B、MAAL_11B和 MAAL_12B 的 抗 旱 性 较 强,MAAL_4B、MAAL_6B、MAAL_8B、MAAL_9B 和MAAL_10B 的耐盐性较强。 陆地棉- 异常棉异附加系可能通过不同的生理生化过程应答干旱或高盐胁迫。 MAAL_1B、MAAL_7B、MAAL_9B 和MAAL_11B 可能通过调控抗氧化酶活性响应干旱或高盐胁迫;MAAL_5B 可能通过调控抗氧化酶活性和脯氨酸含量应答干旱胁迫;MAAL_6B、MAAL_10B 和MAAL_12B 可能通过调控抗氧化酶活性与叶绿素含量响应干旱或盐胁迫环境;MAAL_8B 可能通过调控抗氧化酶活性与叶绿素含量响应干旱胁迫,通过调控抗氧化酶活性响应高盐胁迫。
附表:
详见本刊网站(http://journal.cricaas.com.cn/)本文网页版。
附表1 试验所用SSR 引物
Table S1 SSR primers for the experiment
