杂草稻资源表型多样性分析及综合评价研究
2022-12-16蔡海滨韩光煜傅泳维王云月卢宝荣
蔡海滨,韩光煜,涂 敏,官 鑫,汪 智,傅泳维,王云月,卢宝荣*
杂草稻资源表型多样性分析及综合评价研究
蔡海滨1,2,韩光煜1*,涂 敏2,官 鑫2,汪 智3,傅泳维2,王云月1,卢宝荣3*
1. 云南农业大学农业生物多样性与病虫害控制教育部重点实验室/云南农业大学云南生物资源保护与利用国家重点实验室,云南昆明 650201;2. 中国热带农业科学院橡胶研究所,海南海口 571101;3. 复旦大学生命科学学院/生物多样性与生态工程教育部重点实验室,上海 200438
杂草稻是生长形态与栽培稻相似,而稻米质量、落粒性和休眠性等性状不符合栽培稻生产需求的一类杂草。江苏省是我国杂草稻危害最严重的省份之一,大量研究主要集中在杂草稻的起源、性状和防控,杂草稻群体作为种质资源用于栽培稻品种改良尚无系统研究。丰富的遗传多样性和优良种质资源是栽培稻育种以及品种改良中最重要的物质基础,本研究收集21个江苏省不同地区的杂草稻群体、6个栽培稻品种和2个杂草稻对照群体为材料,通过在海南同质园种植,测定18个表型性状,分析其性状的多样性,并进行相关性分析、主成分分析、聚类分析和综合评价研究。结果表明:杂草稻的齐穗期和齐花期均明显早于栽培稻,杂草稻的实粒数和总粒数均较优于栽培稻,杂草稻资源群体表型性状变异系数为2.99%~196.29%,多样性指数为0.27~2.06,多样性指数最高的是结实率,其次为实粒数、剑叶长/宽和总粒数。相关性分析表明,18个表型性状之间大部分存在显著或极显著相关,始穗期和始花期与株高、穗长、剑叶长宽比、总粒数、实粒数、千粒重、分蘖数间均呈极显著正相关,千粒重与穗长、剑叶长宽比、分蘖数、始穗期、始花期呈极显著正相关。主成分分析将18个表型性状集中在累计贡献率为70.57%的6个主成份,其中正相关影响最大的性状是杂草稻的始穗期、始花期、齐穗期、齐花期、总粒数、实粒数等性状。聚类分析将23个杂草稻群体在欧式距离为5时分为5大类。18个指标综合评价值>0.68的有3个群体,其中,来自南通如皋WJ-NJ1的值最高(0.715),其次是来自常州金坛WJ-CZ1(0.698)和盐城市辖区WJ-YC2(0.682)。本研究为杂草稻种质资源在未来的辅助育种以及栽培稻品种改良等方面提供了重要的理论依据。
杂草稻;种质资源;表型性状;遗传多样性;综合评价
杂草稻的广义概念泛指生长在农田生态系统范围中,与农作物争夺生存资源,并且在分类学上属于禾本科稻属()的一类植物[1]。杂草稻广泛地分布于全球栽培稻田[2–5],由于其强烈落粒性、休眠性和生长优势,对栽培稻的生产造成了严重危害[6]。我国江苏省的水稻生产在中国商品粮生产中占据重要的地位,近年拥有超过220万hm²的水稻耕作面积[7]。由于江苏省的栽培稻种植方式为种子直播,成为我国杂草稻发生危害最严重的省份之一[8-9]。戴伟民等[8]于2005—2007年初次对江苏省杂草稻进行全面调查时,发现江苏省的杂草稻仅在该省的中部和南部地区发生严重,北部地区危害较小;2009年,再次调查时,发现江苏省的杂草稻发生面积已经达到20万hm2,约占直播稻面积的四分之一[10]。通常,杂草稻可以造成水稻平均减产20%左右,但杂草稻发生严重的田块,可导致减产60%~80%[10]。由此可见,杂草稻已对江苏省水稻生产造成严重危害,不仅影响栽培稻的产量和品质,直接降低农民收入,而且杂草稻危害的进一步扩大及恶化,还严重影响了水稻新品种的应用和推广[11]。
学者针对杂草稻的特性、防治与起源等方面开展研究,江苏杂草稻在性状表现和抗性方面均比栽培稻更具优势[12-13],而且江苏杂草稻还蕴含丰富的遗传多样性[14-15]。吴川等[12]研究发现,江苏省杂草稻遗传资源丰富,性状变异丰富,研究结果还表明在生殖生长期杂草稻的粒长、粒宽、有效穗数等性状显著优于栽培稻。此外,杂草稻对比同期栽培稻具有抗性强、适应快、生长旺盛等特点[9]。李玉融等[16]选择了江苏中部、南部、北部地区的3份杂草稻及2份常规稻进行温度、水势等条件下种子萌发能力的影响研究,发现在12.5℃恒温和水势大于‒1.05 MPa的条件下,杂草稻的种子萌发率均高于常规栽培稻,表现出一定程度的耐旱性和耐低温性。张峥等[10]对江苏沿江地区杂草稻进行研究的结果也表明了其分蘖数、落粒率等性状具有优势,生育期比栽培稻短20 d左右。代磊等[17]对江苏采集杂草稻样品与当地栽培稻的23个性状进行分析,发现杂草稻的分蘖数、芒长、落粒率均优胜过栽培稻,且其生长速率更快。
综上所述,江苏省的杂草稻除了具有其杂草的特性,由于其对环境的适应性和农艺性状的优异表现,也可以考虑作为栽培稻的遗传改良的一类新的种质资源和育种材料。因此,对其表型性状及其变异进行评价和研究具有十分重要的意义。目前已报道对江苏省杂草稻的研究,主要集中在不同耕作措施下杂草稻的危害特征以及杂草稻的起源等方面[18-21],而对江苏省杂草稻群体资源进行综合评价的相关研究鲜见报道。本研究以江苏省的杂草稻群体和栽培稻为实验材料,对其进行同质园种植,并测量分析了18个表型性状,旨在综合评价江苏省各地区不同杂草稻群体的表型变异、群体间的差异程度以及与栽培稻性状间的差异,为筛选杂草稻的优异性状作为后续栽培稻品种遗传改良的种质资源材料提供理论依据。
1 材料与方法
1.1 材料
供试杂草稻资源为江苏省各地区的21个杂草稻群体,对照为1个吉林省粳稻型杂草稻群体和1个广东省籼稻型杂草稻群体,各杂草稻群体样本数为30个;同时选择6个栽培稻对比品种,分别为江苏省的‘连粳7号’(G-LG7)、‘淮稻5号’(G-HD5);广东省的‘黄华占’(X-HHZ)、‘粤秀占’(X-YXZ);吉林省的吉‘粳301’(G-JG301)和‘绥粳18’(G-SG18)。杂草稻资源群体的名称、采集地点与经纬度等信息详见表1。所有材料于2018年5月种植于海南省儋州市宝岛新村沙田实验基地,每个群体以7行8列共56株为一个种植小区,行间距为15 cm´30 cm,设置3个重复;按常规稻田管理办法进行日常管理。

表1 本研究杂草稻群体的采样点信息
1.2 方法
按照韩龙植[22]《水稻种质资源描述规范和数据标准》的统一规范和标准,29个群体去除边界株行,选择中间30株,3个重复,每个群体共选90株进行18个表型性状的调查,主要包括:株高(plant height, PH)、第一节间长(first intersegmental length, FIL)、叶毛(leaf Hair, LS)、剑叶长宽比(sword leaf aspect ratio, BLTWR)、分蘗数(tiller number, TN)、始穗期(first panicle, FP)、齐穗期(flush heading stage, FHS)、始花期(first flowering stage, IFS)、齐花期(flush flowering stage, FFS)、穗长(panicle length, PL)、总粒数(total grains number, TGN)、实粒数(number of solid grains, SN)、结实率(seed sitting rate, SSR)、千粒重(thousand sub-grain weight, TSW)、稃毛(leaf hair, LH)、稻壳颜色(rice husk color, RHC)、酚反应(phenol reaction, PR)、落粒性(shattering rate, SP)进行调查。
1.3 数据处理

2 结果与分析
2.1 杂草稻群体与栽培稻品种表型性状比较分析
如表2所示,在18个性状中,杂草稻的剑叶长/宽、始花期变异系数(coefficient of variation,CV)较小,为2.44%~9.59%。稃毛的变异系数较大,为305.19%。杂草稻和栽培稻除了剑叶长/宽与酚反应无显著差异外,其他性状之间都有显著差异。在同质园中,杂草稻齐穗期和齐花期都明显早于栽培稻(10.61<11.24;15.22<16.15),杂草稻实粒数和总粒数都较优于栽培稻(233.87> 217.59;169.12>152.44),这表明在环境变化过程中,杂草稻的环境适应性要强于栽培稻。

表2 杂草稻群体与栽培稻群体表型性状比较
注:同列数据后不同小写字母表示差异显著(<0.05)。
Note: Different lowercase letters after the same column of data indicate significant difference (<0.05).
2.2 29个群体材料主要表型性状的遗传变异
供试材料的18个主要表型性状的均值、标准差、极值、变异幅度、变异系数及多样性指数见表3。表型性状变异系数的变化范围从2.99%(剑叶长/宽)到196.29%(稃毛),各表型性状差异明显,其中稃毛(196.29%)、酚反应(84.54%)与稻壳色(70.70%)的变异系数较大,均在50%以上,说明这3个表型性状具有较大的遗传变异,研究价值较高。多样性指数表明剑叶长/宽、每穗总粒数、每穗实粒数、结实率多样性指数均较高,分别为2.00、2.02、2.04、2.06,说明这个4个性状的多样性最丰富,而稃毛的多样性指数最低,为0.27。由此可见,供试材料主要的表型性状具备丰富的表型多样性,具有较大的挖掘潜力,可作为改良的优质种质资源库。
2.3 表型性状之间的相关性分析
供试材料的18个主要表型性状的相关分析结果(表4)表明,不同表型性状间存在不同程度的相关性,大部分性状表现为显著或极显著相关。始穗期与始花期、齐穗期与齐花期呈极显著正相关关系,同时株高、穗长、剑叶长宽比、总粒数、实粒数、千粒重、分蘖数与始穗期和始花期呈极显著正相关,稃毛、叶毛、稻壳颜色、酚反应、落粒性、第一节间长与始穗期和始花期呈极显著负相关;穗长、剑叶长宽比、分蘖数、始穗期、始花期与齐穗期、齐花期呈极显著负相关,第一节间长、稃毛、稻壳颜色、酚反应、落粒性与齐穗期和齐花期呈极显著正相关。株高与穗长、剑叶长宽比、结实率、分蘖数、酚反应、始穗期、始花期呈极显著正相关,与第一节间长、总粒数、稃毛、叶毛、稻壳颜色、落粒性呈极显著负相关。结实率与株高、实粒数呈极显著正相关,与分蘖数显著相关,与穗长显著负相关,因此在水稻产量选育种过程中,需要充分考虑株高和分蘖数等性状。千粒重与穗长、剑叶长宽比、分蘖数、始穗期、始花期呈极显著正相关,与第一节间长呈极显著负相关,因此如果选育大粒的水稻品种时,应该选取穗长、剑叶长宽比大且第一节间长不宜过长的性状聚集。在育种过程针对目标性状需求,根据各性状的相互关系,协调性状之间相互关系,选取最优的方案聚集性状。
2.4 表型性状主成分分析
对29份供试材料的18个表型性状进行主成分分析,表5显示其Kaiser-Meyer-Olkin检验值为0.635,Bartlett球形度检验的概率为0,表明变量间具有极显著相关性,从中萃取出因子特征值大于1的主成分有6个,结果显示(表6),前6个主成分的累积贡献率为70.57%,表明这18个表型性状作为变量对供试材料进行数量分类是可靠的。第1主成分的特征值和贡献率分别为5.63和31.30%,成分为始穗期因子(0.88);第2主成分的特征值和贡献率分别为2.04和11.31%,主成分为总粒数(0.94)、实粒数(0.86);第3主成分的特征值和贡献率分别为1.74和9.64%,主成分为齐花期(0.86)和齐穗期(0.84);第4主成分的特征值和贡献率分别为1.17和6.48%,成分为结实率因子(0.83);第5主成分的特征值和贡献率分别为1.10和6.09%,主成分为千粒重因子(0.64);第6主成分的特征值和贡献率分别为1.04和5.77%,主成分为结实率因子(0.33)。供试材料的18个性状中,始穗期、始花期、齐穗期、齐花期、总粒数、实粒数、千粒重、结实率和叶毛为重要的表型性状指标,在综合评价杂草稻表现应着重考虑这9个指标。
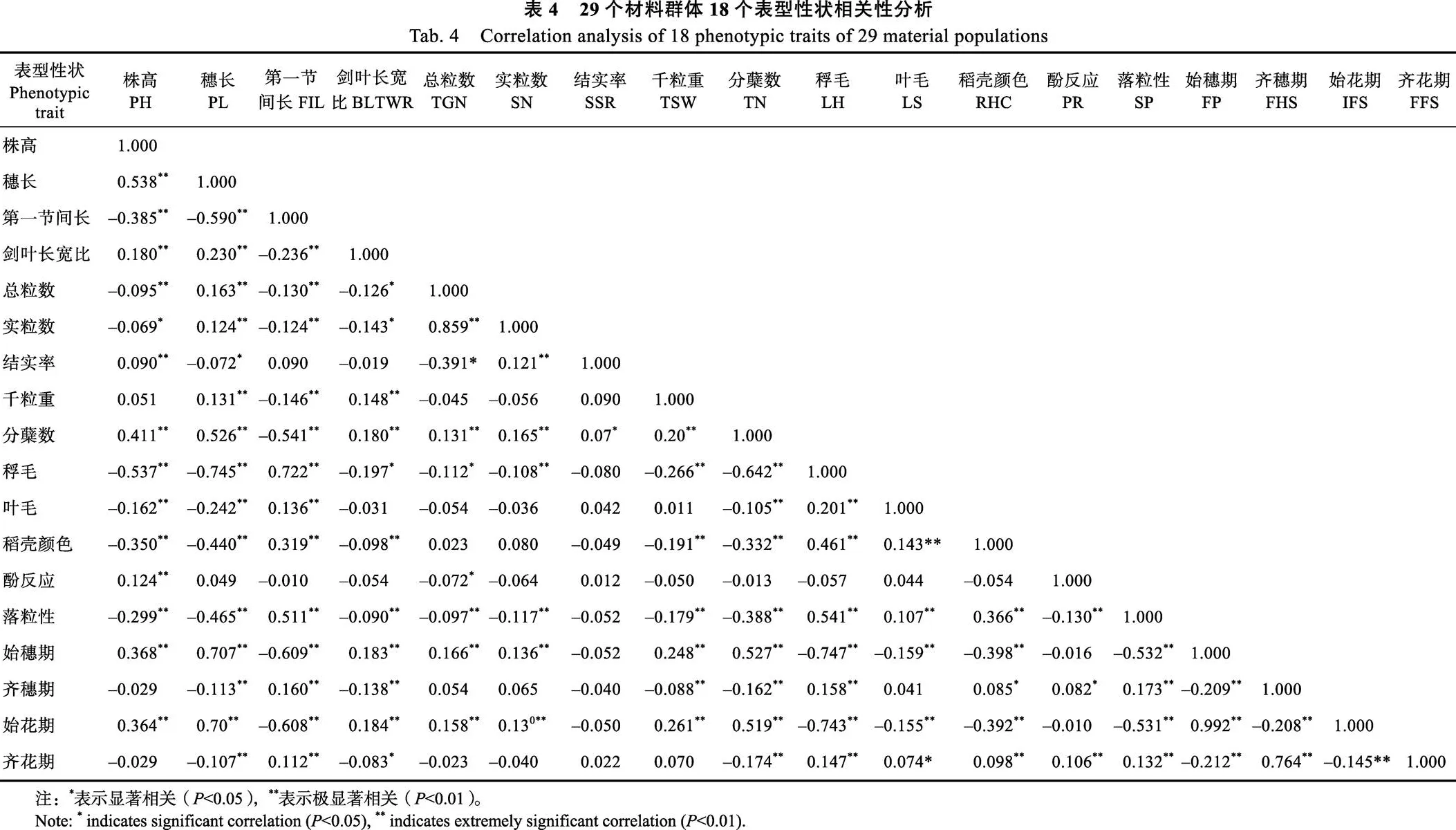

表5 KMO和Bartlett的检验

表6 29个群体材料18个表型性状主成分分析
2.5 杂草稻群体的聚类分析
由所测得的18个表型数据对2份杂草稻对照材料和21份江苏杂草稻材料进行聚类分析,聚类结果如图1所示,在欧式距离为5时,可以分成5个类群,其中第Ⅰ类群包括19份材料;第Ⅱ类群仅有一份来自泰州泰兴的杂草稻群体(WJ-TZ);第Ⅲ类群仅有一份材料徐州睢宁的杂草稻群体(WJ-XZ2);第Ⅳ类群仅有一份材料淮安的杂草稻群体(WJ-XA2);第Ⅴ类群仅有一份材料吉林的杂草稻群体(WC-JL)。
2.6 杂草稻优异资源的筛选
根据主成分分析的结果,计算23份杂草稻资源的综合评价值。依据各主成分贡献率的值分别除以前6个主成分的累积贡献率(70.57%)得到前6个主成分的权重(0.291、0.179、0.143、0.115、0.094、0.093)。再根据权重计算得到来自23份杂草稻资源的综合评价值,值的高低代表此杂草稻资源18个主要表型综合性状的优劣,并根据值的高低对23份杂草稻资源进行综合排名(表7)。从表6中可以看出,综合排名前3名依次为WJ-NJ1、WJ-CZ1、WJ-YC2。来自南通如皋WJ-NJ1的值最高(0.715),来自吉林WC-JL的值最低(0.484),说明WJ-NJ1的表型综合性状较好,可以作为选育优质水稻材料的亲本和中间材料。
以18个表型性状数值与综合得分值构建最优回归方程,筛选杂草稻种质资源最优评价指标。以18个表型性状数值为自变量,以值为因变量,通过逐步线性回归方法构建的最优回归方程式为:=‒0.036+0.0091+0.0012‒0.0563+0.014X‒0.0165+0.0276+0.0097+0.0008‒ 0.0059,方程式中1、2、3、4、5、6、7、8、9分别代表始穗期、实粒数、第一节间长、齐花期、落粒性、叶毛、分蘖数、株高、始花期9个表型性状,方程式的相关系数和判定系数2分别为0.999和0.997,说明这9个自变量可以解释值99.7%的总变异,值为943.316,方程表示为极显著结果。由回归方程可知,在18个表型性状中,始穗期、实粒数、第一节间长、齐花期、落粒性、叶毛、分蘖数、株高、始花期9个性状对杂草稻种质资源表型性状多样性综合值的影响显著,说明其可以作为水稻种质资源综合评价指标。

聚类图最左侧1~23分别对应23份材料的编号。
3 讨论
杂草稻具有落粒性强、发育早、适应能力强等特点[25-27],如果针对育种目标,深入挖掘杂草稻中有利性状进行育种,将优良基因整合到栽培稻中,将会优化其生物学特性[28],提升栽培稻适应不良环境及抵抗病虫害能力,缩短生育期提早开花,提高水稻产量品质等。因此需要广泛开展对杂草稻优良性状基因的收集和保存工作,进一步为水稻研究提供优质的种质基础。本研究对杂草稻和栽培稻的部分性状进行比较,发现杂草稻的落粒性是区别于栽培稻的主要性状之一,除此之外,杂草稻比栽培稻生长速率更快,分蘖数量多,同时植株普遍高过栽培稻。LEOPOLDO等[29]的研究同样也证明了杂草稻苗期生长迅速,分蘖多,植株较为高大且落粒性强的优良特性,说明杂草稻某些表型性状不仅优于当地栽培稻,而且具有极强的变异能力,可作为水稻品种改良的优质基因库。

表7 23个杂草稻群体的综合评价D值与排名
水稻各表型性状之间存在一定的相互关系。陈能刚等[30]发现全生育期与株高、每穗粒数呈显著相关关系,与穗长和穗实粒数呈极显著相关关系,每穗实粒数与粒长宽比、千粒重、结实率呈极显著正相关。杨久等[31]通过对云南籼粳交错区水稻表型性状进行相关性分析,发现每穗总粒数与每穗实粒数,单株粒重与千粒重呈显著正相关,单株粒重与千粒重呈显著正相关,生育期和穗长没有达到显著相关。温广月等[32]对上海地区杂草稻表型性状进行相关性分析,发现生育期与穗长呈显著负相关、与成熟期呈极显著正相关;株高与分蘖角呈显著负相关、与一次枝梗数呈正相关,穗长与千粒重、一次枝梗数、穗粒数和籽粒长宽比呈正相关;一次枝梗数与穗粒数呈正相关。李鑫等[33]研究发现穗粒数与生育期、有效穗、千粒重、结实率呈显著的负相关关系,与着粒密度、单株粒重呈显著或极显著的正相关关系。粒重与生育期、结实率和单株粒重呈显著或极显著正相关,与穗长、穗粒数、有效穗、着粒密度和结实率间呈显著或极显著的负相关关系;李翔等[34]对12个京津唐水稻区试验品种分析,发现穗长、总粒数、实粒数互相之间呈极显著正相关,有效穗与总粒数、实粒数均呈极显著负相关,分蘖率和有效穗数呈极显著正相关。综上所述,可以发现水稻各表型性状间存在着复杂的相互关系,导致与前人研究结果的不同,可能与供试材料的类型、试验地理位置、栽培措施和地区气候等条件有关。本研究发现杂草稻资源中,始穗期与始花期、齐穗期与齐花期呈极显著正相关关系,稃毛、叶毛、稻壳颜色、酚反应、落粒性、第一节间长与始穗期和始花期呈极显著负相关;育种时可以参考相关系数来选择,若想要提前抽穗,就要选择增加始花期这一性状;若要结实率高且粒大的,就要选第一节间长较短的。在水稻品种选育和改良工作中,针对不同的目标需求,依据性状间的相互关系,增强相关目标性状基因的表达,从而达到优质、高产、高抗的目的。
主成分分析法应用降维思维,将原有多个变量转化为个数较少的综合指标,可以避免重复信息干扰、排除主观因素影响,是对种质资源进行综合评价的前提。本研究将18个性状分为6个主成分,其特征值均大于1,累积贡献率70.57%,稃毛、子粒数、齐花期、结实率、千粒重、酚反应,这6个特征性状导致了杂草稻种质性状的多样性。这与代磊等[17]和吴川等[12]主成分分析结果类似。在育种工作中,要通过主成分信息,抓住主要因子,这样有利于更快选育出新品种。
本研究对江苏省部分杂草稻群体资源进行遗传多样性分析,发现其表型性状具有丰富的多样性,随后对其进行综合评价,发现南通如皋WJ-NJ1、常州金坛WJ-CZ1、盐城市辖区WJ-YC2这3个群体的表型性状的综合表现较好,始穗期、实粒数、第一节间长、齐花期、落粒性、叶毛、分蘖数、株高、始花期9个性状能显著影响杂草稻种质资源表型性状多样性,应充分利用以上表型性状,加大力度对杂草稻种质资源进行综合评价,为栽培稻优异种质的选育提供参考依据。
[1] CHEN L J, LEE D S, SONG Z P, SUH H S, LU B R. Gene flow from cultivated rice () to its weedy and wild relatives[J]. Annals of Botany, 2004, 93(1): 67-73.
[2] KARN E, DE L T, ESPION L, Al K K, BRIM D W. Effects of competition from california weedy rice (f. spontanea) biotypes on a cultivated rice variety[J]. Weed Technology, 2020, 34(5): 666-674.
[3] MAHMOD I F, SAIMAN M Z, MOHAMED Z, ISHAK M N, MISPAN M S. Morphological variation, distribution and relationship of weedy rice (L.) in Peninsular Malaysia[J]. Weed Biology and Management, 2021, 21(2): 86-99.
[4] PIVETA L B, NOLDIN J A, ROMA B N,VIANA V E, BEBEDETTI L, OLIVERIRA P J J, LAMEGO F P, DE A L A. Weedy rice (spp.) diversity in Southern Brazil[J]. Weed Science, 2021, 69(5): 547-557 .
[5] SUSHMITA M, SANJOY S, TOTAN A, NITIPRASAD J, PRIYADARSINI S, BHASKAR C P. Performance of cultivated indica rice (L.) as affected by weedy rice[J]. Experimental Agriculture, 2019, 55(6): 875-884
[6] DILIPLUMAR M, KUAMR V, SONG B K ,OLSEN K M, CHUAH T S, AHMED S, QIANG S. Weedy rice (spp.)[M]. Amsterdam: Elsevier, 2021: 285-309.
[7] 江苏省农业农村厅. 2020分市主要农作物面积产量表[EB/OL].(2021-12-17)[2022-09-01].http://nynct.jiangsu.gov.cn/art/2021/12/ 17/art_12552_10218090.html.
Jiangsu Provincial Department of Agriculture and Rural Development. 2020 table of major crop area production by city [EB/OL]. (2021-12-17)[2022-09-01].http://nynct.jiangsu.gov.cn/ art/2021/12/17/art_12552_10218090.html. (in Chinese)
[8] 戴伟民, 宋小玲, 吴 川, 张连举, 左然林, 张 峥, 李淑顺, 曹 旦, 左 娇, 杨 琳, 刘琳莉, 强 胜. 江苏省杂草稻危害情况的调研[J]. 江苏农业学报, 2009, 25(3): 712-714.
DAI W M, SONG X L, WU C, ZHANG L J, ZUO R L, ZHANG Z, LI S S, CAO D, ZUO J, YANG L, LIU L L, QIANG S. A study on weed rice damage in Jiangsu Province[J]. Journal of Jiangsu Agriculture, 2009, 25(3): 712-714. (in Chinese)
[9] 于海艳, 单红保. 宿迁市宿城区杂草稻发生特点及防除对策[J]. 杂草科学, 2011, 29(1): 50-52.
YU H Y, SHAN H B. Characteristics of weed rice occurrence and countermeasures of weed control in Suqian City[J]. Weed Science, 2011, 29(1): 50-52. (in Chinese)
[10] 张 峥, 戴伟民, 章超斌, 强 胜. 江苏沿江地区杂草稻的生物学特性及危害调查[J].中国农业科学, 2012, 45(14): 2856-2866.
ZHANG Z, DAI W M, ZHANG C B, QIAN S. Biological characteristics and damage of weedy rice in Jiangsu riverine areas[J]. China Agricultural Science, 2012, 45(14): 2856-2866. (in Chinese)
[11] 尤国林, 陈梅英. 稻区杂草稻的发生规律与防控技术[J]. 农业科技通讯, 2013(4): 201-202.
YOU G L, CHEN M Y. Occurrence patterns and control techniques of weedy rice in rice areas[J]. Agricultural Science and Technology Newsletter, 2013(4): 201-202. (in Chinese)
[12] 吴 川, 戴伟民, 宋小玲, 强 胜. 辽宁和江苏两省杂草稻植物性状多样性[J]. 生物多样性, 2010, 18(1): 29-36.
WU C, DAI W M, SONG X L, QIANG S. Plant trait diversity of weedy rice in Liaoning and Jiangsu provinces[J]. Biodiversity, 2010, 18(1): 29-36. (in Chinese)
[13] DAI L, SONG X L, HE B Y, VALERDE B E, QIANG S. Enhanced photosynthesis endows seedling growth vigour contributing to the competitive dominance of weedy rice over cultivated rice[J]. Pest Management Science, 2017, 73(7): 1410-1420.
[14] NGUYEN T, ZHOU C l, ZHANG T Y, YU F J, WANG J M. Identification of QTL for seed dormancy from weedy rice and its application to elite rice cultivar ‘Ninggeng 4’[J]. Molecular Breeding, 2019, 39(9): 1-12.
[15] JUAN Z, NILDA R, BURGOS, KUN M, YONG J, ZHOU, RUI M, GENG, LIU-QING Y U. Genetic diversity and relationship of weedy rice in Taizhou city, Jiangsu Province, China[J]. Rice Science, 2008, 15(4): 295-302.
[16] 李玉融, 董立尧, 李 俊, 李振博. 苏南苏中苏北地区3个杂草稻生物型萌发特性比较研究[J]. 杂草科学, 2011, 29(2): 14-18.
LI Y R, DONG L Y, LI J, LI Z B. Comparative study on the germination characteristics of three weed rice biotypes in southern and central Jiangsu and Northern Jiangsu[J]. Weed Science, 2011, 29(2): 14-18. (in Chinese)
[17] 代 磊, 戴伟民, 宋小玲, 强 胜. 江苏省杂草稻植物学性状的多样性[J]. 杂草科学, 2014, 32(1): 10-18.
DAI L, DAI W M, SONG X L, QIAN S. Diversity of botanical traits in weedy rice in Jiangsu province[J]. Weed Science, 2014, 32(1): 10-18. (in Chinese)
[18] 李振博, 董立尧, 李 俊, 张宏军, 杨玉清. 不同环境因子对江苏省杂草稻与常规栽培稻发芽势影响的比较[J].杂草科学, 2009(4): 28-31.
LI Z B, DONG L Y, LI J, ZHANG H J, YANG Y Q. Comparison of the effects of different environmental factors on germination potential of weedy rice and conventional cultivated rice in Jiangsu province[J]. Weed Science, 2009(4): 28-31. (in Chinese)
[19] 曹 旦, 戴伟民, 强 胜, 宋小玲. 栽培措施对杂草稻和栽培稻生长的影响[J]. 杂草科学, 2011, 29(3): 14-21, 29.
CAO D, DAI W M, QIANG S, SONG S L. Effects of cultivation practices on the growth of weedy rice and cultivated rice[J]. Weed Science, 2011, 29(3): 14-21, 29. (in Chinese)
[20] 陈 雷, 金 曼, 张维乐, 王承旭, 吴勇斌, 王治忠, 唐晓艳. 杂草稻的特性及其危害与防治研究进展[J]. 作物学报, 2020, 46(7): 969-977.
CHEN L, JIN M, ZHANG W L, WANG C X, WU Y B, WANG Z Z, TANG X Y. Research progress on the characteristics of weedy rice and its damage and control[J]. Journal of Crop Science, 2020, 46(7): 969-977. (in Chinese)
[21] ZHANG B H, ER j D, ZHANG H, SHI Z H, SONG XL, QIANG S, DAI W M. Use of hybrid rice and the proliferation of weedy rice: a case in Jiangsu, China[J]. Crop Science, 2016, 56(2): 673-681.
[22] 韩龙植. 水稻种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2006.
HAN L Z,. Specifications and data standards for the description of rice germplasm resources[M]. Beijing: China Agriculture Press, 2006. (in Chinese)
[23] 李金梅, 崔 迪, 汤翠凤, 阿新祥, 余滕琼, 马小定, 张恩来, 刘昌文, 徐福荣, 戴陆园, 韩龙植. 两个时期收集的云南水稻农家品种表型多样性比较[J]. 植物遗传资源学报, 2015, 16(2): 238-244.
LI J M, CUI D, TANG C F, A X X, YU T Q, MA X D, ZHANG E L, LIU C W, XU F R, DAI L Y, HAN L Z. Comparison of phenotypic diversity of Yunnan rice farming varieties collected in two periods[J]. Journal of Plant Genetic Resources, 2015, 16(2): 238-244. (in Chinese)
[24] 周红灿, 尹伟丹, 揭红东, 马玉申, 曾 怡, 何鹏亮, 邢虎成, 揭雨成. 薏苡种质资源性状评价及遗传多样性研究[J]. 热带作物学报, 2022, 43(7): 1365-1374.
ZHOU H C, YIN W D, JIE H D, MA Y S, ZENG Y, HE P L, XING H C, JIE Y C. Evaluation of traits and genetic diversity ofgermplasm resources[J]. Journal of Tropical Crops, 2022, 43(7): 1365-1374. (in Chinese)
[25] ZHU Y Q, FANG J, WANG Y, PANG L H, LU B R. Key roles of de-domestication and novel mutation in origin and diversification of global weedy rice[J]. Biology, 2021, 10(9): 828-843.
[26] ZHAO C, XU W R, LI H W, DAI W M, ZHANG Z, QIANG S, SONG X L. The rapid cytological process of grain determines early maturity in weedy rice[J]. Frontiers in Plant Science, 2021, 12: 711321-711333.
[27] ZHOU C C, FENG Y, LI G Y, WANG M L, JIAN J J, WANG Y G, ZHANG W J, SONG Z P, LI L F, LU B R, YANG J. The new is old: novel germination strategy evolved from standing genetic variation in weedy rice[J]. Frontiers in Plant Science, 2021, 12: 699464-699474.
[28] NADIR S, XIONG H B, ZHU Q, ZHANG X L, XU H Y, LI J, DONGCHEN W H, HENRY D, GUO X Q, KHAN S, SUH H S, LEE D S, CHEN L J. Weedy rice in sustainable rice production. a review[J]. Agronomy for Sustainable Development, 2017, 37(5): 1-14.
[29] LEOPOLDO E. E, DAVID R G, RONALD E T. Growth response of rice () and red rice () in a replacement series study[J]. Weed Technology, 2002, 16(2): 401-406.
[30] 陈能刚, 陈惠查, 游俊梅, 阮仁超. 贵州旱稻主要农艺性状的相关通径及聚类分析[J]. 贵州农业科学, 2010, 38(11): 46-49.
CHEN N G, CHEN H C, YOU J M, RUAN R C. Correlation paths and cluster analysis of major agronomic traits of dry rice in Guizhou[J]. Guizhou Agricultural Science, 2010, 38(11): 46-49. (in Chinese)
[31] 杨 久, 丁 鲲, 卢义宣, 辜琼瑶, 刘晓利, 张竹仙, 宋建荣, 李华惠. 云南籼粳交错区水稻农艺性状与产量相关及通径分析[J]. 西南农业学报, 2011, 24(2): 391-395.
YANG J, DING K, LU Y X, GU Q Y, LIU X L, ZHANG Z X, SONG J R, LI H H. Agronomic traits and yield correlation and pathway analysis of rice in the indica-japonica mosaic region of Yunnan[J]. Southwest Journal of Agriculture, 2011, 24(2): 391-395. (in Chinese)
[32] 温广月, 沈国辉, 钱振官, 李 涛. 上海地区杂草稻形态学和农艺性状多样性研究[J]. 中国农学通报, 2012, 28(6): 194-201.
WEN G Y, SHEN G H, QIAN Z G, LI T. Morphological and agronomic trait diversity of weedy rice in Shanghai[J]. Chinese Agronomy Bulletin, 2012, 28(6): 194-201. (in Chinese)
[33] 李 鑫, 孙 杰, 张丽丽, 刘 研, 张素红. 辽宁省水稻种质资源主要农艺性状的相关性分析[J]. 辽宁农业科学, 2014(3): 53-56.
LI X, SUN J, ZHANG L L, LIU Y, ZHANG S H. Correlation analysis of major agronomic traits of rice germplasm resources in Liaoning province[J]. Liaoning Agricultural Science, 2014(3): 53-56. (in Chinese)
[34] 李 翔, 赵长海, 苏建功. 水稻主要农艺性状与产量的相关性分析[J]. 天津农林科技, 2020(3): 1-3.
LI X, ZHAO C H, SU J K. Correlation analysis of major agronomic traits and yield of rice[J]. Tianjin Agriculture and Forestry Science and Technology, 2020(3): 1-3. (in Chinese)
Phenotypic Diversity Analysis and Comprehensive Evaluation of Weedy Rice Germplasm
CAI Haibin1,2, HAN Guangyu1*, TU Min2, GUAN Xin2, WANG Zhi3, FU Yongwei2, WANG Yunyue1,LU Baorong3*
1. Key Laboratory for Agricultural Biodiversity and Pest Control of Ministry of Education, Yunnan Agricultural University / State Key Laboratory of Biological Resources Protection and Utilization, Yunnan Agricultural University, Kunming, Yunnan 650201, China; 2. Rubber Research Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan 571101, China; 3. College of Life Sciences, Fudan University / Key Laboratory of Biodiversity and Ecological Engineering, Ministry of Education, Shanghai 200438, China
Weedy rice is a weed similar to cultivated rice in growth form, but does not meet the requirements of cultivated rice production in rice quality, grain setting and dormancy. Jiangsu province is one of the provinces with the most serious damage to weedy rice in China. Many studies have focused on the origin, characters and control of weedy rice. There has no systematic study on weedy rice population as germplasm resources for cultivar improvement. Rich genetic diversity and excellent germplasm resources are the most important material basis for cultivated rice breeding and variety improvement. In the study, 21 weedy rice populations in different regions of Jiangsu Province, six cultivated rice varieties and two weedy rice populations as the control were planted in a Hainan homogeneous garden, and the diversity of 18 phenotypic traits was analyzed. Correlation analysis, principal component analysis, cluster analysis and comprehensive evaluation analysis were performed. The full panicle stage and flowering stage of weedy rice were significantly earlier than that of cultivated rice, and the rice grain number and total grain number of the weedy rice were relatively better than that of the cultivated rice. The phenotypic traits variation coefficient of the weedy rice resources group was 2.99%‒196.29%, and the diversity index was 0.27‒2.06, with the highest diversity index for seed setting rate, followed by solid grain number, sword leaf length/width, and total grain number. The beginning stage of heading and flowering had extremely significant positive relation with plant height, spike length, sword leaf aspect ratio, total grain number, actual grain number, 1000 grain weight and tillering number. 1000 grain weight had extremely significant positive relation with tiller number, spike length, sword leaf aspect ratio, tillering number, beginning stage of heading and flowering. Cumulative contribution 70.57% was from six principal components, including first heading stage, first flowering stage, full heading stage, full flowering stage, total grain number and full grain number. The 23 weedy rice populations were divided into five categories when the Euclidean distance was 5. There were three populations withvalue >0.68 in the comprehensive evaluation of the18 indicators, with thevalue of WJ-NJ1 from Rugao, Nantong the highest (0.715), followed by WJ-CZ1 from Jintan, Changzhou (0.698) and WJ-YC2 from Yancheng Municipal District (0.682). This study would provide an important theoretical basis for weedy rice germplasm resources in future assisted breeding and improvement of cultivated rice varieties.
weedy rice; germplasm; phenotypic traits; genetic diversity; comprehensive evaluation
S511.2
A
10.3969/j.issn.1000-2561.2022.11.012
2022-09-06;
2022-09-30
农业农村部科技教育司购买服务项目“典型区域农业野生植物资源调查收集与监测”(No. A120201);云南省“高层次人才培养支持计划”青年拔尖人才专项(No. YNWRQNBJ2020296)。
蔡海滨(1983—),男,博士研究生,副研究员,研究方向:杂草稻资源筛选与利用。*通信作者(Corresponding author):卢宝荣(LU Baorong),E-mail:brlu@fudan.edu.cn;韩光煜(HAN Guangyu),E-mail:hanguangyu9745@163.com。
