基于高通量测序的昭通核心烟区植烟土壤细菌多样性分析
2022-12-15刘永军陆文林高顺波盘文政李德文朱永立林先亮普晓明
刘永军, 倪 霞*, 陆文林, 龚 林, 陈 华, 高顺波, 盘文政,李德文, 陈 敏, 夏 蕾, 朱永立, 林先亮, 普晓明
(1.云南省烟草公司 昭通市公司, 云南 昭通 657000; 2.云南云叶化肥股份有限公司, 云南 昆明 650217; 3.云南省昭通市植保植检站, 云南 昭通 657000)
0 引言
【研究意义】烟草(Nicotianatabaccum)是云南的主要经济作物之一,烟草生长发育离不开土壤,土壤为烟株生长发育提供充足养分,不合理的耕作方式影响植烟土壤微生物多样性,导致土地可持续生产能力下降[1-2]。土壤中微生物能提高土壤肥力,促进土壤有机质分解,加快养分循环转化,抑制土传病虫害传播等[3-4]。其中,土壤细菌作为土壤微生物中种类和数量最多的类群,其多样性一定程度上反映土壤功能的多样性,土壤细菌多样性通过影响植物营养吸收进而影响其生长发育[5-6]。因此,探明昭通核心烟区植烟土壤中的细菌多样性有利于土壤细菌资源的开发利用,还能为昭通核心烟区烤烟种植提供参考。【前人研究进展】目前,对植烟土壤微生物多样性已有较多研究,陈尧等[5]采用平板培养方法对不同施肥方式下烤烟根际微生物进行研究表明,解磷菌、解钾菌和固氮菌数量在不同施肥方式中有变化,为科学选择施肥方式提供理论依据;何川等[7]采用BIOLOG生态平板法,探究烟草连作对土壤微生物多样性的影响得出,烟草连作年限超过3年,其土壤微生物多样性显著降低;贾志红等[8]利用PCR-DGGE法对云南玉溪烟区轮作与连作植烟土壤细菌多样性进行分析表明,轮作土壤细菌多样性较连作高,且轮作可以提高土壤细菌群落多样性;王飞等[9]应用高通量测序技术研究不同绿肥对植烟土壤细菌群落结构的影响发现,翻压黑麦草处理对细菌种分类水平影响最大,翻压绿肥可以改善土壤细菌群落结构;杨帅等[10]利用Illumina HiSep高通量测序研究不同轮作方式对烟田土壤细菌多样性的影响发现,烟草-青蒿轮作能显著降低烟草黑胫病的发病率,而烟草-田菁轮作会提高烟草青枯病的发病率;齐虹凌等[11]应用454焦磷酸测序技术分析连作和轮作条件下烟株不同生育期根际土壤细菌多样性的差异发现,烟株生育期是影响烟株根际土壤细菌结构的主要因素,轮作和连作不是影响烟株根际土壤细菌结构的主要因素,但轮作能提高土壤细菌的多样性。【研究切入点】微生物多样性扩增子测序是一种利用高通量测序技术对16S、18S、ITS等微生物特征序列进行PCR扩增并测序分析的研究方法。其中,16S rDNA(即16S rRNA gene)是原核生物核糖体RNA对应的基因片段,常用于细菌、古细菌的多样性分析。此类方法不需要对微生物进行分离纯化培养,基于提取的总DNA即可开展丰富的微生物群落研究,在医学、工业、食品、环境科学等各领域应用广泛。良好的土壤细菌结构和较高的土壤细菌多样性能改善土壤理化性质,提高土壤肥力。烟草是云南昭通主要的经济作物,尚未见分析其土壤细菌多样性的报道。【拟解决的关键问题】基于高通量测序平台,从分子水平探究昭通8个种烟县(区)的新烟区、土传病害发病严重的连作区以及典型的轮作区的土壤细菌多样性,以期了解昭通核心烟区各耕作方式下土壤细菌的多样性,为改善昭通核心烟区土壤肥力,提高耕地可持续生产能力及烟叶质量提供技术支撑。
1 材料与方法
1.1 供试土壤样品
供试土壤样品采集于昭通8个种烟县(区)的新烟区、土传病害(“两黑”病、青枯病和根结线虫病等)发病严重的连作区以及典型的轮作区(表 1),合计388个土壤样品。

表1 供试土壤样品
1.2 试验设计
从样本中提取基因组DNA后,用带有barcode(测序接头)的特异引物扩增rDNA的保守区,V3~V4的引物序列,341F,CCTACGGGNGGCWGCAG;806R,GGACTACHVGGGTATCTAAT,全长466 bp。然后PCR扩增产物切胶回收,使用Qubit 3.0进行产物定量。扩增产物进行等量混合,连接测序接头,构建测序文库,测序策略PE250在Illumina上机测序。
1.3 信息分析流程
测序得到raw Reads之后,先对低质量Reads进行过滤,然后将双端Reads拼接为Tag,再对Tag进行低质量过滤,得到的数据称为Clean Tag。基于Clean Tag,使用Usearch进行聚类,去除聚类过程中检测到的嵌合体Tag,获得OTU的丰度和OTU代表序列。
若存在有效分组,则进行组间差异比较和统计检验。最后,结合其他因素(如环境因子)进行特定的如CCA等高级分析,以解答微生物与环境之间的关系。
1.4 数据处理
将新烟区、连作区和轮作区的植烟土壤样本分别设为A组、B组和E组。根据试验需要,利用Wilcoxon秩和检验模型对不同类型土壤的细菌类群的相对丰度进行比较,明确细菌物种丰度在不同类型土壤中是否有显著差异。Sobs、Chao及ACE指数主要体现样本的物种丰富程度,值越大物种丰富程度越高;香浓指数和辛普森指数主要反映物种分布的均匀度,数值越大越均匀;香浓指数值越大,多样性越高。
2 结果与分析
2.1 昭通核心烟区土壤细菌群落组成
根据OTUs的分类关系,昭通核心烟区土壤细菌群落包括44个门、138个纲、332个目、486个科和1 044个属。
2.1.1 门水平 由图1可知,水平相对丰度排前10位的细菌群落门分别为变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、浮霉菌门(Planctomycetes)、芽单胞菌门(Gemmatimonadetes)、疣微菌门(Verrucomicrobia)、拟杆菌门(Bacteroidetes)、髌骨细菌门(Patescibacteria)和装甲菌门(Armatimonadetes)。其中,绿弯菌门和髌骨细菌门在新烟区土壤细菌群落中相对丰度较高,放线菌门在连作区土壤细菌群落中相对丰度较高,变形菌门、酸杆菌门、浮霉菌门、芽单胞菌门、疣微菌门、拟杆菌门和装甲菌门在轮作区土壤细菌群落中相对丰度较高。变形菌门、放线菌门、酸杆菌门和绿弯菌门为昭通核心烟区土壤细菌群落的优势门,4个门的序列占全部序列的68.19%~73.15%。新烟区、连作区和轮作区的土壤细菌群落最优势类群均为变形菌门,新烟区和连作区次优势类群均为放线菌门,轮作区次优势类群为酸杆菌门。
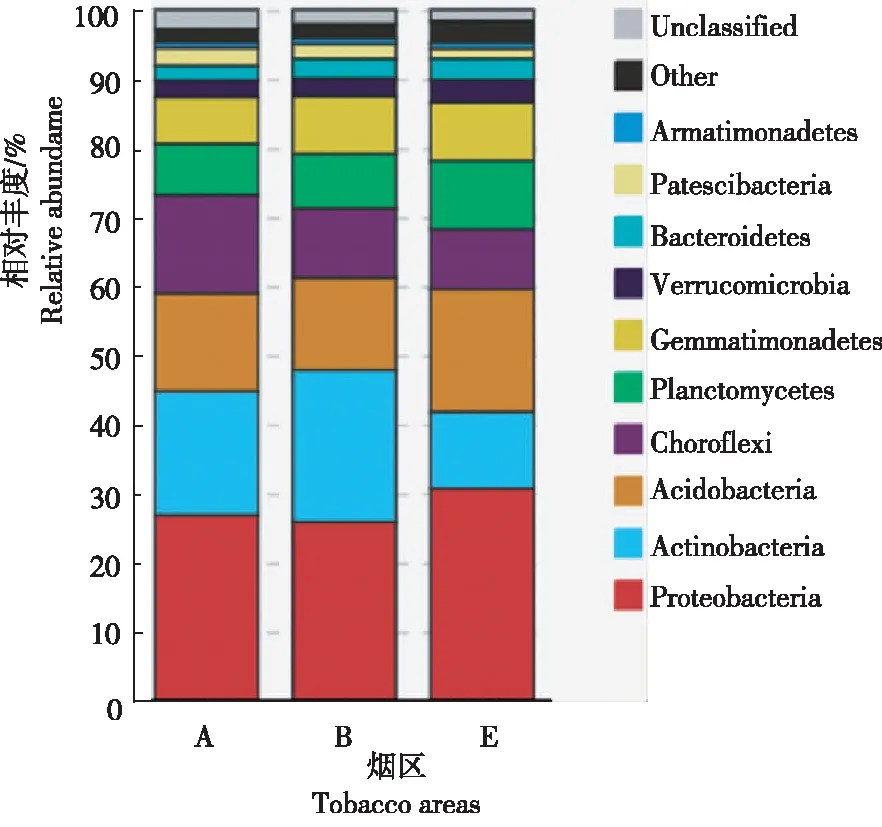
图1 3个烟区门水平上细菌类群的组成及相对丰度
2.1.2 属水平 由图2可知,属水平相对丰度前10位的分别为鞘氨醇单胞菌属(Sphingomonas)、芽单胞菌属(Gemmatimonas)、Chujaibacter、链霉菌属(Streptomyces)、MND1、Bryobacter、RB41、Candidatus_Solibacter、Candidatus_Udaeobacter和Ellin6067。在新烟区土壤中,芽单胞菌属、Chujaibacter、Bryobacter和Candidatus_Solibacter相对丰度较高;在连作区土壤中,链霉菌属和Ellin6067相对丰度较高;在轮作区土壤中,鞘氨醇单胞菌属、ND1、RB41和Candidatus_Udaeobacter相对丰度较高。因此,昭通核心烟区土壤的最优势细菌类群为鞘氨醇单胞菌属,占所有序列的4.44%~6.58%。在新烟区土壤中,细菌群落相对丰度前3位的属分别为鞘氨醇单胞菌属、芽单胞菌属和Chujaibacter;在连作区土壤中,相对丰度前3位的属分别为鞘氨醇单胞菌属、芽单胞菌属和链霉菌属;在轮作区土壤中,相对丰度前3位的属分别为鞘氨醇单胞菌属、RB41和芽单胞菌属。
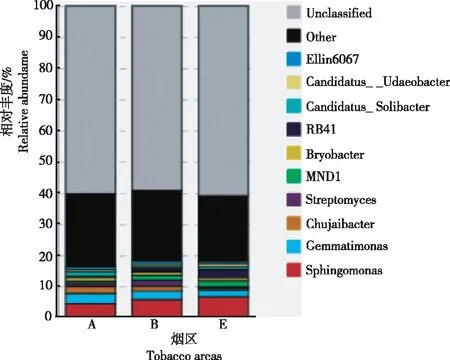
图2 3个烟区属水平上细菌类群的组成及相对丰度
2.2 昭通核心烟区土壤细菌群落指示物种
2.2.1 优势物种 由图3可知,在门水平,放线菌门、绿湾菌门、芽单胞菌门、疣微菌门、拟杆菌门和髌骨细菌门等6个门的相对丰度在新烟区与连作区土壤细菌群落间均存在显著差异,变形菌门、放线菌门、酸杆菌门、绿弯菌门、浮霉菌门、芽单胞菌门、疣微菌门、拟杆菌门和髌骨细菌门等9个门的相对丰度在新烟区与轮作区土壤细菌群落间均存在显著差异,变形菌门、放线菌门、酸杆菌门、浮霉菌门、疣微菌门和髌骨细菌门等6个门的相对丰度在连作区与轮作区土壤细菌群落间均存在显著差异。新烟区土壤中绿湾菌门的相对丰度显著高于连作区和轮作区,芽单胞菌门和拟杆菌门的相对丰度显著低于连作区和轮作区;连作区土壤中放线菌门的相对丰度显著高于新烟区和轮作区;轮作区土壤中变形菌门、酸杆菌门、浮霉菌门和疣微菌门的相对丰度显著高于新烟区和连作区。
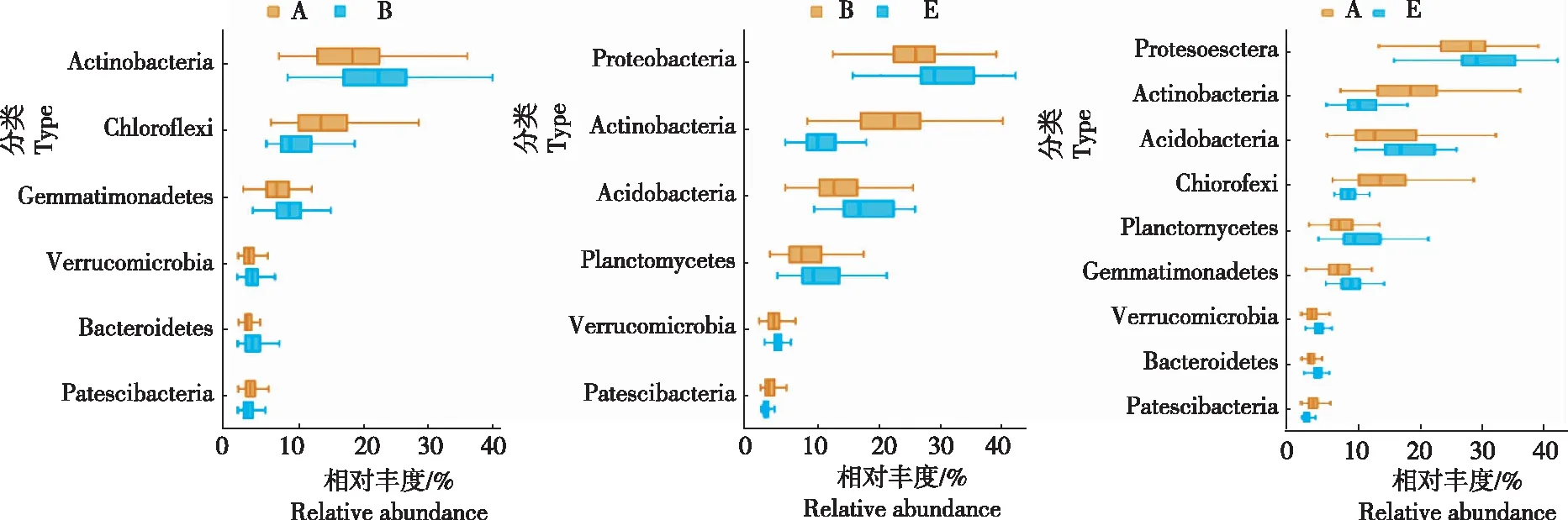
图3 在门水平上细菌群落相对丰度的Wilcoxon秩和检验
由图4可知,在属水平,鞘氨醇单胞菌属、芽单胞菌属、链霉菌属、MND1、Bryobacter、RB41、Candidatus_Solibacter和Ellin6067等8个属的相对丰度在新烟区与连作区土壤细菌群落之间均存在显著差异,鞘氨醇单胞菌属、芽单胞菌属、Chujaibacter、链霉菌属、MND1、Bryobacter、RB41、Candidatus_Solibacter和Candidatus_Udaeobacter等9个属的相对丰度在新烟区与轮作区土壤细菌群落之间均存在显著差异,鞘氨醇单胞菌属、芽单胞菌属、Chujaibacter、链霉菌属、MND1、RB41、Candidatus_Solibacter、Candidatus_Udaeobacter和Ellin6067等9个属的相对丰度在连作区与轮作区土壤细菌群落之间均存在显著差异。新烟区土壤中鞘氨醇单胞菌属、MND1和RB41的相对丰度显著低于连作区和轮作区,芽单胞菌属、Bryobacter和Candidatus_Solibacter的相对丰度显著高于连作区和轮作区;连作区土壤中Ellin6067的相对丰度显著高于新烟区和轮作区;轮作区土壤中Chujaibacter和链霉菌属的相对丰度显著低于新烟区和连作区,Candidatus_Udaeobacter的相对丰度显著高于新烟区和连作区。

图4 在属水平上细菌群落相对丰度的Wilcoxon秩及检验
2.2.2 特有物种 由图5和表2可知,新烟区土壤细菌群落有2个特有纲、7个特有目、11个特有科和29个特有属,连作区有3个特有门、5个特有纲、11个特有目、19个特有科和53个特有属,轮作区有1个特有门、3个特有纲、6个特有目、8个特有科和13个特有属。连作区土壤细菌群落特有物种多于新烟区和轮作区。

注:a~e分别为门、纲、目、科及属水平物种VENN图。
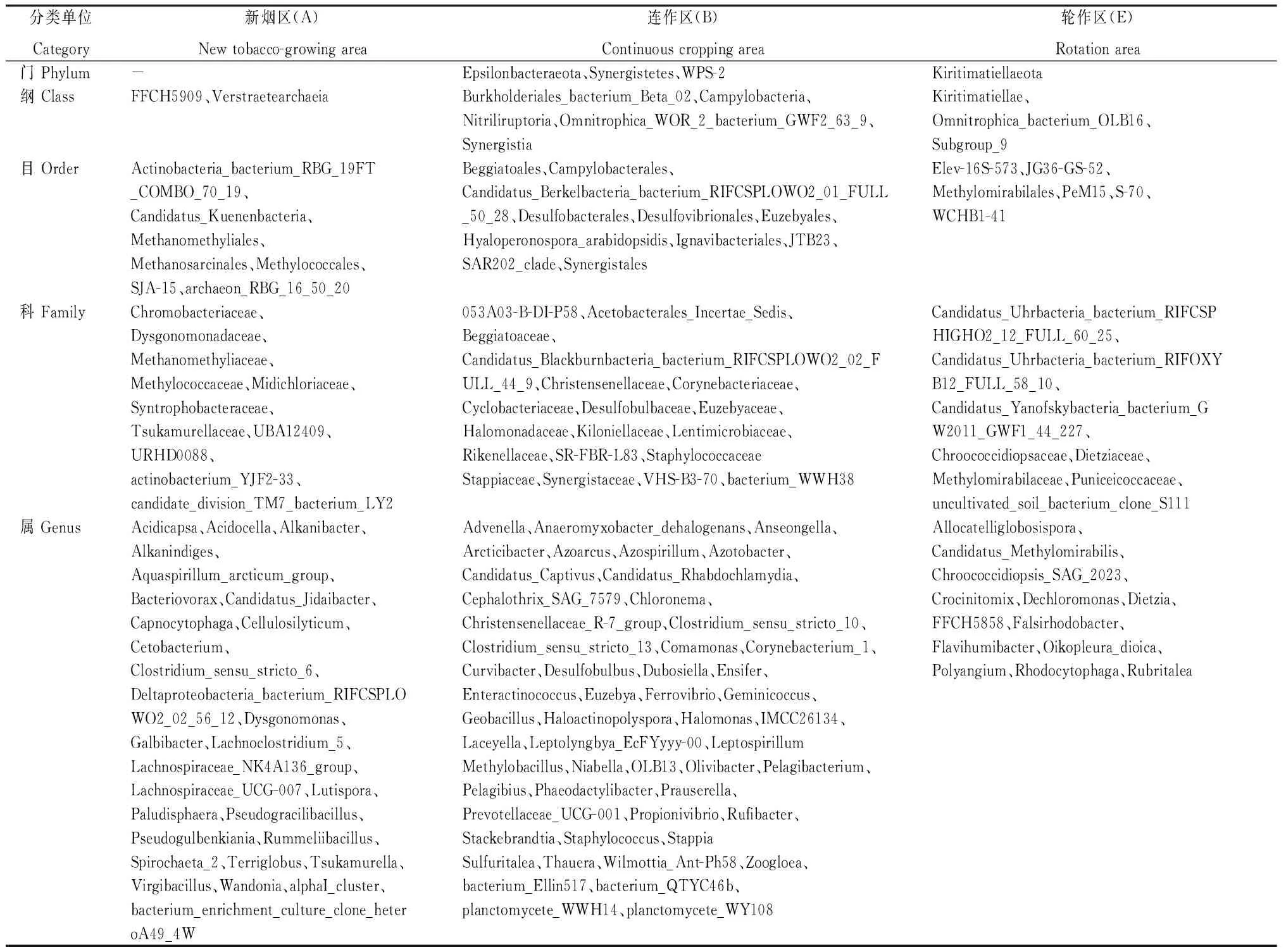
表2 昭通核心烟区不同类型土壤的细菌群落特有物种
2.2.3 指示物种 从图6看出,新烟区、连作区和轮作区的土壤细菌群落有重叠部分,但有部分样本点相互的距离较远,且X轴(32.09%)与Y轴(14.10%)累计贡献量达46.19%,说明不同类型土壤的细菌群落结构组成有一定差异。

图6 昭通核心烟区3种类型土壤细菌群落组间比较
根据分类学组成对新烟区、连作区和轮作区土壤进行线性判别分析(LDA),找出对不同类型土壤产生显著性差异影响的群落,仅LDA得分值大于4的被展示(图7)。新烟区土壤细菌群落中差异指示物种为嗜酸杆菌纲、酸杆菌目、Frankiales目、纤线杆菌(从纲到科)和罗丹诺杆菌科,连作区差异指示物种为放线菌纲和嗜热油菌纲,轮作区差异指示物种为Pyrinomonadaceae(从目到科)、鞘氨醇单胞菌(从目到属)、δ-变形菌纲、β-变形菌目。以上物种对昭通核心烟区新烟区、连作区和轮作区土壤间细菌群落结构组成的差异起重要作用。
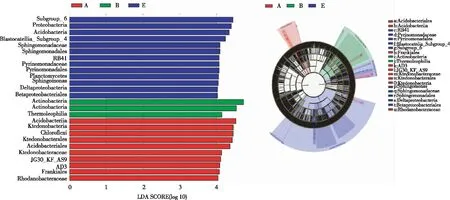
图7 昭通核心烟区3种不同类型土壤细菌群落的特殊群落及影响力
2.3 昭通核心烟区土壤细菌群落生物多样性
从表3可见,昭通核心烟区土壤细菌群落的Sobs指数、Chao指数、ACE指数和香浓指数均以轮作区最高,连作区次之,新烟区最低;辛普森指数以轮作区最高,新烟区次之,连作区最低。由图8可见,轮作区的Sobs指数、Chao指数、ACE指数、香浓指数和辛普森指数与连作区均有极显著差异,Sobs指数、Chao指数、ACE指数和香浓指数与新烟区均有极显著差异,辛普森指数与新烟区有显著差异;连作区的Sobs指数与新烟区有显著差异,Chao指数、ACE指数、香浓指数和辛普森指数与新烟区差异不显著。生物多样性是指特定生境或生态系统内的多样性情况,通常利用物种丰富度(种类情况)与物种均匀度(分布情况)2个重要参数判定。可见,轮作区土壤细菌群落的生物多样性最高;连作区土壤细菌群落的物种丰富度高于新烟区,物种均匀度与新烟区相当;连作区细菌群落生物多样性略高于新烟区。
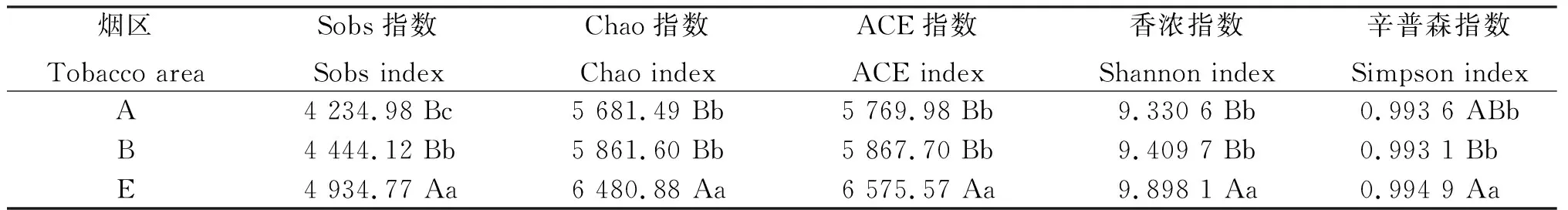
表3 昭通核心烟区3种不同类型土壤细菌群落的生物多样性指数

图8 昭通核心烟区3种不同类型土壤细菌群落的生物多样性指数
由图9可知,3种不同类型土壤细菌的稀释曲线均趋于平缓,说明基于研究的测序深度,土壤中包括稀有物种在内的所有细菌均已分析,真实地反映了该研究区的细菌群落组成。

图9 昭通核心烟区3种不同类型土壤细菌的稀释曲线
2.4 昭通核心烟区土壤细菌群落的PICRUSt功能
从表4可见,昭通核心烟区土壤细菌群落Pathway level 1有6条代谢通路,Pathway level 2有35条代谢通路。在生态环境类似的条件下,新烟区、连作区和轮作区土壤细菌群落代谢通路相似,但代谢通路的丰度值不同。如图10 所示,除外源生物降解与代谢、细胞运动性和信号传导相关代谢通路的丰度值新烟区和连作区差别较小外,其余代谢通路的丰度值以连作区最高,新烟区次之,轮作区最低。如图11所示,丰度值排列的代谢通路中碳水化合物代谢、氨基酸代谢、辅助因子和维生素的代谢、萜类和聚酮类代谢、其他氨基酸的代谢、脂类代谢、外源生物降解与代谢、能量代谢、复制和修复以及折叠、定位和降解等通路的丰度值在新烟区与连作区间、新烟区与轮作区间、连作区与轮作区间均存在显著差异。整体看,土壤细菌群落代谢通路的丰度值连作区最高,新烟区次之,轮作区最低,表明土壤细菌群落代谢功能潜力连作区最高,新烟区次之,轮作区最低。
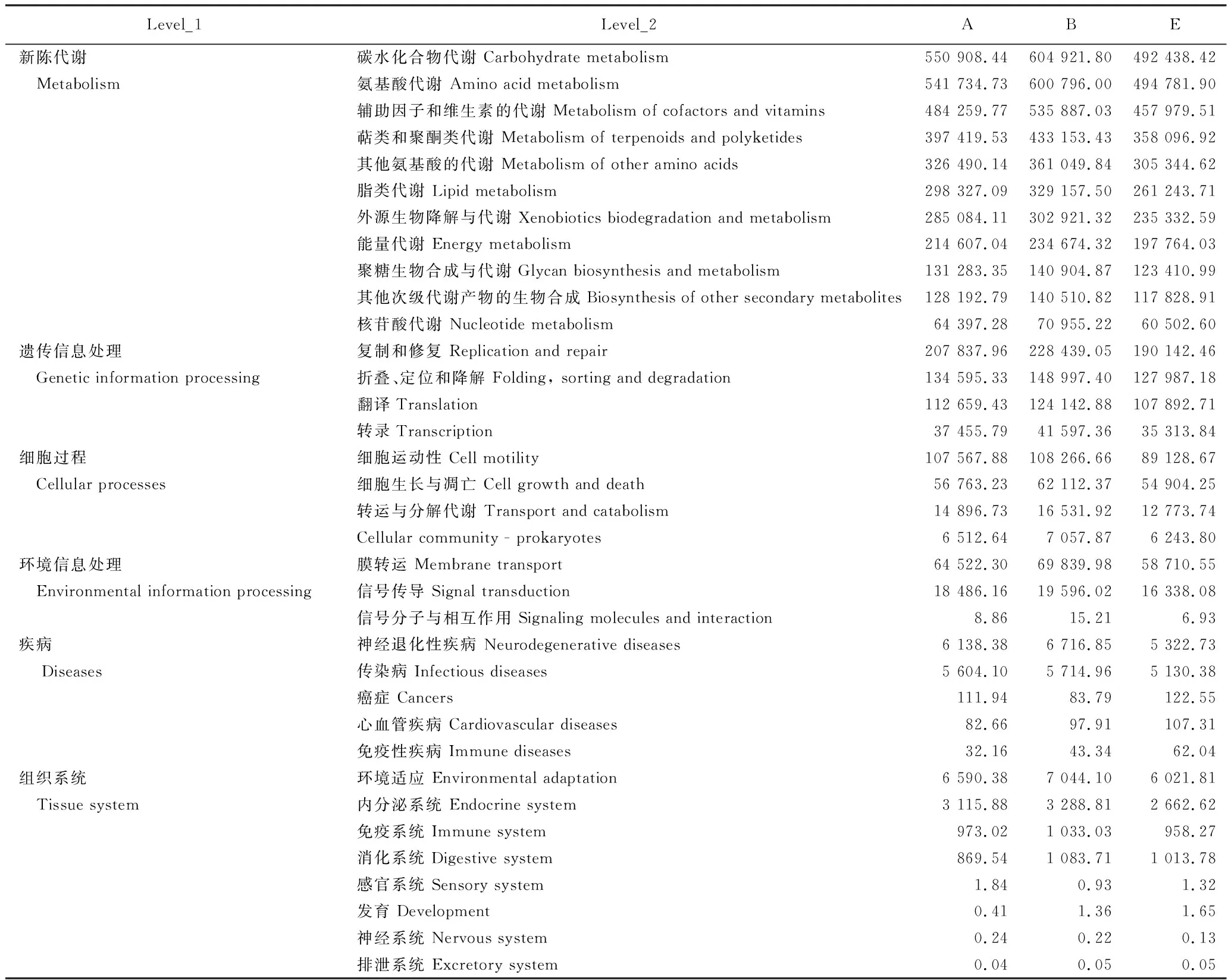
表4 昭通核心烟区3种不同类型土壤的细菌群落功能分布

图 10 昭通核心烟区3种不同类型土壤的细菌群落功能丰度热图
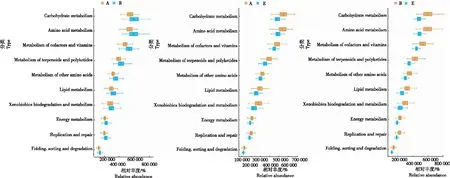
图11 细菌群落代谢通路丰度值的Wilcoxon秩及检验
3 讨论
高通量测序技术可对几百万个DNA分子进行有效的序列测定,能够客观真实地反映环境微生物群落结构特征[12],被广泛应用于环境样品微生物群落组成的分析研究[13-14],其研究手段不依赖于传统的微生物分离培养,而是直接从环境中获取基因信息,其测序结果可以包含大多数弱势种群,可以更全面地反映微生物多样性的真实水平[15-16]。微生物多样性研究主要分为alpha、beta多样性研究、物种分析、功能研究、环境关系研究。本研究应用高通量测序技术对昭通核心烟区土壤细菌群落结构进行分析表明,昭通核心烟区土壤细菌菌群极其丰富,包括44个门、138个纲、332个目、486个科和1 044个属。细菌群落门水平相对丰度前10位的分别为变形菌门、放线菌门、酸杆菌门、绿弯菌门、浮霉菌门、芽单胞菌门、疣微菌门、拟杆菌门、髌骨细菌门和装甲菌门,其中,变形菌门、放线菌门、酸杆菌门和绿弯菌门为昭通核心烟区土壤细菌群落的优势门,这4个门的序列占所有序列的68.19%~73.15%,其相对丰度因土壤类型而异,与前人研究结果[17-20]一致。属水平相对丰度前10位的分别为鞘氨醇单胞菌属、芽单胞菌属、Chujaibacter、链霉菌属、MND1、Bryobacter、RB41、Candidatus_Solibacter、Candidatus_Udaeobacter和Ellin6067,其中,鞘氨醇单胞菌属为昭通核心烟区土壤细菌群落的最优势属,占所有序列的4.44%~6.58%,其相对丰度因土壤类型而异。同时,在3类土壤样品中,Unclassified bacteria占有一定比例,说明在昭通核心烟区土壤中仍存在大量未知和稀有的细菌[21]。
不同类型土壤的细菌群落结构有一定差异[22]。本研究中,3类土壤细菌群落最优势门均为变形菌门,新烟区和连作区次优势门均为放线菌门,轮作区次优势门为酸杆菌门;新烟区土壤细菌群落芽单胞菌属的相对丰度显著高于连作区和轮作区,连作区链霉菌属的相对丰度显著高于新烟区和轮作区,轮作区鞘氨醇单胞菌属的相对丰度显著高于新烟区和连作区。因为连作导致土壤理化性质恶化、养分失调以及微生物区系发生变化[23],可使土壤中鞘氨醇单胞菌属、芽孢杆菌属等有益菌数量减少,从而破坏微生物群落平衡,导致土壤连作障碍的发生[24]。
轮作能提高植烟土壤细菌生物多样性[8,18]。本研究基于种的分类水平对新烟区、连作区和轮作区土壤细菌群落多样性指数进行分析表明,轮作区土壤细菌群落的生物多样性最高;连作区土壤细菌群落的物种丰富度略高于新烟区,物种均匀度与新烟区相当,其细菌群落生物多样性略高于新烟区。前人研究表明,连作使植烟土壤细菌种类减少,群落结构趋于简单,可能是由于连作种植单一或存在连作障碍因子等,限制了土壤某些细菌的生长[25],同时刺激了其他种类细菌的生长而促使其成为优势种类,使得土壤细菌种类较少[26]。土壤微生物群落数量及结构的变化直接影响着根际微生物整体功能,一般来说,多样性指数较高,系统的结构也较复杂,稳定性相对较高[27]。经对昭通核心烟区土壤细菌群落进行功能预测表明,在生态环境类似的条件下,新烟区、连作区和轮作区土壤细菌群落代谢通路相似,但代谢通路的丰度值不同,以连作区土壤细菌群落代谢通路的丰度值最高,新烟区次之,轮作区最低。
4 结论
昭通核心烟区土壤细菌群落组成具有多样性,轮作区土壤细菌群落的生物多样性最高,连作区土壤细菌群落生物多样性略高于新烟区。轮作区、连作区和新烟区的土壤细菌优势门均为变形菌门,优势属均为鞘氨醇单胞菌属。不同类型土壤的细菌群落结构组成有一定差异,连作区土壤细菌群落特有物种多于新烟区和轮作区。在生态环境类似的条件下,新烟区、连作区和轮作区土壤细菌群落代谢通路相似,但是代谢通路的丰度值不同,连作区植烟土壤细菌群落代谢功能潜力最高。
